 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
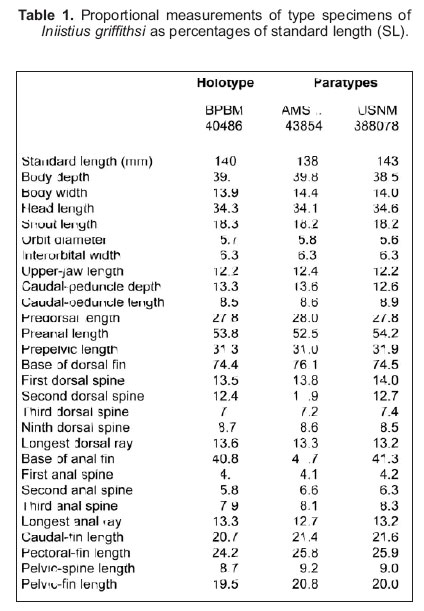
Smithiana, Publications in Aquatic Biodiversity, Bulletin 7, May, 2007, pp. 9-13 Iniistius griffithsi, a new razorfish (Perciformes: Labridae) from Mauritius John E. Randall Bishop Museum, 1525 Bernice St., Honolulu, HI 96817-2704, USA. e-mail: jackr@hawaii.rr.com (Received 24 October 2006; accepted 2 January 2007) Code Number: sm07002 ABSTRACT The razorfish Iniistius griffithsi is described from three male specimens caught by handline off the south coast of Mauritius in 120 m. It is distinct in lacking scales on the cheek except for a curved row of 6 small scales from behind to below the eye; 19–20 gill rakers; body depth of 2.5–2.6 in standard length; first two dorsal spines slender, flexible, and not long (the first 2.45–2.55 in head length); space between first and second dorsal spines more than twice that between second and third spines; and in colour. The most conspicuous colour markings are a broad oblique pale blue band containing many small black spots from high on the nape to above the opercle, and black spots of about pupil size, each edged in pale blue, on the side of the body above the tip of the pectoral fin. KEY WORDS: Taxonomy, Labridae, Iniistius griffithsi sp. nov., Mauritius INTRODUCTION Labrid fishes of the six genera of the subfamily Xyrichtyinae (Hemipteronotinae of Norman 1957) contain species that are able to escape predation by quickly diving into sand: Ammolabrus, Cymolutes, Iniistius, Novaculichthys, Novaculoides and Xyrichtys. Three of the genera are monotypic: Ammolabrus dicrus Randall & Carlson is a zooplankton feeder that forms aggregations over open sand substrata; Novaculichthys taeniourus (Lacepéde) is usually seen over sand and rubble areas near reefs; and Novaculoides macrolepidotus (Bloch) typically hides in seagrass or algal beds, which it resembles in its colouration. The species of the other three genera may be found over broad open stretches of sand far from the shelter of reefs or dense plant growth needed for shelter by other benthic shore fishes. The three species of Cymolutes have been given the common name knifefishes, and those of Xyrichtys and Iniistius are called razorfishes, in both cases alluding to their compressed bodies and the sharp leading edge of their forehead and snout, specialisations for quick entry into sand. Some razorfishes have been classified in the genus Hemipteronotus Lacepéde. As recommended by Randall & Bauchot (1993), this generic name was suppressed by Opinion 1799 of the International Commission on Zoological Nomenclature to preserve the names Naucrates and Xyrichtys. Iniistius has been considered as a synonym of Xyrichtys by some authors. Randall & Earle (2002) followed Tri-thuc Nguyen (1974) who differentiated the two genera osteologically. Externally, the two genera can be distinguished by three characters of the dorsal fin: the origin of the fin is over or less than half an orbit diameter behind the eye in Iniistius, but an eye diameter or more behind the eye in Xyrichtys. The first two dorsal spines of Iniistius are flexible, whereas only the first is flexible in Xyrichtys. The space between the second and third dorsal spines in Iniistius is much broader than the space between the third and fourth spines; by contrast these two spaces are about equal in Xyrichtys. Randall & Earle provided a diagnosis of Iniistius. Species of Xyrichtys occur in the Atlantic, eastern Pacific and the Indo-Pacific region, whereas those of Iniistius are confined to the Indo-Pacific, except I. pavo (Valenciennes), which has extended its range to the eastern Pacific. As might be expected from their habits, razorfishes are not well represented in museum collections. One is described here in the genus Iniistius from three specimens from Mauritius. MATERIALS AND METHODS Type specimens are deposited in the Australian Museum, Sydney (AMS); Bernice P. Bishop Museum, Honolulu (BPBM); and U.S. National Museum of Natural History, Washington, D.C. (USNM). The length of specimens is given as standard length (SL), measured from the median anterior end of the upper lip to the base