 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
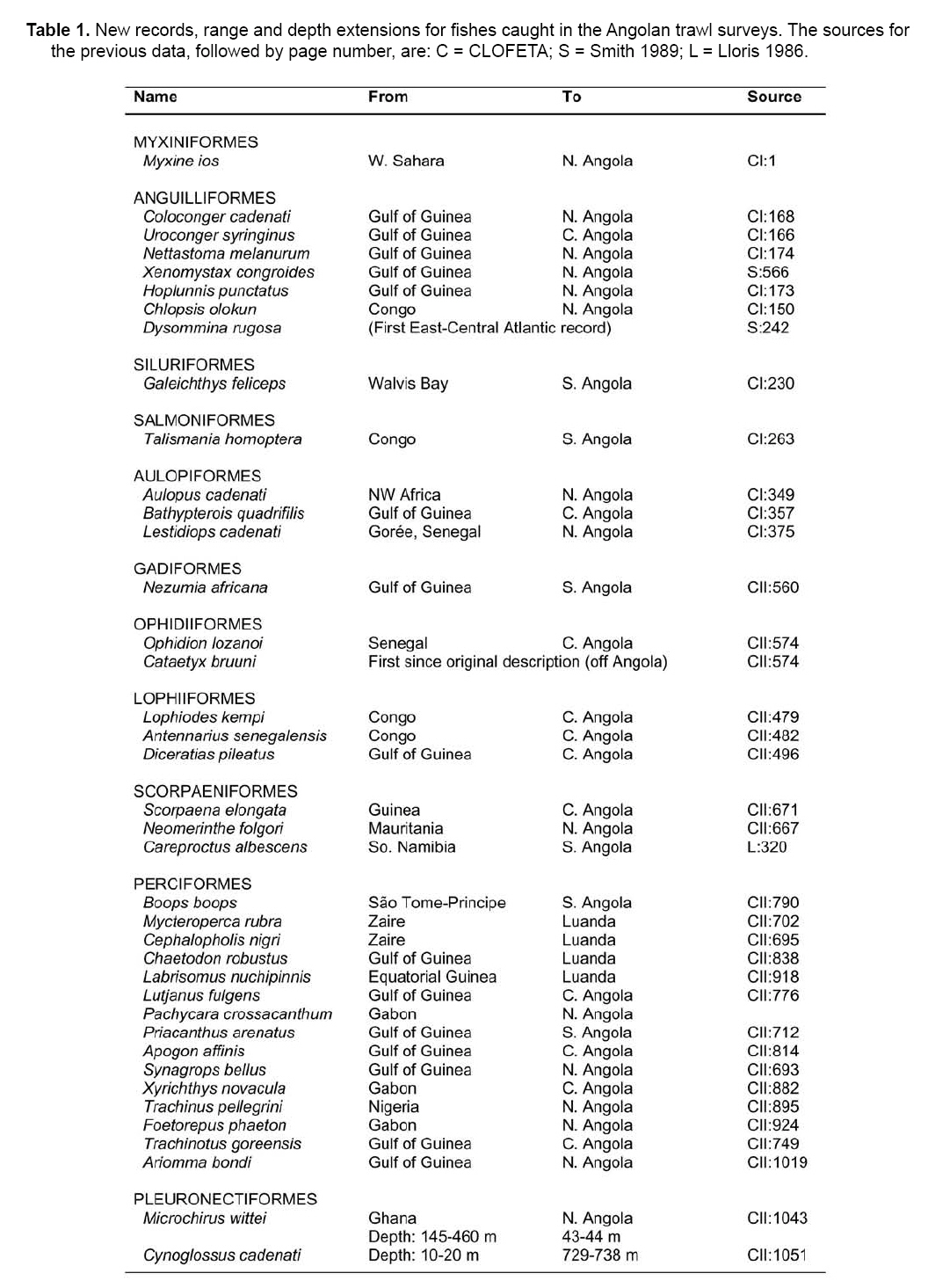
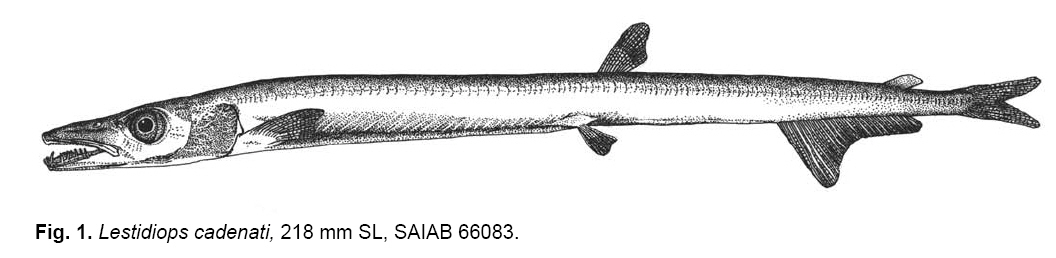
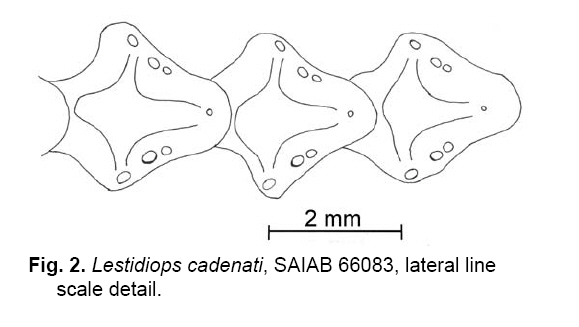
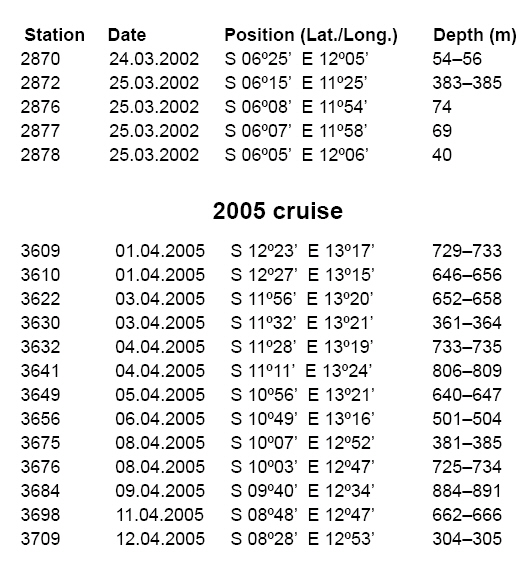
Smithiana, Publications in Aquatic Biodiversity, Bulletin 8, February, 2008, pp. 3-24 A collection of marine fishes from Angola, with notes on new distribution records Denis Tweddle and M. Eric Anderson South African Institute for Aquatic Biodiversity, Private Bag 1015, Grahamstown 6140, RSA Received 16 March 2007; accepted 1 October 2007 Code Number: sm08001 ABSTRACT Collections of fishes from demersal trawl surveys to 800 m depth off the Angolan coast in 2001, 2002 and 2005 resulted in several range extensions tabulated here. Specimens of species poorly represented in previous collections allowed improved diagnoses of Myxine ios, Torpedo bauchotae, Dysommina rugosa, Pisodonophis semicinctus, Xenomystax congroides, Lestidiops cadenati, Ophidion lozanoi, Cataetyx bruuni, Dibranchus atlanticus, Diceratias pileatus, Himantolophus paucifilosus, Neomerinthe folgori, Careproctus albescens, Paracaristius maderensis and Pachycara crossacanthum. All specimens accessioned into the Fish Collection of the South African Institute for Aquatic Biodiversity are listed and colour plates show a selection of species from the trawl catches. KEY WORDS: demersal trawl, Gulf of Guinea, Benguela Current, range extensions, species diagnoses INTRODUCTION The Angolan coast is an area of high biodiversity interest as it forms the boundary between the fauna of the temperate Benguela current, arising from upwelling off the Namibian coast to the south, and the tropical waters of the Gulf of Guinea to the north. The sea off Angola therefore supports a rich and diverse fauna. Few survey studies of fish off the Angolan coast have been made. Poll (1951, 1953, 1954, 1959) reported on work carried out by L'Expédition Océanographique Belge dans l'Atlantique Sud of 1948-49. Lloris (1984, 1986) reported on mainly deepwater fishes from Spanish government surveys run through the Instituto de Ciencias del Mar, Barcelona, along the adjacent Namibian coast. Surveys in the area by the former Soviet Union were conducted during the 1970s and 1980s (Trunov 1981), but many of these collections remain unreported. A few other summaries and guides have been published that are relevant to the Angolan marine fish fauna. The FAO Species Identification Sheets for the eastern central Atlantic (Fischer et al. 1981) provide identification material for mostly commercial species. The Check-list of the fishes of the eastern tropical Atlantic (or 'Clofeta'), in three volumes, is a comprehensive guide to the relevant literature on fishes of the area (Quero et al. 1991). Smiths' Sea Fishes (Heemstra & Smith 1986) covers the southwestern African coast up to Namibia and thus is a partial guide to the Benguela system fishes, while the dated Blache et al. (1970) covers the eastern Atlantic coast to the north of Angola. Extensive demersal trawl surveys off Angola in March 2001 and March 2002 in depths of 20 to 800 m by the RV Dr Fridtjof Nansen under a UNDP/NORAD programme provided South African Institute for Aquatic Biodiversity (SAIAB) with an opportunity to make large representative collections of the fishes of the region. The collection had two purposes, (1) to enhance the Institute's existing fish collection, and (2) to provide the fisheries researchers on the vessel with an updated species list, the documentation of range extensions and descriptions of the rarer species encountered. Because of space and transport limitations, preparations for the surveys included a review of the species likely to be encountered, using existing guides to the area (Bianchi 1986; Bianchi et al. 1993; Schneider 1990) to determine which species should be collected. Decisions were made on which of the known species to collect based on presence or absence from the existing fish collection at SAIAB and on taxonomic status. Those species that required further taxonomic study had higher priority than well-known species. For the lesser-known, deep-water species, efforts were made to obtain representative samples of as many as possible, but concentrating on specimens that were in good condition. Any specimens that were not identified on board were retained for later study. A complete list of all species collected is appended to this paper, together with a summary of trawl station data for cross reference. On the basis of the Clofeta synopsis and some more recent literature, we record range extensions (Table 1) of 34 fish species from more northerly waters, two (Galeichthys feliceps and Careproctus albescens) from more southerly waters and the first eastern Atlantic record of the apparently widespread eel Dysommina rugosa. A new morid species collected, Physiculus cyanostrophus Anderson & Tweddle 2002, was described earlier. Colour plates in this paper illustrate a small selection of species caught during the surveys, including some of the species discussed in the text. MATERIALS AND METHODS The trawl surveys were designed to obtain biomass estimates using the swept area method (Sparre & Venema 1998) and covered the entire length of the Angolan continental shelf from 20 to 800 m in depth. A total of 342 stations (summarised in Appendix A) were fished in the 2001 and 2002 surveys, with trawl bottom time usually of 30 minutes unless interrupted by hard ground. Samples were taken from each trawl by the fisheries scientists on board for sorting, weighing and measurement. The fishes in this sample were checked and if there was any uncertainty over identification, the fishes were examined further in the vessel's laboratory, using available identification keys on board. In addition to the sample taken, each trawl haul was examined on deck for any unusual species that were present. Using the target species list prepared in advance, up to ten good specimens were collected for each of the species and preserved in 10% formalin. Rare, unidentified or questionable specimens were also retained. Most specimens were