 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
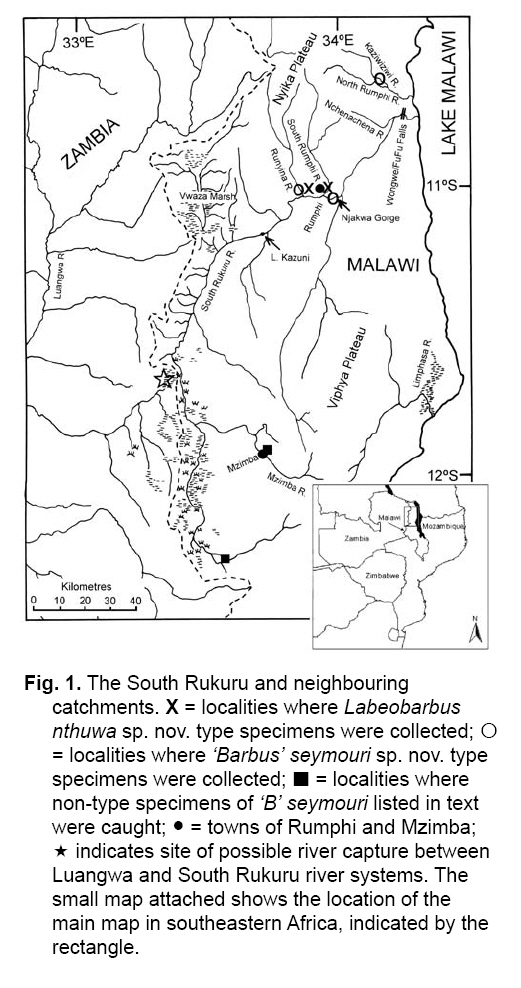
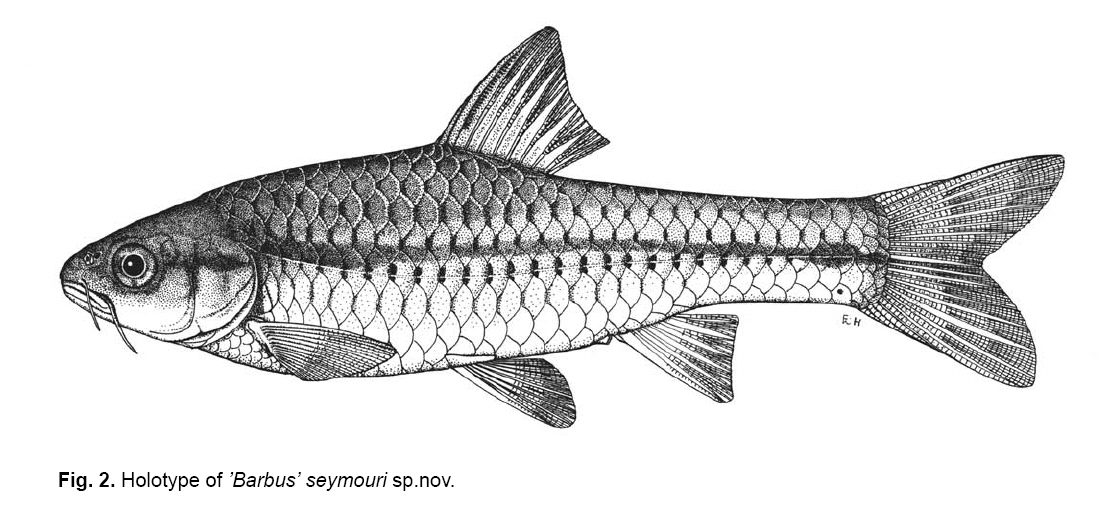
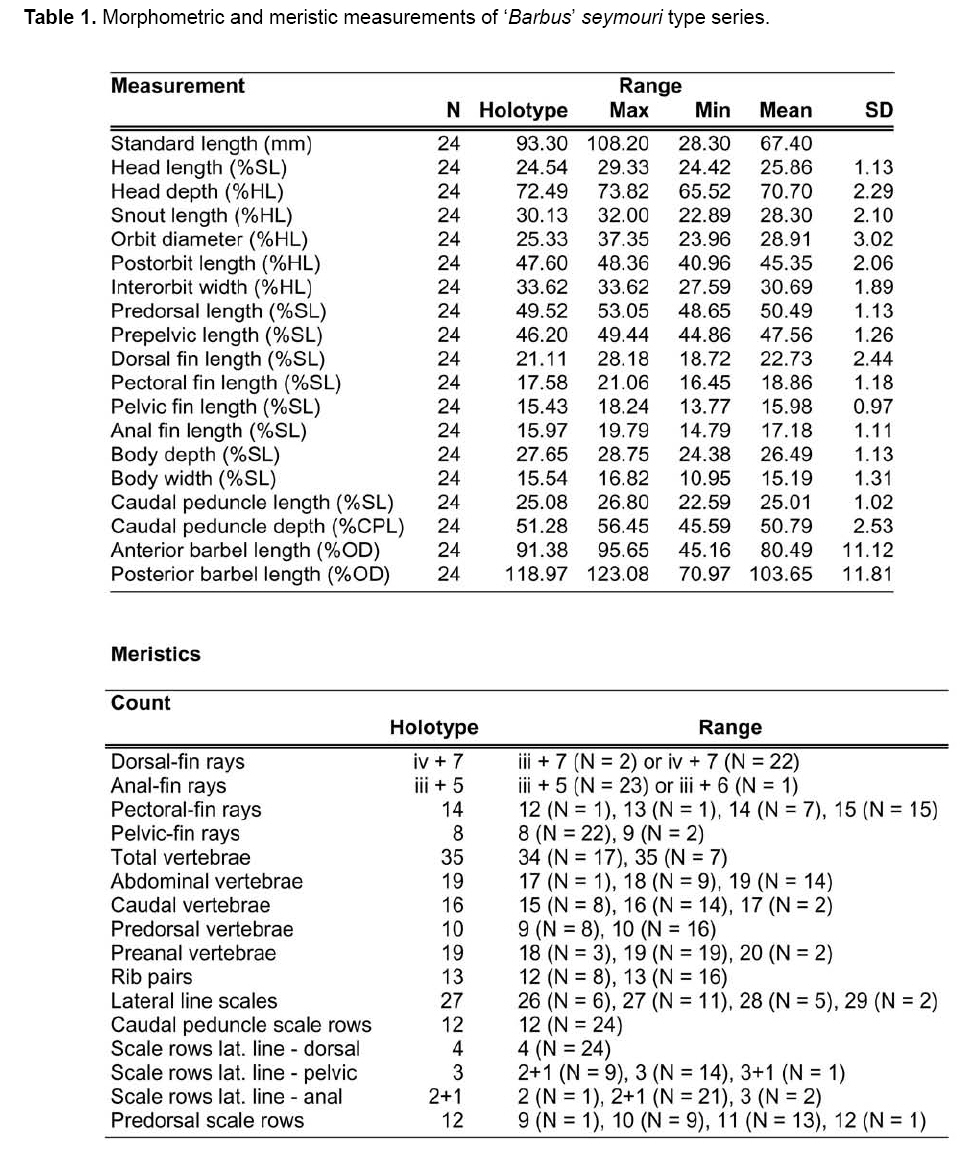
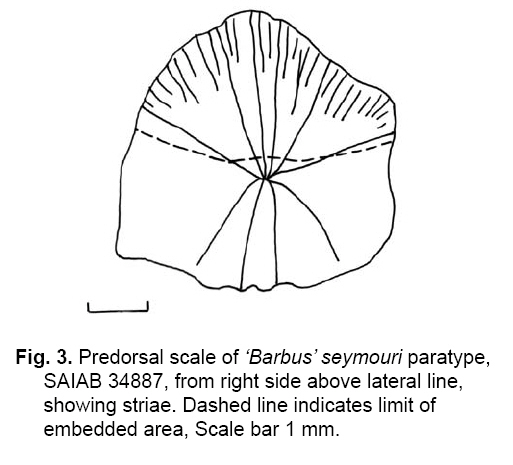
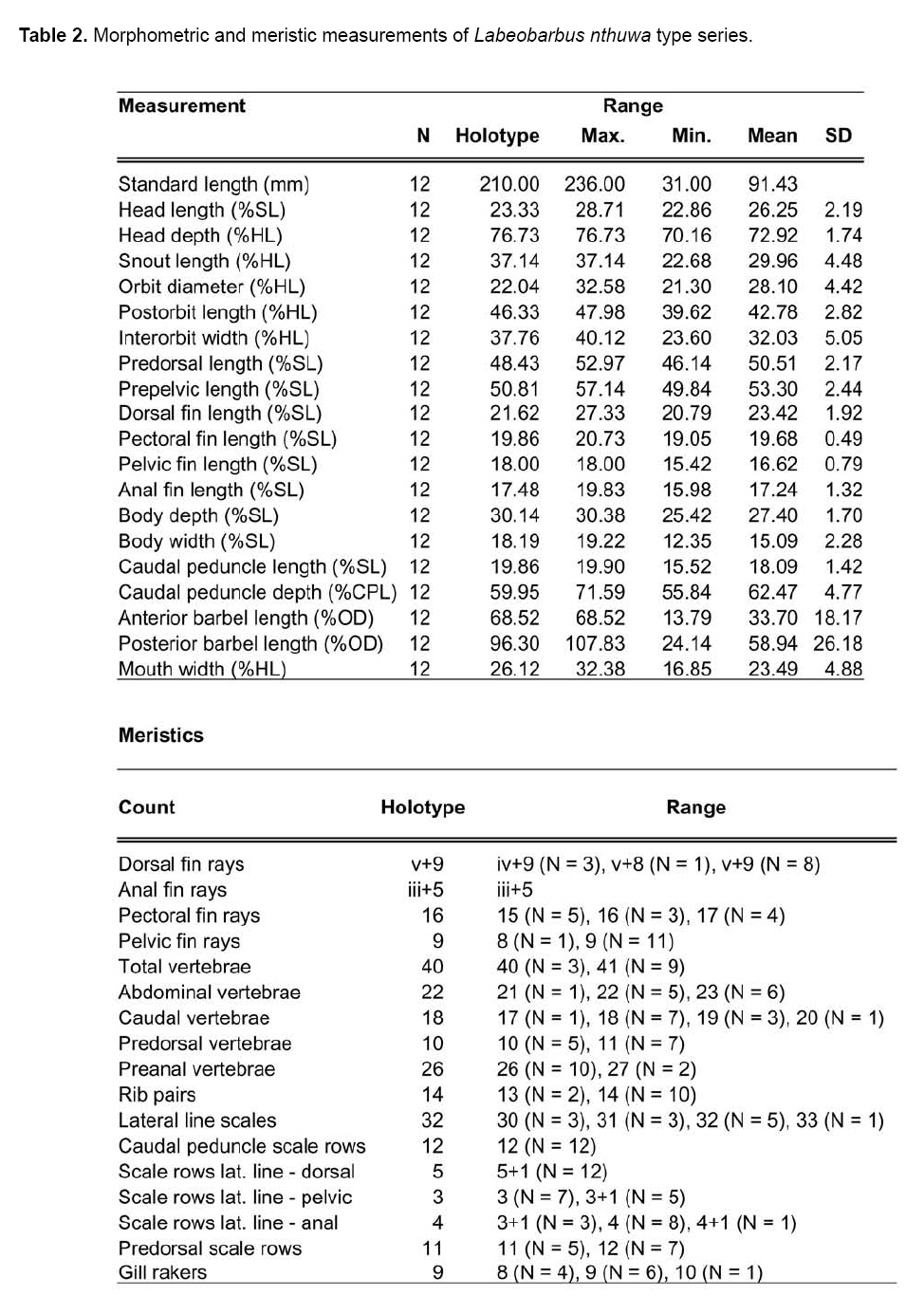
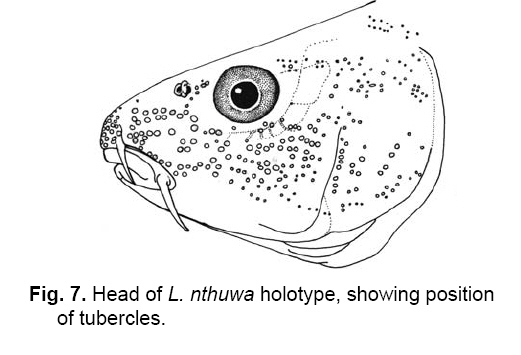
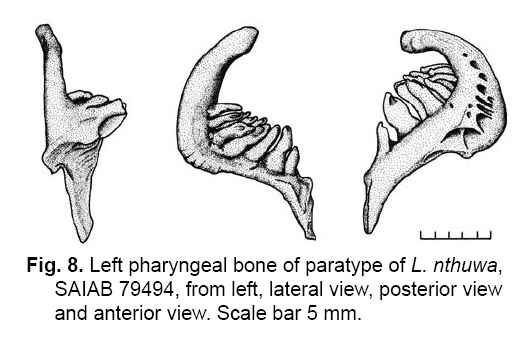
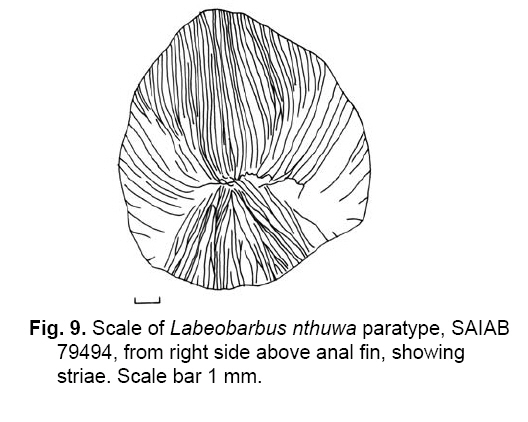
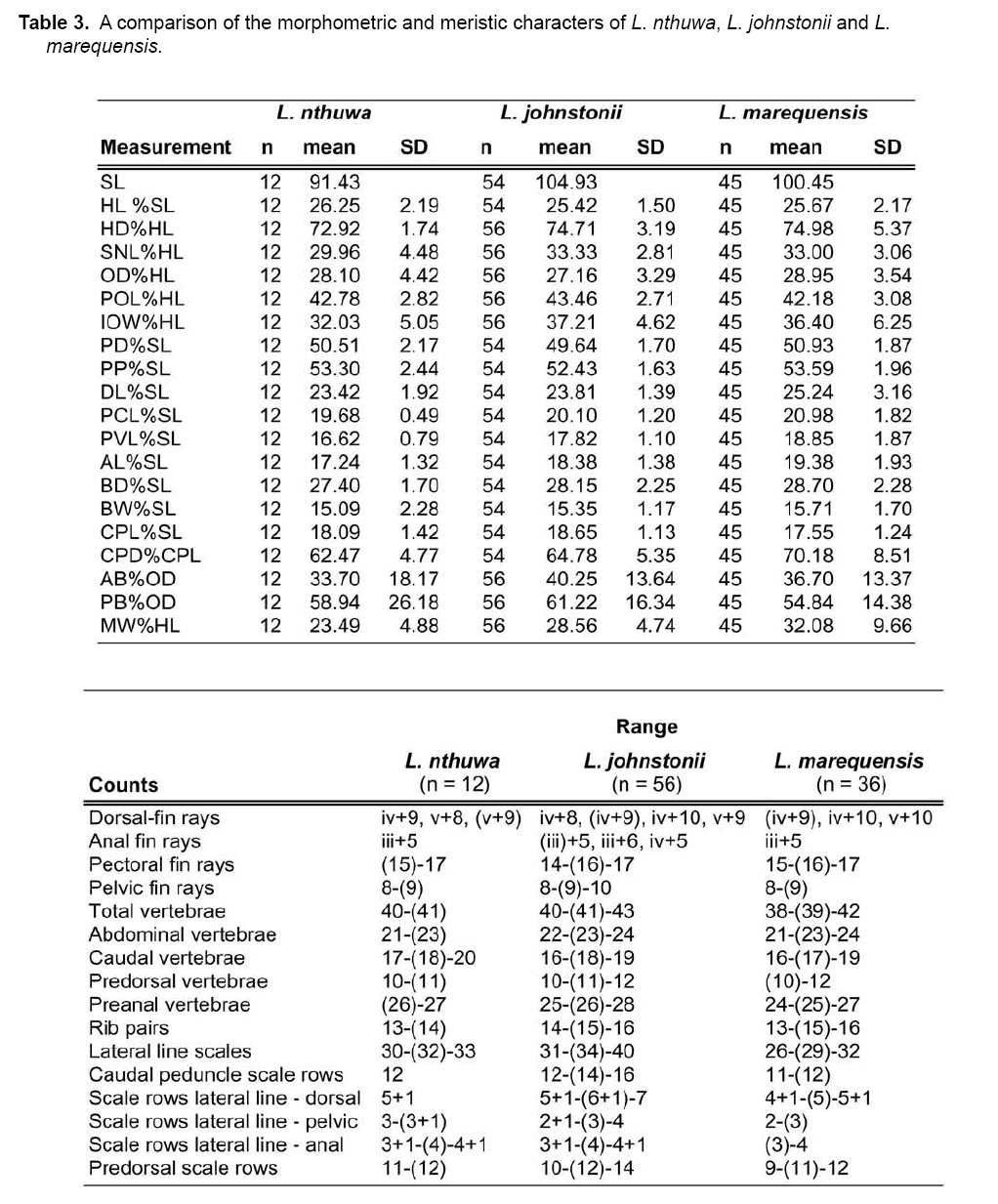
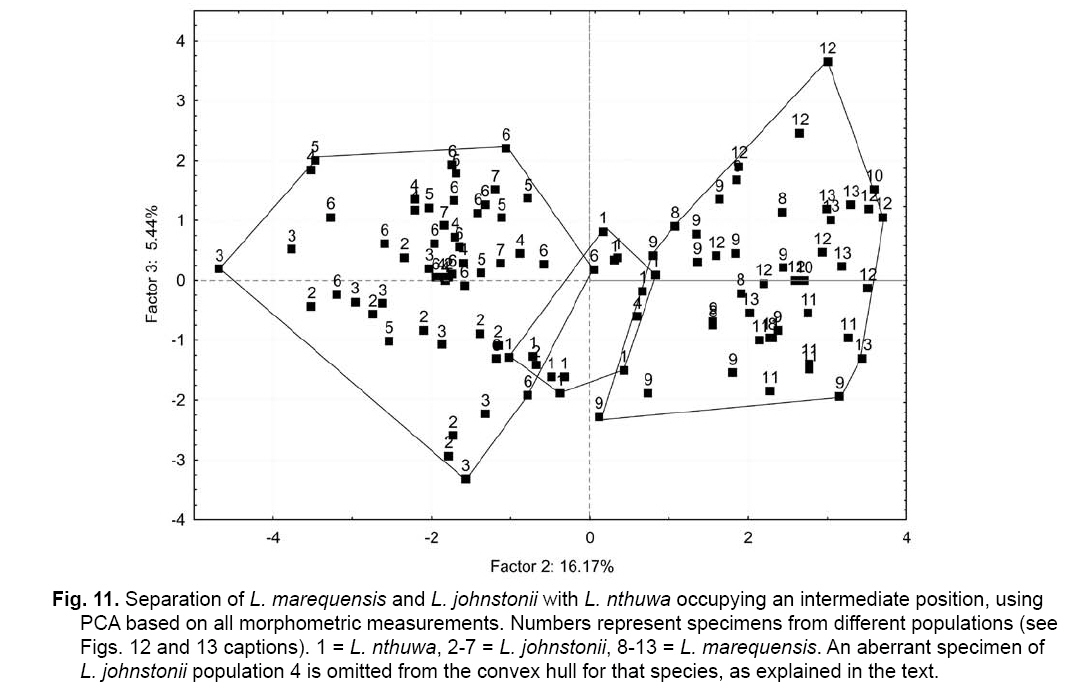
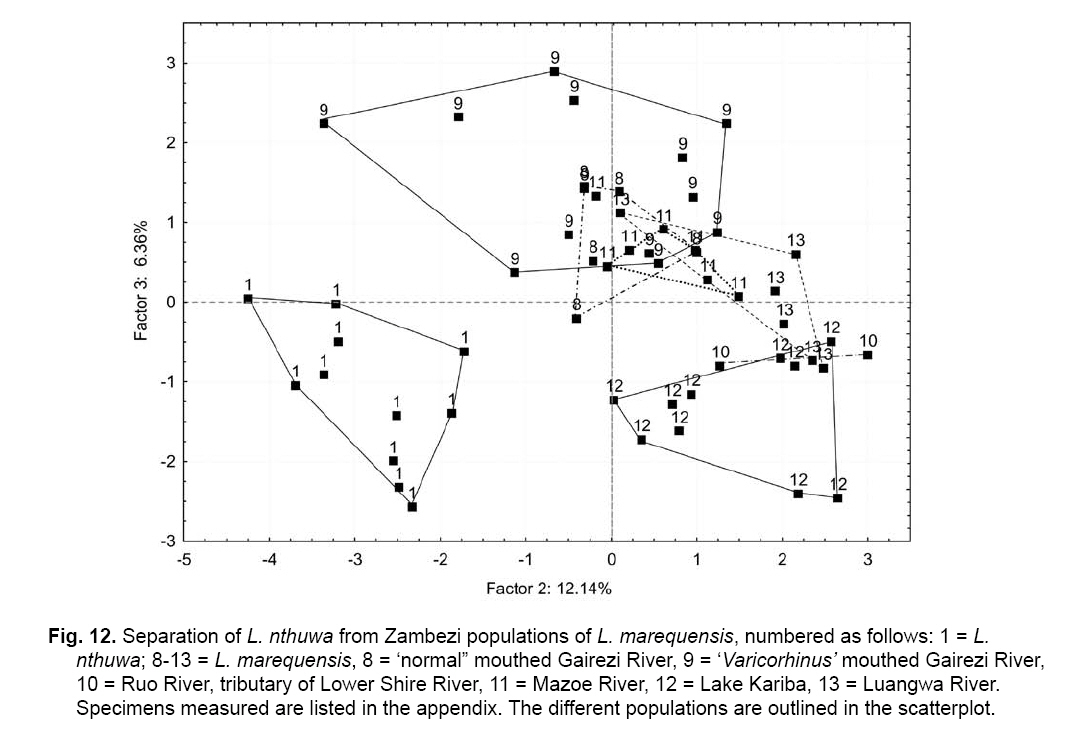
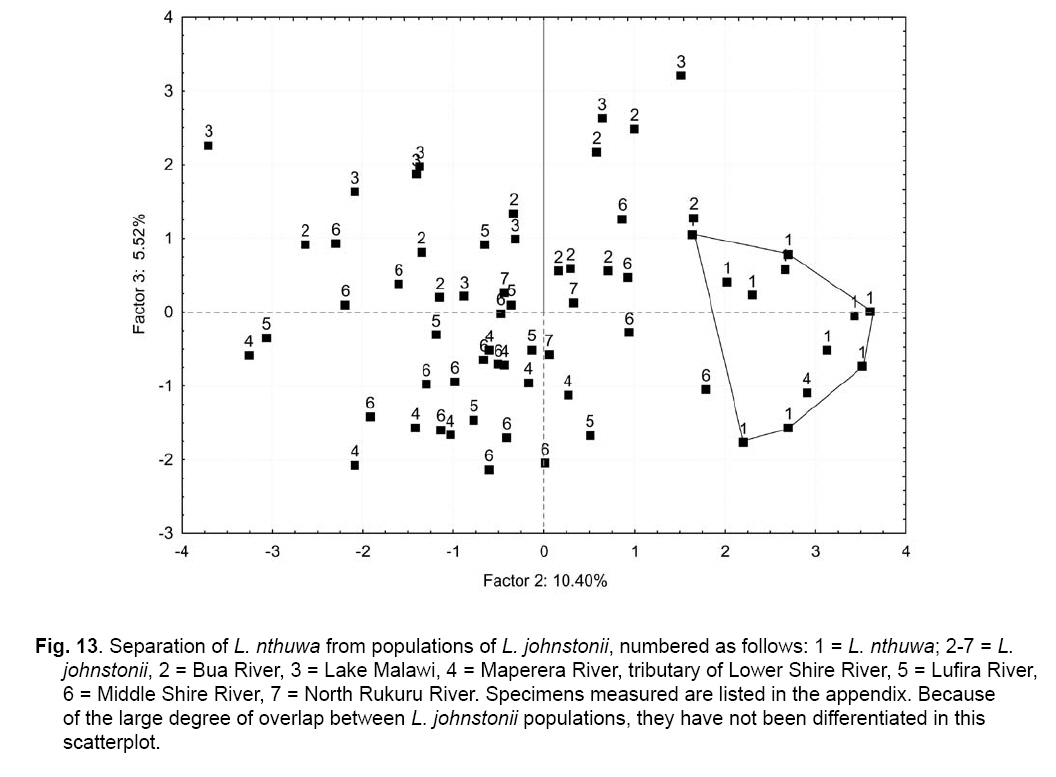
Smithiana, Publications in Aquatic Biodiversity, Bulletin 8, February, 2008, pp. 25-39 New species of 'Barbus' and Labeobarbus (Teleostei: Cyprinidae) from the South Rukuru River, Malawi, Africa. Denis Tweddle and Paul H. Skelton South African Institute for Aquatic Biodiversity, Private Bag 1015, Grahamstown, South Africa. Received 23 February 2007; accepted 14 November 2007. Code Number: sm08002 ABSTRACT Two new barbine species are described from the South Rukuru River, which flows into Lake Malawi. 'Barbus' seymouri sp. nov. differs from the 'B' eutaenia / 'B'. miolepis species complex in the absence of the sheath of enlarged scales at the base of the dorsal fin, which is prominent in the other species of the complex, and in the pigmentation pattern of the mid-lateral stripe. Labeobarbus nthuwa, sp. nov. differs from L. marequensis of the Middle and Lower Zambezi River system and from L. johnstonii of the Lake Malawi system in the presence of a bony dorsal spine, and from L. codringtonii of the Upper Zambezi in the absence of the tall dorsal fin. It also differs from L. johnstonii in scale counts, and from L. marequensis in gill raker counts and caudal peduncle proportions. The fish fauna of the South Rukuru River is separated from that of Lake Malawi and all other inflowing rivers by steep rift escarpment waterfalls. Evidence is presented for river capture from the Luangwa River, which flows to the Middle Zambezi. Other Zambezian species found on the northern Malawi lakeshore are considered to be evidence for a separate, former link to the Zambezi system via the Chambeshi system. Key Words: new species, Lake Malawi, Luangwa River, river capture INTRODUCTION The South Rukuru River rises in the central uplands of Malawi and flows to Lake Malawi (Fig. 1). In its upper reaches it flows north, close to the Zambian border, before turning east and passing between the high Viphya and Nyika plateaux. For much of its northward passage, the terrain is fairly flat and the river slow flowing, but it is fed by fast-flowing mountain streams that arise on the two plateaux, many of which are perennial. After cutting through the Precambrian granitic basement complex rocks of the Njakwa gorge, the river continues to the lake down the rift escarpment as a series of rapids cut through a heavily faulted Karooage trough. Shortly before entering the lake, the river drops over the Wongwe and FuFu waterfalls, which form a barrier preventing the upstream movement of lake fishes (Tweddle 1982). The fish fauna of the South Rukuru River, its tributaries and neighbouring rivers has been sampled on several occasions and differs considerably from that of all other affluent streams of Lake Malawi (Tweddle 1982; 1996; Tweddle & Willoughby 1978). The species collected in the South Rukuru have been compared with known species from other river systems in Africa, and three of the barbine species are new to science. Two species are described here, while the third will be described later in a review of small spotted 'Barbus' species in the region (Tweddle, in prep.). The use of 'Barbus' follows Berrebi et al. (1996), under which the generic name Barbus is restricted to Barbus barbus (L.) and its large European congeners. The small African