of the caudal fin (posterior end of the hypural plate); body depth is the greatest depth fromthe base of the dorsal spines to ventral edge of the abdomen (correcting for any malformation of preservation); body width is measured just posterior to the gill opening; head length is taken fromthe upper lip to the posterior end of the opercular flap; orbit diameter is the greatest fleshy diameter, and interorbital width the least bony width; snout length is measured from the median anterior point of the upper lip to the nearest fleshy edge of the orbit; upper-jaw length from the same anterior point to the posterior end of the maxilla; caudal-peduncle depth is the leastdepth, and caudal-peduncle length the horizontal distance between verticals at the rear base of the anal fin and the caudal-fin base; lengths of spines and rays are measured to their extreme bases; caudal-fin and pectoral-fin lengths are the length of the longest ray; pelvic-fin length is measured from the base of the pelvic spine to the tip of the longest soft ray. Morphometric data are presented in Table 1 as percentages of the standard length. Proportional measurements in the text are rounded to the nearest 0.05. Lateral-line scale counts include the last pored scale that overlaps the end of the hypural plate; scales in transverse series are counted from the origin of the anal fin obliquely upward to the base of the first dorsal fin; the count of gill rakers is made on the first gill arch and includes all rudiments. Meristic and morphometric data in parentheses refer to paratypes. Iniistius griffithsi, sp. nov. Holotype: BPBM 40486, male, 140 mm SL, Mauritius, south coast, 0.5 km south of mouth of Riviére des Anguilles, about 1 km offshore from point at 20°31.2’S, 57°33’E, 120 m, handline, Jeremy Griffiths and Tonio Isidore, 1 May 2006. Paratypes: AMS I.43854-001, male, 138 mm and USNM 388078, male, 143 mm, same data as holotype, except March, 2006. DIAGNOSIS. Dorsal rays IX,12; anal rays III,12; pectoral rays 12; lateral line interrupted, the pored scales 19–20 + 5 or 6; cheek naked except for a curved oblique row of 6 small scales from behind to below eye; 1 or 2 small scales dorsoanteriorly on opercle; gill rakers 19-20; body depth 2.5–2.6 in SL; dorsal profile of snout nearly vertical; first two dorsal spines slender and flexible, the first slightly longer, 2.45–2.55 in head length; space between second and third dorsal spines more than twice that between first and second spines; colour when fresh, pale grey, scale edges dorsally on body grey-brown; head grey-brown, operculum suffuse d with orange, with vertical blue bands; a broad oblique pale blue band containing numerous small black spots from interorbital space to above opercle; one to several black spots, edged in pale blue, on side of body above tip of pectoral fin; a dusky streak below posterior end of maxilla and lower lip; orangish patch on side of caudal peduncle, each scale with a pale blue spot; median fins bluish grey; pectoral fins orange. DESCRIPTION. Dorsal rays IX,12, all soft rays branched (first 4 or 5 rays of paratypes unbranched), the last to base; anal rays III,12, all branched (first ray of one paratype unbranched), the last to base; pectoral rays 12, the uppermost very short and spine-like, the second unbranched; pelvic rays I,5, all soft rays branched; principal caudal rays 12, the uppermost unbranched; upper procurrent caudal rays 6; lower procurrent caudal rays 5; lateral-line scales 20 + 5 (19–20 + 5–6), the last on base of caudal fin; scales above lateral line to origin of dorsal fin 4; scales above lateral line to middle of spinous portion of dorsal fin 2 (uppermost small); scales below lateral line to origin of anal fin 9; circumpeduncular scales 14; gill rakers 20 (19–20); branchiostegal rays 5; vertebrae 25. Body deep, the depth 2.55 (2.5–2.6) in SL; body very compressed, the width 2.8 (2.75) in body depth; head length 2.9 (2.9–2.95) in SL; snout length (as measured from lower edge of orbit to front of upper lip) 1.9 (1.85–1.9) in head length; dorsal profile of snout nearly vertical to level of lower edge of eye, then convex to above eye (indentation of profile above