identified on board, though some species are the subject of further study, as discussed below. Each fish kept was labelled with a code number written in pencil on plastic paper, which was inserted in the mouth or gillcover, cross-referenced with the field notes and station number. Full collection details were added later from the vessel's trawl records at SAIAB where the collection of 1704 specimens of 277 species was transferred to 60% propanol or 70% ethanol and catalogued. The full species list is given in Appendix B. When time permitted and specimens of the various species were available in good condition, photographs were taken, generally against a standard yellow background, using the vessel's Olympus digital camera. Some photographs were also taken on slide film. Most photographed specimens were kept and preserved, with the specimen and photo cross-referenced, though some larger and/or commoner species were not retained. In addition to the 2001 and 2002 survey collections, specimens of the uncommon Lestidiops cadenati, Cataetyx bruuni and Pachycara crossacanthum were obtained by Dr Tomio Iwamoto, California Academy of Sciences (CAS), during the UNDP/NORAD cruise of March-April 2005. Data from these specimens were incorporated into our accounts, with the station data added to Appendix A. Counts and measurements in the species accounts follow Hubbs & Lagler (1947), Fernholm & Hubbs (1981) and McMillan & Wisner (1984) for hagfishes, Robins & Robins (1970) and Smith & Kanazawa (1977) for eels, Bradbury (1988, 1999) for batfishes, Pietsch (1974) and Bertelsen and Krefft (1988) for anglerfishes, Eschmeyer (1969) for scorpionfishes, Anderson (1994) for zoarcids and Robins (1960, 1962) for ophidioids. Counts of vertebrae and unpaired fin rays of elongate fishes were taken from radiographs. Abbreviation used are as follows: SL - standard length; TL - total length; HL - head length; Pelv. - pelvic fin rays. Authorities for original descriptions are available in Eschmeyer's on-line Catalog of Fishes, http://www. calacademy.org/research/ichthyology/catalog/ fishcatmain.asp, and are not included in the reference list here. SYSTEMATICS MYXINIDAE Myxine ios Fernholm, 1981 This hagfish was the first to be captured in West African waters, and is known to occur from off Western Sahara to Iceland (Fernholm & Vladykov 1984). Fernholm (1981) found the species exists in two populations, a northern one with a light grey body and whitish head, and an African one with a uniformly darker grey colour. A single specimen, 303 mm TL, of the African population was collected off northern Angola in 700 m at station 2799 during the 2002 cruise (Appendix B). It agrees with the original description in every respect except that it has 101 slime pores, compared with 103-116 for the West African population, and slight differences in a few morphometric features. Head and body uniformly dark purplish-grey. Ventral part of snout, mouth, barbels, gill apertures, slime pores, cloaca and distal half of ventral fin fold white. Proportions as percent total length: prebranchial length 30; trunk length 57; tail length 13; body width 4.8; body depth including fin fold: 6.5; body depth excluding fin fold: 5.6; body depth at cloaca: 5.3; tail depth 5.7; first barbel length 1.3; second barbel length 1.3; third barbel length 1.5. Slime pore counts, left side: prebranchial 29, trunk 61, tail 11 = 101 total. Seven gill pouches on each side. Tooth counts: cusps on multicuspids 2/2 each side; unicuspids, right side: outer row 9, inner row 9; unicuspids left side: outer row 10, inner row 9; total cusps 45. TORPEDINIDAE Torpedo bauchota This electric ray was described from two juveniles taken off the Congo and Senegal, depths unknown. Other (unpublished) specimens are known from Ivory Coast and Senegal (B. Seret pers. comm., 2003). Two juvenile females, 148-150 mm TL, were taken off northern Angola during the 2001 cruise in 24 m (Appendix B). Anterior edge of disk straight, pelvic fins gently rounded. Characteristic black variegations snaking around disk; these wider, therefore disk darker, in 148 mm specimen than in the larger individuals. Pale spots surrounded by thin black lineations in this specimen, spots paler (whitish), without lineations in 150 mm specimen. Spiracle surrounded by 9-10 finger-like papillae. Molariform teeth in four rows anteriorly in both jaws. Proportions as percent disk width: total length 1.4-1.5; disk length 83-87; snout to origin of first dorsal fin 93-99; snout to origin of second dorsal fin 1.0; interdorsal distance 3.7-4.1; first dorsal fin base 9.3-9.8; second dorsal fin base 7.5-7.7; transverse spiracle diameter 2.7-2.9; interspiracular width 7.0- 7.9; interorbital width 4.8-5.0; mouth width 9.3-9.4; internasal width 5.8-5.9; snout to mouth 13; first gill slit interdistance 25; fifth gill slit interdistance 23-24. SYNAPHOBRANCHIDAE Dysommina rugosa Ginsburg 1951 This cutthroat eel has been reported reliably from upper slope depths in the northwestern Atlantic and Hawaii. A single 362 mm TL specimen was taken off northern Angola in 536-542 m during the 2002 cruise (Appendix B) that agrees with North Atlantic specimens (Robins & Robins 1989). Total vertebrae 133; predorsal vertebrae 11; Dorsal fin rays (D) 314; Anal fin rays (A) 290; Pectoral fin rays (P) 16; Caudal fin rays (C) 15. Pores: preoperculomandibular 5 (pore in position five, under posterior margin of eye, not open on both sides); infraorbital 5; supraorbital 3. No supratemporal commissure. Vomer with four caniniform teeth. Proportions as percent TL: predorsal length 17; preanal 28; body depth at anus 5.9; head length 13; head width 3.3. Proportions as percent head length (HL): head width 26; snout 28; eye diameter 13; jaw length 45; gill opening 16; interbranchial distance 12; pectoral-fin 24; interorbital 15. Colour uniformly dark chocolate brown. OPHICHTHIDAE Pisodonophis semicinctus (Richardson 1848) This distinctive snake eel was originally described as Ophisurus semicinctus Richardson 1848. This name, however, is a junior homonym of Ophisurus semicinctus Lay & Bennett 