barbine species with radiate scale striations are not closely related and therefore do not belong in the same genus. Until further research clarifies their relationships, the temporary epithet 'Barbus' will be used. One of the species described herein is a member of the group of smaller 'Barbus' species with radially striated scales, a serrated dorsal spine and a mid-lateral stripe. This species also occurs in the Kaziwiziwi River (Fig. 1), a tributary of the North Rumphi River that arises close to tributaries of the South Rukuru on the southern flanks of the Nyika Plateau. The Kaziwiziwi River joins the North Rumphi before it falls steeply over the rift escarpment to Lake Malawi. Below the escarpment, sampling in the North Rumphi yielded only the typical lakeshore stream fauna (unpublished catch data; the lakeshore stream fauna is described by Tweddle (1996)). The second species to be described is a member of the large African barbine group with parallel-striated scales recognised as the recently reinstated genus Labeobarbus Rüppell 1835 (Skelton 2001). In Lake Malawi and its other inflowing rivers, this genus includes two species, L. brevicauda (Keilhack) and L. johnstonii (Boulenger). METHODS Specimens were collected from various sites (Fig. 1) in the South Rukuru River and tributaries and in the Kaziwiziwi River by electric fishing and the three largest specimens of Labeobarbus nthuwa (sp. nov.), were purchased from a local angler. Specimens were preserved on site in 10% formalin, transferred to 60% N-propyl alcohol, and were later transferred to 70% ethyl alcohol. Measurements were made by vernier calipers to the nearest 0.1 mm following Skelton (1988). Skeletal meristics and fin ray counts were taken from radiographs. Vertebral counts include the four Weberian vertebrae and a single (PU + U1) caudal fin vertebra. Abdominal vertebrae include all anterior vertebrae without an extended haemal spine. Caudal vertebrae possess an extended haemal spine. Lateral line scales were counted from the first scale row behind the gill cover to the end of the body, i.e. the point of flexure of the caudal fin. This usually excludes the last pored scale and smaller scales on the fin base. In transverse scale counts where a count is given as e.g. 2+1, this means two full size scales and either one small scale or a gap between scale and fin with the next scale overlapping the fin base. Colour descriptions were made from field observations and colour transparencies of live fishes. Pharyngeal bones were dissected and macerated in trypsin for several days before being defleshed. Scales were removed and stained with alizarin red. Scales and pharyngeal teeth were drawn using a camera lucida. Institutional abbreviations follow Leviton et al. (1985). The following abbreviations are used in text and tables: SL, standard length; HL, head length; HD, head depth; SNL, snout length; OD, orbit diameter; POL, postorbital length; IOW, interorbital width; PDL, predorsal length; PPL, prepelvic length; DL, dorsal fin length; PCL, pectoral fin length; PVL, pelvic fin length; AL, anal fin length; BD, body depth; BW, body width; CP, caudal peduncle; CPL, caudal peduncle length; CPD, caudal peduncle depth; AB, anterior barbel length; PB, posterior barbel length; LJW, lower jaw width; LL, lateral line. STATISTICA 8 software was use for the principal component analysis. 