upper lip of holotype the result of a wound, probably from the hook); front of snout narrowing to sharp ridge that extends to above eye; front of chin also a ridge though not as sharp as that of snout; eye set high on head, but nearly an orbit diameter vertically below dorsal profile of interorbital space; orbit diameter 6.0 (5.9-6.2) in head length; interorbital width 5.45 (5.4-5.5) in head length; caudal-peduncle depth 2.6 (2.5-2.75) in head length; caudal-peduncle length 4.05 (3.9-4.0) in head length. Mouth moderately large, the maxilla extending to or nearly to a vertical at anterior edge of orbit, the upper-jaw length 2.8 (2.75–2.85) in head length; mouth slightly oblique, forming an angle of about 20° to horizontal axis of body; a pair of large, recurved, outflaring canine teeth at front of jaws that overlap lips when mouth closed, the lower pair medial to upper; side of jaws with a close-set series of strong conical teeth, 14 on upper jaw and 13 in lower jaw of holotype; two to three irregular rows of small, slightly nodular teeth medial to row of conical teeth in upper jaw, and one to two rows in lower jaw. Tongue broadly rounded, set far back in mouth. Lips thin, the lower with a well-developed flap along side ofmandible. Gill rakers short, the longest on first arch about one-half length of longest gill filaments. Posterior edge of preopercle free nearly an eye diameter below ventral edge of orbit, the ventral edge nearly or just reaching a vertical at posterior edge of maxilla; a narrow fleshy flap surrounding orbit from below anterior edge of pupil to above posterior edge of pupil. Nostrils very small, before lower fourth of eye, the anterior two-thirds orbit diameter before eye, porelike with a short rim and dorsoposterior flap; posterior nostril a short, near-horizontal slit dorsoposterior to anterior nostril, the internarial space nearly one-half pupil diameter. Cephalic sensory pores tiny; suborbital pores 6, with 2 isolated pores one above the other, nearly an orbit diameter below front edge of eye; a series of 8 preopercular pores, continuing anteriorly as 3 mandibular pores. Scales cycloid and very thin; lateral-line scales with a single horizontal tubule, ending posteriorly in a pore; scales on chest about half height of largest scales on side of body; head naked except for one or two small scales dorsally on opercle and a curved series of six scales, progressively smaller ventrally from behind lower part of eye to below posterior edge of pupil (paratypes with two or three small partially embedded scales ventral to lowermost scale in oblique row); no scales on dorsal and anal fins; two or three rows of scales basally on caudal fin; no pelvic axillary scale; a single scale extending posteriorly from between base of pelvic fins, its length about equal to half length of pelvic spine. Origin of dorsal fin above posterior edge of orbit, the predorsal length 3.6 in SL; first two dorsal spines slender and flexible, the remaining spines sharppointed and stiff; space between second and third dorsal spines more than twice space between first two spines; membrane deeply notched between second and third spines; first dorsal spine longest, slightly longer than second, 2.55 (2.45) in head length; third dorsal spine 4.85 (4.7–4.75) in head length: remaining dorsal spines progressively longer, the ninth 3.95 (3.95–4.05) in head length; fifth and sixth dorsal rays longest, 2.5 (2.55–2.6) in head length; origin of anal fin below base of first dorsal soft ray, the preanal length 1.85 (1.85–1.9) in SL; third anal spine longest, 4.35 (4.15–4.2) in head length; fifth anal soft ray longest, 2.6 (2.6–2.7) in head length; caudal fin slightly rounded, 4.85 (4.65) in SL; third and fourth pectoral rays longest, reaching to above origin of anal fin, 4.15 (3.85–3.9) in SL (distal end of about upper fourth of pectoral fin of right side of holotype missing); pelvic fins nearly or just reaching anus, the first soft ray longest, 5.15 (4.8–5.0) in SL; pelvic spine 2.25 (2.2–2.25) in length of longest pelvic soft ray. Colour of holotype in alcohol: head, chest and most of abdomen purplish grey; body above lateral line dull greyish