1839, another snake eel now placed in the genus Leiuranus Bleeker (McCosker 1977). Richardson's name has been wrongly treated as valid by authors as Pisodonophis semicinctus. Also, the species does not belong to Pisodonophis sensu stricto, thus a replacement name is required (J. McCosker, pers. comm.). This eel ranges from the Mediterranean coast of Algeria to Angola at inner shelf depths (Bauchot 1986). Three specimens, 616-813 mm TL, were taken during the cruises off northern and central Angola in 25-40 m (Appendix B). Recent descriptive literature for the species includes Blache et al. (1970), Blache & Saldanha (1972), Saldanha (1982) and Bauchot (1986), summarised below. Total vertebrae 155-162. Lateral line pores: prepectoral 10-12; preanal 53-59. Pectoral-fin rays 10- 1. Caudal fin absent, tip of tail a stiff lobe. Proportions as percent total length (TL) (Blache & Saldanha 1972): predorsal length 6.7-8.9; preanal 32-43; head 11- 12; body depth 3.3-4.3. Proportions as percent HL: predorsal length 63-79; body depth 29-40; snout 19-21; eye diameter 6.6-10.0; interbranchial distance 18-20; pectoral-fin 28-33. Head cuneiform in lateral view when mouth closed. Dorsal and anal fins well developed, dorsal-fin origin distinctly in advance of gill opening. Background colour yellow, lighter ventrally, with 14-18 dark brown saddles from nape to tail tip. Pectoral fin yellow. Head with large dark saddle blotch and smaller black spots. CONGRIDAE Xenomystax congroides This eel is chiefly known from the Gulf of Mexico to off northern Brazil in depths of about 150-500 m, but is also known in the eastern Atlantic from 10 specimens taken from off Liberia to Congo in 156-400 m (Smith 1989). Two specimens were taken off northern Angola during the 2001 cruise at depths of 327-355 m (Appendix B). One specimen (SAIAB 64867, 445 mm TL) has lost the end of its tail (172 total vertebrae) and has a regenerated pseudocaudal fin. Both specimens agree well with the Gulf of Guinea population in the original description but have one less supraorbital pore. Predorsal vertebrae 4; preanal vertebrae 39- 40; precaudal vertebrae 60-61; total vertebrae 216 (undamaged specimen). Branchiostegal rays 10. Pectoral-fin rays 12-13. Pores: lateral line to anus 36-37; preoperculomandibular 14; infraorbital 5 + 0 + 3; supraorbital 1 + 4; supratemporal commissure 3. Proportions as percent TL (undamaged specimen): predorsal 12; preanal 34; head 14. Proportions as percent preanal length (both specimens): predorsal 34- 35; head length 41; head width 9.0-10.1; depth at anus 11; pectoral fin 9.2-10.1. Proportions as percent HL: snout 32; eye 11; upper jaw 55-56; gill opening 9.2-10.3; interbranchial 10; pectoral fin 23-25. Body and tail brownish dorsally with scattered melanophores. Lateral line area pale. Ventral surface pale yellowish. Edges of vertical fins posteriorly and gut black. Intermaxillary tooth patch with three large canines anteriorly (Smith 1989, fig. 598). PARALEPIDIDAE Lestidiops cadenati (Maul 1962) This barracudina was described from a single beach-cast juvenile, 82.5 mm SL, found near Dakar, Senegal in 1953 (Maul 1962). Subsequent references that we found all refer to the holotype (Blache et al. 1970; Post 1972, 1991). Nineteen specimens, 210-233 mm SL, were collected during the two cruises off northern Angola at five stations ranging in bottom depth from 259 m to 364 m (Appendix B). The 2005 CAS collection contains seven specimens 217-318 mm SL from stations 3630, 3656, 3675, 3698 and 3709 (Appendix A). Barracudinas are considered coastal, mesopelagic fishes (Rofen 1966). Vertebral and caudal-fin counts were taken from radiographs of 15 specimens. Other counts and measurements were taken from all 26 fish where possible. Vertebrae 36-40 + 41-45 = 79-84; D 9-11 (usually 10); A 25-27; P 11-12; Pelv. 9; C xiii-xiv + 10-11 + 9 + ix-xvii (19-20 principals); lateral line sections 130-140; gill rakers 4-13 + 22-29 = 26-41; branchiostegal rays 5; pseudobranch filaments 16-19. Proportions as percent SL: head length 22-24; head width 4.1-4.8; head depth 6.4-7.5; postorbital length 8.4-9.3; upper jaw length 8.410.2; lower jaw length 11-13; predorsal length 60-63; preanal length 79-80; prepelvic length 56-60; pectoral-fin length 7.9-9.3; pelvic-fin length 5.2-5.9; body depth at dorsal-fin origin 5.2-7.3; body depth at anal-fin origin 5.0-6.1; gill slit length 11-12; caudal peduncle length 5.2-7.2; caudal peduncle least depth 2.5-2.8; dorsal-fin base 4.5-5.6; anal-fin base 14-15. Proportions as percent HL: head width 19-21; head depth 28-34; postorbital length 38-41; upper jaw length 38-43; lower jaw length 50-57; pectoral base depth 9.2-11.6; pectoral-fin length 35-42; pelvic-fin length 23-25; snout length 44-46; orbit diameter 16-18; gill slit length 50-52; bony interorbital width 9.2-11.0. Head depressed, snout long and tapering to tip of upwardly-protruding lower jaw that fits into the median diastema of the upper jaw. Over 60 tiny teeth on each premaxilla. Maxilla closely joined to premaxilla, edentate, as is vomer. Palatines with 6-9 retrorse, caniniform teeth in anterior half, usually arranged in groups of two, with innermost the larger; single row of 8-19 smaller, retrorse teeth posteriorly. Dentary with double row of teeth posterior to symphysis, outer teeth small, retrorse, inner teeth long, dagger-like. Eye large, rounded, with adipose eyelid covering anterior and posterior third of pupil. Gill cavity black, gill arches and pseudobranch filaments elongate. Gill rakers reduced to placodes bearing 1-4 strong denticles, with 4-13 placodes on epibranchial, 12-17 on ceratobranchial and 9-12 on hypobranchial. Operculum and suborbital regions with complex, radiating pattern of sensory canals. Body strongly laterally compressed. Ventral carinas (adipose keels) between pectoral and pelvic fins and pelvics and anal fin dotted with tiny melanophores. Lateral line scales (sections) one per myomere anteriorly; three upper and three lower pores posteriorly on each scale to mid-anal fin (Fig. 2; Maul 1962, Fig. 5; compare with Rofen 1966, figs. 101, 102). Scales abruptly