'Barbus' seymouri sp. nov. Holotype. SAIAB 34888, unsexed, 93.3 mm SL; Kaziwiziwi River, North Rumphi River basin, Lake Malawi affluent, Malawi, 10º38'S, 34º05'E; D. Tweddle, 25 September 1980. Paratypes. SAIAB 34887, 17 specimens, 52.4-108.2 mm SL, collected with holotype. SAIAB 34946, 1 specimen, 74.0 mm SL, collected with holotype. SAIAB 34881, 5 specimens, 28.3-92.4 mm SL, South Rukuru River at Njakwa gorge, 11º02'S, 33º54'E, and Runyina tributary, 11º01'S, 33º47'E, at Rumphi, Malawi, D. Tweddle, 22-23 October 1980. Other material examined. SAIAB 40797, 20 specimens, 31.6-51.2 mm SL, Mzimba River (South Rukuru tributary), Mzimba, Malawi, 11º54'S 33º36'E, D. Tweddle & P.H. Skelton, 2 July 1992; BMNH 1978.8.3.537-556, 20 specimens, South Rukuru River, 12º16'S, 33º29'E, and Mzimba tributary, 11º54'S, 33º36'E, D. Tweddle & N.G. Willoughby, October 1976; BMNH 1978.8.3.1596, 1 specimen, South Rukuru River 12º16'S, 33º29'E, D. Tweddle & N.G. Willoughby, October 1976; further specimens are also deposited in the museum in the Monkey Bay Fisheries Research Unit Museum, Malawi, without accession numbers. DIAGNOSIS. A moderate-sized (up to 108 mm SL), robust bodied, 'Barbus' species with an ossified, serrated last dorsal simple ray, 26-29 lateral line scales, fins tinted pinkish-orange, and a midlateral black stripe that extends through the caudal fin to the fork, but does not extend onto the snout. Differs from other orange-finned, serrated-spined 'Barbus' species in the region (including 'B.' eutaenia Boulenger, 1904, 'B.' miolepis Boulenger, 1902, 'B.' choloensis Norman, 1925, and several other species, some of which are undescribed, recently discovered in Upper Zambezi tributaries (Tweddle et al., 2004)) in the absence of the prominent sheath of enlarged scales at the base of the dorsal fin present in the other species, and in the pattern of pigmentation of the mid-lateral stripe, which is distinct in each species. DESCRIPTION. Based on the holotype and paratypes. Morphometric and meristic data are given in Table 1. Body fusiform, slightly compressed, maximum depth approximately equal to head length, located before dorsal fin. Nape rises in hump behind head, particularly in larger specimens. Head moderately deep, four times in SL. Eyes lateral in position, visible from above and below, orbit becoming proportionately smaller in large specimens (OD as %HL = -0.5952 HL + 39.184, r² = 0.7346). Snout rounded and short, equal to or slightly shorter than orbit diameter. No tubercles on the head. Nostrils small, short tubular anterior naris adjacent to open posterior naris; nostrils level with dorsal margin of eye and separated from orbit by less than one orbit radius. Mouth subterminal, crescent-shaped and reaching to below anterior border of orbit, lips moderately well-developed. Two pairs of simple barbels, anterior slightly shorter than posterior, which is approximately equal to orbit diameter. Gill cover opening from level with dorsal margin of orbit, attached ventrally close to isthmus. Gill arches with four short, stub-like, widely-spaced gill rakers on ceratobranchial of anterior arch, two on epibranchial. Pharyngeal bones typical of small African 'Barbus' species with three rows of peg-like pharyngeal teeth with pointed cusps, formula 5,3,2 2,3,5. Origin of dorsal fin equidistant between tip of snout and base of caudal fin, above or just behind origin of pelvic fins. Dorsal fin becomes relatively shorter with increasing size of specimen, from approximately 30% of SL in smallest specimens to 20% in largest individuals, anteriormost branched ray longest, distal margin concave. Last unbranched dorsal ray ossified and finely serrated (approximately 30 prominent serrations and up to 40 in total) on its posterior side. Pectoral fins reach ⅔ of distance to base of pelvics, tips rounded and distal margin straight. Pelvic fins reach ⅔ of distance to base of anal fin, relatively small and rounded. Anal fin short, extending ⅓ length of caudal peduncle, last unbranched ray longest, distal margin straight. Caudal fin forked, outer rays twice length of median rays, lobes rounded. Caudal peduncle long, length twice depth. Anus and genital opening immediately anterior