orange; broad middle zone of body yellowish white with a narrow midlateral dull orange stripe along middle of scale row; three black spots the size of pupil forming a triangle on side of body above distal end of pectoral fin, the uppermost spot on seventh lateral-line scale; scales on posterior fourth of body and base of caudal fin purplish grey, the scale edges paler; a broad oblique whitish band containing numerous small black spots of variable size from above eye, passing above operculum to level of lower edge of eye; lips and chin pale with a grey streak below ventral flap of lower lip to end ofmaxilla; dorsal and caudal fins translucent grey; anal and paired fins pale yellowish. Colour of holotype when fresh as shown in Fig. 1. One of the paratypes has only a single pupil-size black spot below the seventh lateral-line scale and no others below. The other has black spots on nine scales, including sixth and seventh lateral-line scales of left side and sixth scale of right side, with one spotted scale above and six or seven on the two scale rows below. In addition, the second paratype has an isolated scale on one side below outer part of pectoral fin with two black spots. All of these spots were edged in pale blue in the newly caught fish. It is regretted that no female specimens were obtained. In view of the sexual dichromatism known for the species of Iniistius, the female can be expected to exhibit some difference in colour pattern. Juveniles will probably have still another pattern, most likely one with dark bars. ETYMOLOGY. This species is named for the collector, Jeremy Griffiths. REMARKS. The three specimens of Iniistius griffithsi were caught by handline from 120 m, hence deeper than normal scuba-diving depths. Fishes of this genus are highly esteemed as food fishes in spite of their relatively small size. They are often consumed by the fishermen themselves, therefore not often found in markets. Two specimens of Iniistius pavo (Valenciennes) were caught from the same location and depth as the three of I. griffithsi. Iniistius pavo and the western Pacific I. dea (Temminck & Schlegel) are easily distinguished from all other species of the genus by the much longer and more forward position of the first two dorsal spines and their complete separation from the remaining spinous portion of the fin. As noted by Randall et al. (2002), the remaining species of the genus Iniistius are divisible into two lineages, one with a broad band of scales that extend ventrally from below the eye to at least the level of the corner of the mouth, and other with just a few scales below the eye. Iniistius griffithsi falls in the latter group for which only I. aneitensis (Günther) and I. cyanifrons (Valenciennes) are known. It is easily distinguished from these two species by its striking black-spotted pattern. ACKNOWLEDGEMENTS I am most grateful to Owen Griffiths in Mauritius for noting that this species is an unusual fish, arranging for the collection of an additional specimen, and for its photograph by Dominique Lebreton. When informed of the plan to describe the fish in his honour, he asked that it be named for his son Jeremy, one of the two fishermen who caught the type specimens. Griffiths’ first two specimens were given to the Australian Museum; Mark A. McGrouther kindly passed them on loan to me. Thanks are also due Loreen R. O’Hara for taking x-rays. LITERATURE CITED
ADDENDUM While this paper was in press, the author determined that Gerald R. Allen observed about 10 individuals of Iniistius griffithsi varying from about 8–18 cm in total length in the outer section of Loky Bay, northeastern Madagascar (12°43.532’S, 49°42.386’E) on a clean sand bottom at a depth of 8 m in March 2006. His underwater photographs of two adult individuals are reproduced here as Figs. 2 and 3. Allen observed a similar fish at Christmas Island in the Indian Ocean in July, 2006. © 2007 The South African Institute for Aquatic Biodiversity, Grahamstown, South Africa The following images related to this document are available:Photo images[sm07002t1.jpg] [sm07002f3.jpg] [sm07002f1.jpg] [sm07002f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}