diminish in size near posterior end of anal fin, becoming two per myomere and with a single dorsal and single ventral pore. Lateral line scales extend to end of caudal peduncle where they are three per myomere, with a few scales turning dorsally right at end. Anus on vertical through dorsal-fin rays 4-8, center of anus situated 20.8-26.2% HL posterior to pelvic-fin insertion. Fins relatively small; pectorals wedge-shaped, dorsalmost five rays longest. Anal fin deeply notched anteriorly, rays 4-6 longest. Dorsal fin low, usually of 10 rays, its origin one eye diameter posterior to vertical through pelvic-fin origin. Colouration: dorsum dusky brown, pale silvery ventrally. Eye and opercular areas dark blue. Peritoneal pigment patches absent in our material. Fins of largest specimen (CAS 222645; 318 mm SL) dusky. OPHIDIIDAE Ophidion lozanoi Matallans 1990 This cusk-eel was described from two specimens taken off Western Sahara (holotype) and Senegal (paratype). No further published records are known (J. Matallans pers. comm.), but Robins (1999: 40) listed it from Spain as well. Twenty-one specimens, 117-208 mm SL, were collected during the 2002 cruise off central and northern Angola (Porto Amboim to the Congo River mouth) at depths of 40-259 m (Appendix B). Counts of vertebrae and unpaired fin-rays were taken from six radiographed specimens. Other counts and measurements were taken from 15 specimens 144-208 mm SL (the two types were 130 & 138 mm SL). Vertebrae 15-16 + 51-53 = 67-69; D 130-147; A 108-119; P 24-25; C 9; Pelv. 2; lower limb gill rakers 4; branchiostegal rays 7; pseudobranch filaments 4-5. Proportions as percent SL: head length 21-22; head width 8.7-11.3; head depth 13-14; predorsal length 26-29; preanal length 40-41; prepelvic length 5.2-6.4; gnathoproctal length 37-38; lateral line length 88-90; body depth at occiput 14-25; body depth at dorsal-fin origin 14-15; body depth at anal-fin origin 13-14; gill slit length 12-15; occipital length 13-14; postorbital length 12-13. Proportions as percent HL: upper jaw length 49-54; lower jaw length 55-60; pectoral base depth 18-19; pectoral-fin length 43-46; pelvic-fin length 33-53; snout length 19-21; orbit diameter 24-27; gill slit length 56-68; bony interorbital width 14-17; occipital length 61-63; postorbital length 57-59. Anterodorsal edge of snout high as result of large rostral spine; tip of snout vertical (Fig. X). Tail laterally compressed, lateral line coursing along its dorsal third, extending to posterior seventh of total length. First dorsal-fin pterygiophore inserted between vertebrae 6 and 7. Epibranchial tooth plates 2-3, four elongate gill rakers on ceratobranchial. Vomerine teeth 28-51, in triangular or rhomboidal patch. Palatine teeth in three (smallest fish) to five (largest) irregular rows. No pyloric caeca. Peritoneum and orobranchial chamber black, mouth dusky toward sides. Body greyish-brown dorsally with yellowish tinges ventrally in smaller specimens. BYTHITIDAE Cataetyx bruuni (Nielsen & Nybelin 1963) This rare brotula was originally described in the genus Oculospinus Koefoed 1927 from three specimens taken off Angola at depths of 235-510 m, but was assigned to Cataetyx by Cohen (1981). No other descriptive records were found, but Nielsen (1990) mentioned there were a 'few specimens from the Gulf of Guinea'. We found that these are 27 specimens in four lots in the Museum de La Rochelle, France, and the National Museum of Natural History, Washington, D.C., USA. They range from Senegal to Gabon at depths of 329-1355 m. Three specimens were collected during the 2002 cruise off central Angola (north of Benguela to off Cabo Ledo) at depths of 504-707 m (Appendix B), an adult male 92 mm SL and two gravid females 87-100 mm SL. The CAS 2005 collection is of 21 specimens 65-122 mm SL from stations 3601, 3622, 3641, 3649 and 3656 at depths of 604-809 m (Appendix A). Vertebrae 15-16 + 48-50 = 61-66; D 107-120; A 81-89; P 26-28; C 10; Pelv. 1; lower limb gill rakers 3; branchiostegal rays 9; pseudobranch absent. Proportions as percent SL: head length 23-26; head width 10-14; head depth 11-13; predorsal length 31-34; preanal length 47-51; prepelvic length 19-21; gnathoproctal length 42-46; body depth at occiput 11- 14; body depth at dorsal-fin origin 11-17; body depth at anal-fin origin 11-13; gill slit length 15-16; postorbital length 14-15; pectoral fin length 13-15. Proportions as percent HL: upper jaw length 43-49; lower jaw length 49-56; pectoral base depth 21-26; pectoral fin length 56-63; pelvic fin length 31-42; snout length 21-23; orbit diameter 19-20; gill slit length 63-66; bony interorbital width 14-16; postorbital length 56-60. Head ovoid, depressed; tip of snout steeply sloping (Fig. 3). Tail laterally compressed; lateral line not evident. First dorsal-fin pterygiophore inserted between vertebrae 7 and 8. Rudimentary gill rakers on epibranchials 2-3; three elongate rakers on upper part of ceratobranchial and seven rudiments ventrally. Vomerine teeth 16-27, in two arcing rows. Palatine teeth in three irregular rows. No pyloric caeca. Opercle with stout, V-shaped spine arrangement; uppermost projecting posteriad, lower projecting ventroposteriad. Skin flap on opercle above posteriorly-directed spine ending in a wide pore. Cleithrum with posteriorly-directed spine just above pectoral base. Posteriorly-directed preorbital spine directly behind posterior nostril. Pelvic fins inserted under opercle. Top of head and body brown, head darker. Opercle, lower side of head and area under eye blue. Cheeks yellowish. Orobranchial chamber and peritoneum black. OGCOCEPHALIDAE Dibranchus atlanticus Peters 1876 This batfish is known in the western Atlantic from Canada to Brazil and in the eastern Atlantic from the Gulf of Guinea to Angola in depths of about 100-1260 m (Bradbury 1999). Twenty-six specimens were taken off central and northern Angola during the cruises at depths of 112-614 m (Appendix B). The specimens agree with Bradbury's detailed description, but most have very large tubercles more like those of D. tremendus Bradbury 1999 than typical D. atlanticus (Bradbury 1999, Fig. 8). The central spines of our specimens are not greatly elongated on the tubercles of the tail, as in D. tremendus. All tubercles are decorated with spinules, however, and this, with the small sizes and shallow collection depths, identify the species as D. atlanticus. Counts and body proportions from 10 specimens 81-105 mm SL: D 5-6; A 4; P 13-14; Pelv. 5. Neuromast counts: preopercular 2; subopercular 5-6; tail 10-11. Proportions as percent SL: disk margin length 42-44; skull length 28-31; cranium width 20-22; eye width 9.7-10.7; distance jaw to anus 49-53; jaw to anal fin origin 71-74; predorsal length 58-60; interorbital width 8.6-9.4; jaw length 11-12; mouth width 14-16. DICERATIIDAE Diceratias pileatus Uwate 1979 This double anglerfish is known in the literature from 45 specimens ranging across the tropical Atlantic from the Bahamas to the Gulf of Guinea (Uwate 1979; Fujii 1983). Pietsch & Randall (1987) reported the first occurrence of the species outside the Atlantic (off Hawaii), which is the largest specimen known at 275 mm SL. Twenty-two female specimens, 20-114 mm SL, from nine stations were collected during our cruises off Angola (Appendix B). This species is mesopelagic and bottom depths ranged from 446-769 m. Following counts and proportions taken from the ten best specimens, 20-65 mm SL: D 6-7; A 4; P 12-13; C 9; vomerine teeth 0-8 (absent in 20 mm fish); upper jaw teeth 9-17; lower jaw teeth 14-22. Proportions as percent SL: predorsal length 76-79; preanal length 86- 89; body depth 59-62; illicial length 36-40; head length 37-40; head depth 57-59; head width 33-36; lower jaw length 44-47. Esca a simple bulb with a small cap (absent in 20 mm fish). Body solid black, covered in minute spinules. HIMANTOLOPHIDAE Himantolophus paucifilosus This footballfish was described from 17 females taken at mesopelagic depths across the tropical Atlantic. A single female, 147 mm SL, was taken off central Angola in 2002, bottom depth 672 m (Appendix B). Counts and proportions as percent SL: D 5; A 4; P 16; C 9; head length 39; head depth 56; predorsal length 87; preanal length 87; body depth 65; illicial length 46; diameter escal bulb 5.5; length distal escal appendage 2.0; anterior appendage absent; length posterior appendage 9.5; length illicial appendage 15. Two illicial appendages just below esca, each with an unbrancing filament. Colour solid black. SCORPAENIDAE Neomerinthe folgori (Postel & Roux 1964) This rare scorpionfish was described from a single specimen, 287 mm SL, taken off the Cape Verde Islands in 180-200 m. We found that only two other specimens have been reported, one from Mauritania in 310 m (Cervignon 1960, reported as Scorpaena sp. (Eschmeyer 1969)) and another from Namibia in about 200 m (Penrith 1980; Eschmeyer 1986). The species was not mentioned by Blache et al. (1970). A gravid female, apparently the largest specimen known at 417 mm SL, was collected off northern Angola during the 2001 cruise in 322-324 m (Appendix B). It agrees with published descriptions, but as it is much larger than the other three (to 340 mm), has significantly shorter predorsal and snout lengths and fewer gill rakers (lowermost somewhat coalesced); variation in other morphometric features is insignificant. D XII, 11; A III, 5; P 17; C 16; Pelv. I, 5; gill rakers 8 + 8; tubed lateral line scales 28; lateral line scale rows 78. Proportions as percent SL: head length 45; head width 25; head depth 32; body depth 36; predorsal length 35; preanal length 73; prepelvic length 36; pectoral-fin length 23. Proportions as percent HL: head width 57; head depth 71; postorbital length 55; orbit diameter 19; snout length 31; upper jaw length 47; pectoral-fin length 51. Preorbital bone with two spinous lobes over upper jaw, with 11 lateral points and five cirri. Suborbital ridge with 34 small points along centre line. First preopercular spine longest, second small (absent on left side), third and fourth present, fifth a mere nub under skin. Supplemental preopercular spine present. Postorbital ridge with upper posttemporal spine embedded (unexposed); supracleithral, lower posttemporal, pterotic and sphenotic spines present, letter two with several small points. Scales on body ctenoid, 78 lateral scale rows (the count by Postel and Roux 1964, of 85 in the holotype was corrected by Poss & Duhamel 1991, to 77). Cirri on head weakly developed, mostly absent. Background colouration yellowish with black and brownish variegations. Four variegated black bands on body. Caudal fin with narrow bands, posterior margin without wide black band as in holotype. LIPARIDAE Careproctus albescens Barnard 1927 Careproctus griseldea Lloris 1982 Vertebrae 11 + 51 = 62; D 55; A 49; P 26 + 11 = 37; C 11; branchiostegal rays 6. Proportions as percent SL: Predorsal length 23; preanal length 36; head length 23; head width 14; head depth 18; body depth 20; gill slit length 8.2; pectoral-fin length 19. Proportions as percent HL: head width 60; head depth 77; upper jaw length 37; lower jaw length 43; pectoral base depth 32; pectoral-fin length 82; body depth 86; disk width 23; disk length 26; snout length 32; eye diameter 22; gill slit length 36; bony interorbital width 19. Colour uniformly pale grey anteriorly grading into blackish tail. Peritoneum dusky (densely dotted with melanophores, typical of adult, shallow-dwelling Careproctus species). Teeth mostly simple, retrorse, a few in inner rows of lower jaw trilobed (tips triangular). CARISTIIDAE The manefishes are in great need of systematic revisionary study. No keys to all the species exist (but see below) and a few undescribed genera and species are known in collections (M. Leiby pers. comm. 2002). Until recently, two subgenera of Caristius Gill & Smith 1905 were usually recognised, Platyberyx Zugmayer 1911, with a high, curved lateral line and vomerine and palatine teeth, and Caristius Gill & Smith 1905, with lateral line straight or absent and with or without vomerine or palatine teeth. The manefishes were not considered by Blache et al. (1970), but Post (1991) lists four species for the