of base of anal fin. Gut short, about equal to SL, in a single simple S-flexure. Scales moderately large, cycloid and rounded, well developed in regular rows. Scales radially striate with about 10 radii in total (Fig. 3). Lateral line complete, anteriorly dipping one scale row below the horizontal myoseptum, joining and extending straight along the midline at the anterior end of the caudal peduncle. No sheath of enlarged, elongate scales along base of dorsal fin; short pelvic axil scale present; breast scales well developed,. Colouration. Live colours (Fig. 4): body and head olive dorsally and sides silvery with whitish-gold sheen ventrally; black midlateral stripe bordered with golden yellow; black post-opercular vertical bar. Dorsal fin with dark olive rays and clear membranes; caudal fin colouration similar but with pinkish-orange tinge, particularly on lower half; pectoral, pelvic and anal fins pale brownish-orange tinted distinctly pinkish-orange. Iris dark anteriorly and posteriorly, pale yellow ventrally with orange highlight dorsally. Specimens from turbid water uniformly pale with faint dark midlateral stripe. In preserved specimens (60% propyl alcohol), dorsal surface scales densely pigmented with fine melanophores, with a crescent of closely spaced larger melanophores at both anterior and posterior borders of exposed part of scale. Pigmentation less intense in centres of lateral line scales and in the row below the lateral line only a few scattered melanophores present on each scale, although anterior crescent is still fairly prominent. In next row below, only a few scattered melanophores present anteriorly and dorsally on each scale. Below this, all scales pigment-free except for a few melanophores along the ventral scale row between anal and caudal fins. Crescent of dark pigment present behind operculum from lateral line to pectoral fin base. Horizontal myoseptum darkly pigmented beneath the scales, forming a fine but fairly intense mid-lateral stripe, most prominent on the caudal peduncle. In many specimens, underlying pigmentation emphasised by additional melanophores along the line on the scales themselves. At base of caudal fin, lateral stripe broadens slightly as a result of heavier scale pigmentation. Stripe continues through to posterior margin of caudal fin. Mid-dorsal stripe present for full length of body. Rays of the dorsal, pectoral and caudal fins lightly pigmented with fine melanophores, pelvics and anal almost pigment free. Operculum and snout covered in melanophores, which are more abundant but finer on the snout. Both pairs of barbels lightly pigmented. DISTRIBUTION AND HABITAT. 'Barbus' seymouri is found throughout the South Rukuru system above the falls that isolate the fauna from that of Lake Malawi, and in the Kaziwiziwi River above the escarpment down to the lakeshore. The sites where the species is caught are generally clear, strongly-flowing streams with cover in the form of vegetation and/or rocks. In the Mzimba River in Mzimba town, the river changed dramatically between sampling in 1976 and 1992. The narrow, well-vegetated, clear stream of 1976 yielded many large well-coloured specimens of 'B'. seymouri, whereas in 1992 the wide, shallow, sandy stream at the same site yielded small, pallid specimens only from the road bridge gabions. ETYMOLOGY. Named after the late Tony Seymour (Fig. 5), a close friend of the first author and colleague for many years in the Malawi Government Fisheries Department, in recognition of his many years of service to Malawi not only in Fisheries but also in many other aspects of environmental management and conservation, and in particular for his long-term commitment to supporting Lake Malawi's fishermen. COMPARISONS. 'Barbus' seymouri is one of a group of small African 'Barbus' with radiate striated scales, a bony, serrated last unbranched dorsal-fin ray, orange tinted fins and a dark midlateral stripe. The three most similar species so far recorded in eastern, central and southern Africa are 'B.' choloensis, 'B.' eutaenia and 'B.' miolepis. 'Barbus' choloensis occurs in the south of Malawi in two Lower Shire River (i.e. Lower Zambezi system) tributaries, the Ruo River and the Mwabvi River (Tweddle & Willoughby 1979). 'Barbus' eutaenia is currently recognised as being widespread in southern-central Africa while 'B.' miolepis is considered to be a species of the Upper Zambezi, Kafue and Congo river systems (Skelton 2001). All three are robust species with yellow or orange tinted fins, a black mid-lateral stripe and a serrated dorsal spine. The 'B.' eutaenia / 'B.' miolepis species complex needs detailed revision and several other species new to science exist, including another possibly distinct species in the lower reaches of the Lufirio River, which flows into northern Lake Malawi from Tanzania (SAIAB 54766). In the Upper Zambezi system several other species in this complex occur, some of which are undescribed (Tweddle et al. (2004). Taxonomic review of these species will not affect the status of 'B.' seymouri. 'Barbus' seymouri has larger scales than 'B.' choloensis with 4 scales between the dorsal fin and lateral line compared to 5-6 scales in 'B.' choloensis. 'Barbus' seymouri has 12 scales around the caudal peduncle and 'B.' choloensis has 14-16. 'Barbus' seymouri has a more rounded snout, thus the mouth is slightly sub-terminal, whereas the snout of 'B.' choloensis is pointed and the mouth terminal. In similar-sized specimens, 'B.' choloensis has a much larger eye. 'Barbus' choloensis has pointed caudal fin lobes whereas those of 'B.' seymouri are rounded. In 'B.' choloensis, the mid-lateral stripe is diffuse, confined to the posterior half of the body, broadens to an indistinct oval spot one full scale wide on the caudal peduncle, and does not extend noticeably on to the caudal fin rays. The mid-lateral stripe of 'B.' seymouri is narrower and better defined, extends the full length of the body following the curve of the scale row above the lateral line, broadens only slightly on the caudal peduncle and continues through the caudal fin to its fork. 'Barbus' seymouri differs from the other species in the 'B.' eutaenia / 'B'. miolepis complex in that it lacks the enlarged scales that form a sheath at the base of the dorsal fin in the other species. It also has more lateral line scales (26-29 v. 23-27). The mid-lateral stripe of 'B.' seymouri is finer than the broad heavy stripe of the other species. The well-defined stripe in the other species extends through the operculum and to the tip of the snout, whereas in 'B.' seymouri there is only a suggestion of heavier pigmentation in the centre of the operculum and only diffuse melanophores on the snout. Labeobarbus nthuwa sp. nov. Holotype. SAIAB 39341, 210 mm SL; Runyina tributary of South Rukuru River, Lake Malawi affluent, Malawi, 11º01'S 33º47'E; D. Tweddle & P.H. Skelton, 3 July 1992. Paratypes. SAIAB 79494, 2 specimens, 217-236 mm SL, collected with holotype. SAIAB 40787, 6 specimens 31.0-36.7 mm SL, Runyina, 11º01'S 33º47'E, and South Rumphi, 11º01'S 33º52'E, tributaries of South Rukuru River; D. Tweddle & P.H. Skelton. SAIAB 39293, 1 specimen, 42.1 mm SL, Runyina tributary of South Rukuru River, 11º01'S 33º47'E; D. Tweddle & P.H. Skelton. SAIAB 51928, 2 specimens, 210-217 mm SL, South Rukuru River at Chikulamayembe, Malawi, 10º45'S 34º07'E; D. Tweddle, R. Bills & P.H. Skelton, 22 October 1995. DIAGNOSIS. A Labeobarbus species with parallel-striated scales, a heavily ossified, unserrated, last unbranched dorsal-fin ray, five unbranched dorsal rays