Clofeta area, two Platyberyx (P. opalescens and P. groenlandicus) and two Caristius (C. macropus and C. maderensis). Trunov et al. (2006) erected Paracaristius for C. maderensis (type species) and a new southern hemisphere species (P. heemstrai), but did not comment on the generic status of C. macropus. Nine specimens, 56-173 mm SL, of a manefish were taken during our cruises off central and northern Angola at bottom depths of 601-776 m (Appendix B). Caristius macropus, originally described from Japan (Bellotti 1903) and well known in the Pacific, has been reported from the Atlantic a few times (e.g., Fraser-Brunner 1931; Parin et al. 1974, but not Norman 1930 [see Maul 1949: 26 for corrected counts]). We distinguish our species from C. macropus, and therefore identify it as P. maderensis, on the basis of its lower counts and lack of vomerine and palatine teeth. The material agrees well with Mauls' original description and that of Trunov (1981), each based on one specimen. We offer here a tentative key gleaned from other Caristius specimens in the SAIAB Fish Collection, the literature and an unpublished key of R. Britz, Natural History Museum, London. KEY TO ATLANTIC SPECIES OF MANEFISHES 1A. Lateral line high on dorsum, arched ................... 2 2A. D 28-31; A 17-19 ................... Platyberyx opalescens 3A. Rear margin of upper jaw not reaching beyond middle of eye; vomerine and palatine teeth absent; D 26-30; A 15-17; P 15-17; vert. 32-33 ...... ............................................. Paracaristius maderensis 3B. Rear margin of upper jaw reaches beyond middle of eye; vomerine and palatine teeth present; D 33-37; A 21-23; P 17-20; vert. 39-40 ..................................................... Caristius macropus Paracaristius maderensis (Maul 1949) Counts and proportions from Angola material (vertebral counts from radiographs of five specimens): Vertebrae 15 + 17-18 = 32-33; D 27-28; A 17-18; P 16-17; C 17 (principal rays); Pelv. 6; gill rakers 6-8 + 14 = 20-22; branchiostegal rays 7; pseudobranch filaments 11-20. Proportions as percent SL: head length 30-36; head width 13-15; upper jaw length 11-12; lower jaw length 15-20; predorsal length 8.8-9.5; preanal length 56-57; prepelvic length 25-30; length longest dorsal ray 58-81; length longest anal ray 25-43; pectoral length 26-31; pelvic length 59-61; body depth 47-48. Proportions as percent HL: head width 41-44; upper jaw length 32- 36; lower jaw length 49-56; pectoral base depth 21-23; snout length 11-13; pupil diameter 31-36. ZOARCIDAE Pachycara crossacanthum Anderson 1989 This rare eelpout was described from six specimens, 227-378 mm SL, collected by French researchers in fish traps set off Senegal and Gabon in depths of 900-1050 m. No other specimens have been reported. Four late juvenile females, 137-155 mm SL, were collected during the two cruises off central and southern Angola at two stations in depths of 672-783 m (Appendix B). The CAS 2005 collection is of four specimens 122-265 mm SL from stations 3609, 3676 and 3684 taken at depths of 725-891 m (Appendix A). Vertebrae and median fin rays were counted from radiographs of all eight fish. Vertebrae 25-28 + 76-81 = 103-107; D 98-101; A 80- 82; C 11-12; P 17; Pelv. 3; vomerine teeth 5-9; palatine teeth 7-13; gill rakers 3-4 + 13-14; branchiostegal rays 6; pseudobranch filaments 4-5. Proportions as percent SL: head length 17-19; head width 8.3-8.7; head depth 8.8-9.6; pectoral-fin length 10-11; predorsal length 19- 22; preanal length 41-44; body depth 9.4-10.7; gill slit length 6.6-7.5; caudal-fin length 3.0-3.7. Proportions as percent HL: head width 44-50; head depth 50-54; upper jaw length 31-38; pectoral-fin length 58-66; snout length 18-22; eye diameter 18-24; gill slit length 38-42; interorbital width 7.9-11.2; interpupillary width 23-27; pelvic-fin length 13-21; pectoral base/length ratio 42-47. The four SAIAB and two CAS juveniles agree in every respect with the larger specimens but have slightly longer heads, pectoral fins and gill slits. Eight preoperculomandibular pores, seven suborbitals, two supraorbitals, four postorbitals (except CAS 222451 and 222647 with three), and no interorbital or occipitals. Only dorsalmost 5-7 gill rakers on lower limb furcate in the six juveniles (Anderson, 1989, Fig. 8). Body lateral lines originating just posterior to postorbital pore four, with mediolateral and ventral branches diverging just above anterior quarter of pectoral fin. Dorsal fin origin associated with vertebra four, with no free pterygiophores. Anal fin origin associated with ultimate precaudal vertebra, with 2-3 ray-bearing pterygiophores inserted anterior to haemal spine of first caudal vertebra in all four fish. Caudal fin with two epural and 9-10 hypural rays. ACKNOWLEDGEMENTS The authors are indebted to the Institute of Marine Research, Bergen, Norway, particularly the late Guillermo Burgos, for the invitation to take part in the cruises and for logistical support and specimen shipment. We also thank Alvheim Oddgeir and the IMR for permission to use photographs from the IMR's on-board identification digital photograph collection. Scientists and crew from both Norway and Angola were friendly and supportive. Particular thanks go to cruise leaders Åge Høines, Sigborn Mehl, Diana Zaera-Perez and Bjørn Erik Axelsen, and to scientists Haraldur Einarsson, Magne Olsen and Maria Sardinha for their assistance and friendship. SAIAB, a national facility of the National Research Foundation in South Africa provided support to both authors for the analysis and paper preparation. Elaine Heemstra rendered all text drawings. Specimens were identified and curatorial needs provided by David Smith, Smithsonian Institute, Bernard Seret and Martine Desoutter, Museum National d'Histoire Naturelle, and John McCosker and Tomio Iwamoto, California Academy of Sciences. Phillip Heemstra, John McCosker and Stuart Poss commented on the manuscript. LITERATURE CITED
APPENDIX A. A summary of station data for the 2001 and 2002 cruises. The 2005 survey stations from which specimens were provided by T. Iwamoto are also included.