in total, and 30-33 lateral line scales. Differs from the two Lake Malawi Labeobarbus species (L. johnstonii Boulenger 1907 and L. brevicauda Keilhack 1908) in scale counts and the presence of the bony dorsal spine. Differs from L. marequensis (A. Smith 1841) of the Zambezi River and east coastal rivers south to the Phongolo in gill raker counts, caudal peduncle proportions and the presence of the bony dorsal spine. Differs from L. codringtonii (Boulenger 1908) of the Upper Zambezi River in the absence of the high dorsal fin characteristic of that species. DESCRIPTION. Based on the holotype and paratypes. Morphometric and meristic data are given in Table 2. Body fusiform, slightly compressed, body depth approximately equal to head length in smaller specimens, becoming proportionately deeper with size, to approximately 1½ times head length in adults, deepest at origin of dorsal fin. Head length 4 times in standard length, head depth approximately ¾ head length. Dorsal profile of head in larger specimens straight to slightly concave (Fig. 7), shallowly convex in smaller specimens. Eyes lateral in position, visible from above and below, becomingproportionatelysmallerinlargespecimens(OD as %HL = 54.662 HL-0.236, r² = 0.971). Snout pronounced, particularly in larger specimens, longer than orbit diameter in fish over 40 mm SL, shorter than orbit diameter in smaller specimens. Small conical tubercles present on snout, operculum and sides of head, not extending on to dorsal surface of head or on to body. Nostrils prominent, short tubular anterior naris adjacent to open posterior naris and separated by raised rounded septum. Nostrils at horizontal through dorsal half of eye, separated from orbit by less than one orbit radius. Mouth sub-terminal, not reaching below anterior border of orbit; lips variable, ranging from specimens with straight, keratinised scraping edge to the lower lip (Varicorhinus-like) to specimens with rounded, fleshy lips. Two pairs of barbels, anterior just over half length of posterior, which ranges from being equal to orbit diameter in largest specimens examined to less than one-half orbit diameter in smallest specimens. Gill cover opening from level with dorsal margin of orbit, attached ventrally close to isthmus. Gill arches with 8-10 robust gill rakers visible on anterior arch. Pharyngeal bones robust with pharyngeal teeth in three rows, formula 5,3,2 2,3,5 (Fig. 8). Origin of dorsal fin equidistant between tip of snout and base of caudal fin, positioned before origin of pelvic fins, last simple ray heavily ossified and unserrated, anterior branched ray longest, distal margin of fin strongly concave. Radiographs show five simple rays in dorsal fin in all but smallest three specimens, where first, very small ray may have not yet developed to a size where it becomes visible. Pectoral fins falcate, reaching ¾ of distance to base of pelvic fins. Pelvic fins pointed with square distal margin, reaching ⅔ of distance to base of anal fin. Anal fin last unbranched ray longest, distal margin shallowly concave, reaching to ½ length of caudal peduncle. Single rows of prominent tubercles present on posterior edge of branched rays of anal fin in adult males. Caudal fin deeply forked, outer rays nearly four times length of median rays, lobes pointed. Caudal peduncle of moderate length, depth generally more than 60% of length. Anus and genital opening located at anterior of base of anal fin. Gut extended and involuted, its length several times longer than SL. Scales moderately large, cycloid and rounded, well developed in regular rows. Scales with numerous parallel striations on exposed field, less numerous on medial area of embedded field, focus ill-defined (Fig. 9). Lateral line complete, anteriorly dipping one scale row below midline, which it rejoins anterior to anal fin, passing mid-laterally to base of caudal fin. Elongated pelvic axil scale present; breast scales well developed but markedly reduced in size between bases of pectoral fins. Colouration. Live colour (Fig. 10) silvery-olive dorsally, silver laterally and ventrally with faint purple sheen that is most prominent on operculum. Dorsal and caudal fins dark olive-grey; anal, pelvic and pectoral fins paler with brownish-orange bases, greyish extremities. Preserved specimens (60% propyl alcohol) with scales on dorsal surface lightly pigmented with fine melanophores, more closely spaced at anterior border of exposed part of scale. Pigmentation less intense ventral to lateral line scales with melanophores confined to anterior of exposed part of scale. Ventral scales unpigmented. Vertical crescent of dark pigment behind operculum from lateral line to pectoral fin base. Fin membranes between rays of dorsal, caudal and pectoral fins heavily pigmented, outlining the rays. Similar pigment appears on pelvic and anal fins, but less prominently. Caudal fin with fine, dark posterior edge as result of membrane pigmentation. Dorsal surface of the head very finely pigmented, with larger melanophores in the centre of operculum. Posterior edge of operculum unpigmented. Ventral surface of head, lips and barbels are unpigmented. DISTRIBUTION AND HABITAT. Adult L. nthuwa occur in the main South Rukuru River down to the Wongwe and FuFu falls near the lake (Fig. 1), but have not been found in the lake itself. The upper distribution limit is unknown, but the species occurs at least as far upstream as Lake Kazuni. The species also occurs in larger perennial tributaries flowing from the Nyika Plateau, such as the Runyina, where adults and juveniles were caught, and the South Rumphi, a smaller stream that yielded juveniles only. All sampling sites where the species has been caught were rocky, fast flowing stretches with some deeper pools. The species has not been recorded beyond the South Rukuru River system. ETYMOLOGY. Nthuwa (pronounced 'ntoowa') is the vernacular name used for this species in the vicinity of the town of Rumphi, and is used here as a noun in apposition. COMPARISONS. There are two similar large Labeobarbus species in Lake Malawi and all perennial rivers and streams flowing to the lake with the exception of the South Rukuru above the Wongwe and FuFu Falls. Labeobarbus brevicauda (Keilhack), formerly known as L. eurystomus (Keilhack) (Seegers 1995), is easily distinguished from L. nthuwa by its higher lateral line scale count (33-37, mean 35 v. 30-33, mean 31), long, robust barbels of equal length and enlarged, molariform pharyngeal teeth. Labeobarbus johnstonii is closer in appearance to L. nthuwa and has similar pharyngeal bones and teeth. Because L. johnstonii is variable in form depending on habitat and locality, e.g. Shire River specimens from torrential stretches of river are deeper bodied than specimens from standing or slow moving waters in Lake Malawi and inflowing streams, it was necessary to investigate the full range of variation in the species. A total of 56 specimens from Lake Malawi, various inflowing rivers at different altitudes in different habitat types, the out-flowing Shire River, and small tributaries of that river in the Lower Zambezi system were measured. Labeobarbus johnstonii can be separated from L. nthuwa by its higher lateral line scale count (31- 40, mean 35), by its higher gillraker count (L. johnstonii, 10-15, mean 11; L. nthuwa, 8-10, mean 9) (Table 3), and by having a flexible and not heavily ossified last simple dorsal ray, as in L. nthuwa. Labeobarbus nthuwa has five unbranched dorsal-fin rays visible on radiographs of specimens >38 mm SL, whereas in L. johnstonii, the usual count is