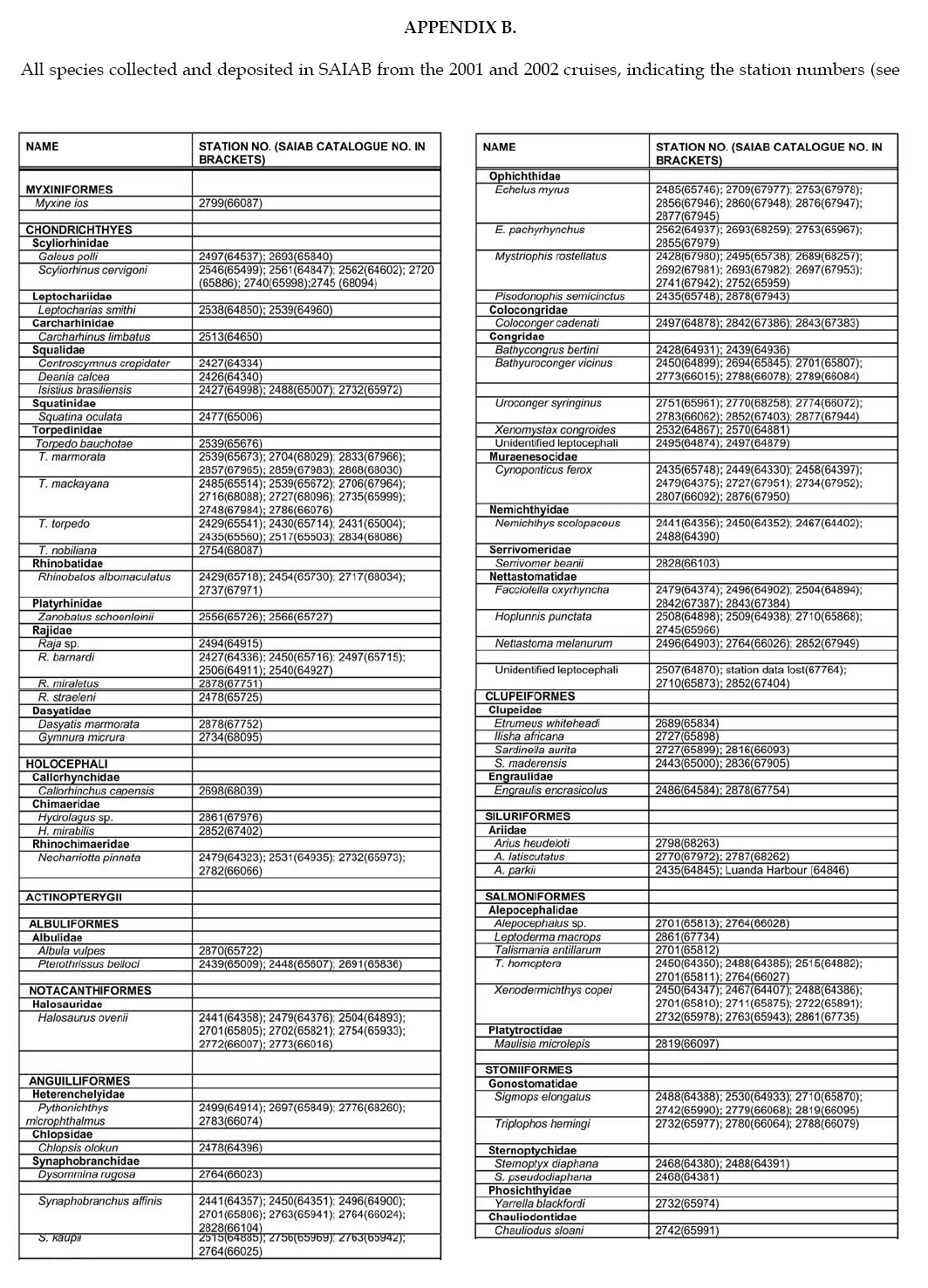
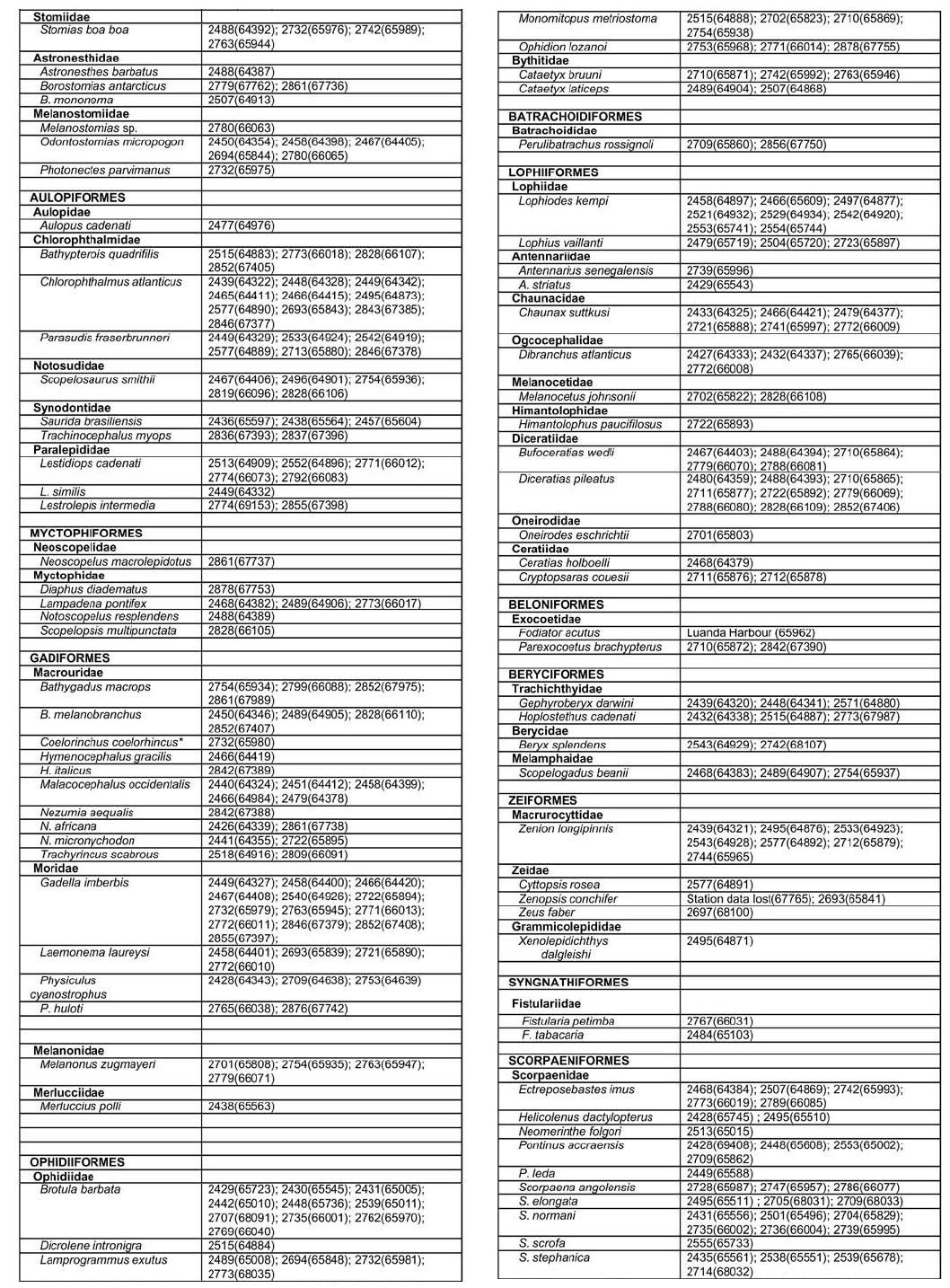
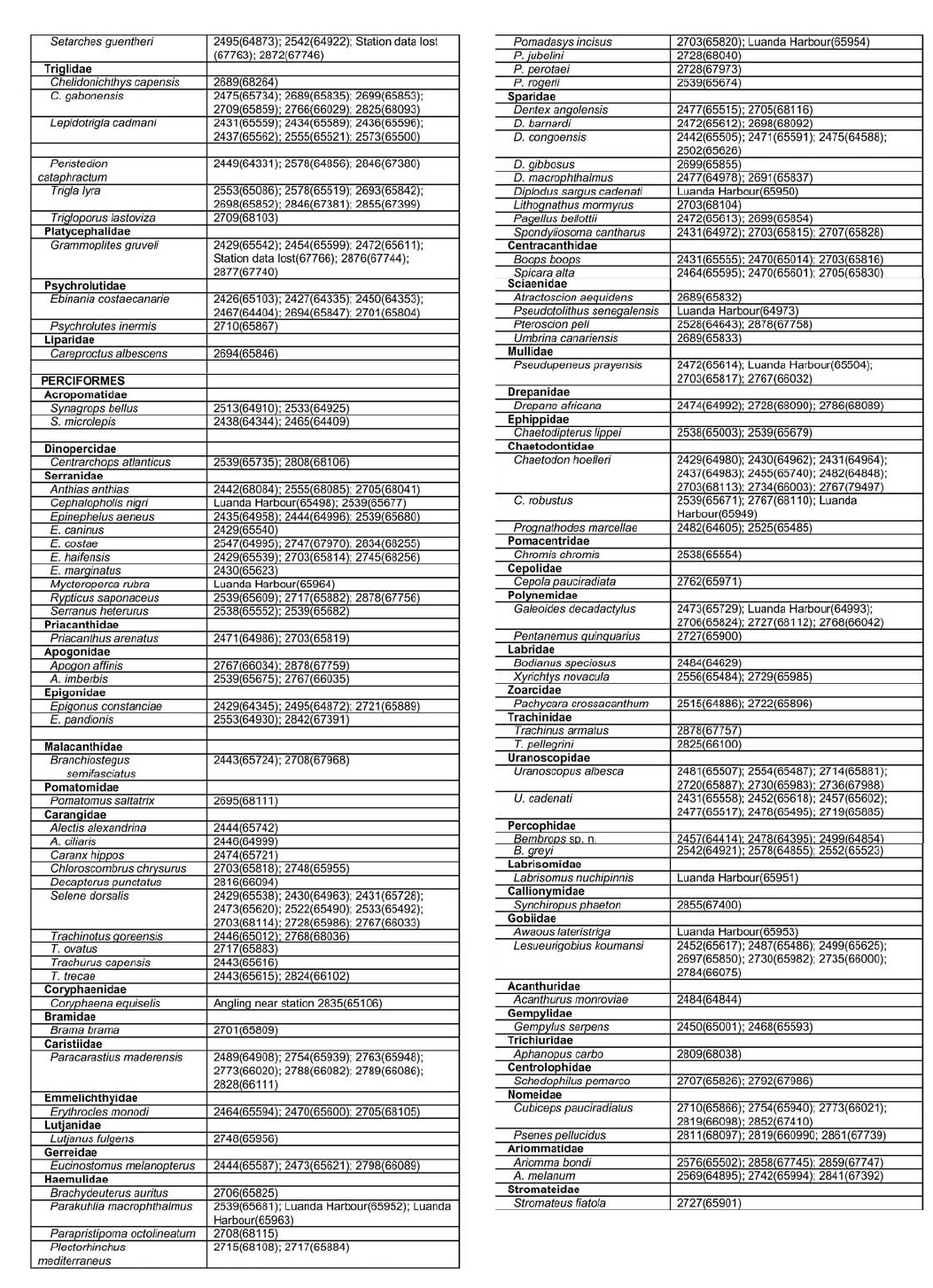
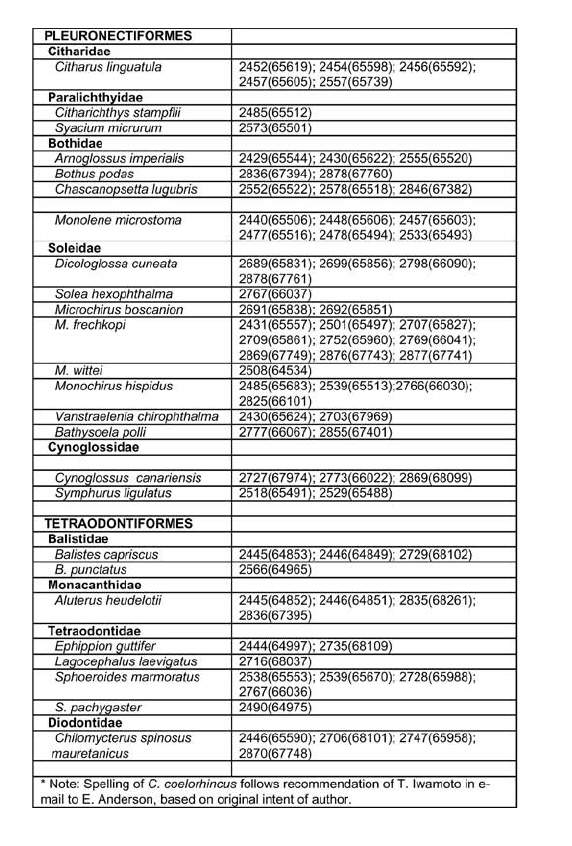
Appendix B: part 1, 2, 3, 4 © 2008 The South African Institute for Aquatic Biodiversity, Grahamstown, South Africa The following images related to this document are available:Photo images[sm08001p3.jpg] [sm08001p2.jpg] [sm08001f1.jpg] [sm08001p1.jpg] [sm08001f4.jpg] [sm08001ab3.jpg] [sm08001aa.jpg] [sm08001f3.jpg] [sm08001ab1.jpg] [sm08001ab4.jpg] [sm08001t1.jpg] [sm08001f2.jpg] [sm08001f5.jpg] [sm08001ab2.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}