four: only four of 58 specimens radiographed have five unbranched rays. Labeobarbus nthuwa was also compared with L. marequensis from the Zambezi system. Labeobarbus marequensis, as currently recognised, is found from the Middle and Lower Zambezi south to the Incomati system in South Africa, including the type locality, the Marico River (Skelton 2001). L. marequensis populations show considerable variation, e.g, populations to the south of the Zambezi have higher scale counts and narrower caudal peduncles (discussed by Farquharson 1962). Following preliminary examination of the other populations, our study was restricted to Zambezi populations as these are closest to Malawi drainages, most similar in scale counts, and most likely to be related to L. nthuwa. Meristic and morphometric measurements were taken for 46 specimens of L. marequensis from the Middle and Lower Zambezi River and tributaries (Table 4). From the Gairezi tributary of the Lower Zambezi, both typical L. marequensis and chisel-mouthed forms previously identified as Varicorhinus nasutus Gilchrist and Thompson 1911 (see Tweddle & Skelton 1998, for discussion of validity of V. nasutus) were measured. The various Zambezi populations differ in the depth of the caudal peduncle. Specimens from the Lower Shire tributary of the Lower Zambezi have very broad caudal peduncles, as deep as long, whereas specimens from the Gairezi tributary have narrow caudal peduncles. A list of the measured specimens of L. johnstonii and L. marequensis is presented in the Appendix. Labeobarbus marequensis can be separated from L. nthuwa by its higher gillraker count (9-14, mean 11 cf. 8-10, mean 9) and the absence of the ossified last simple dorsal-fin ray. Specimens of L. marequensis from Lake Kariba have slightly more robust last unbranched dorsal-fin rays than other populations, but not to the extent that they could be considered spinous. Only two of 36 radiographed specimens of L. marequensis had five unbranched dorsal-fin rays, the rest had four. Because of the wide geographic variation in L. johnstonii and L. marequensis, caution is needed when describing a new species. To assess the validity of L. nthuwa, principal component analysis was carried out on the measurements from all specimens examined. The first principal component summarised body size, as indicated by its large weights for linear measurements and low weights for meristic variables, and did not help to differentiate the species. Components 2 and 3, however, separated L. marequensis and L. johnstonii, based largely on scale counts (factor 2), barbel lengths and caudal peduncle depth (factor 3) (Fig. 11), whereas L. nthuwa was intermediate, with slight overlap with both species. A single aberrant specimen from the Maperera tributary of the Lower Shire River had 32 lateral line scales and only 12 around the caudal peduncle, and was possibly a L. johnstonii/L. marequensis hybrid as both species occur in the Lower Shire. This specimen is excluded from the convex hull drawn to encompass the L. johnstonii data points in Fig. 11. Labeobarbus nthuwa was compared with the different populations of L. marequensis (Fig. 12) and L. johnstonii (Fig. 13) in separate analyses. Labeobarbus nthuwa and L. marequensis were clearly separated, with scale counts and caudal peduncle depth (factor 2) and gillraker counts and barbel lengths (factor 3) identified as key variables. Some clustering of different Zambezi populations of L. marequensis is also noted on this figure, and further investigation using genetic and morphometric techniques on fresh L. marequensis material is needed to ascertain the relationships between populations of this species. In the third analysis, L. nthuwa was also distinguished from L. johnstonii (Fig. 13), based on scale and gillraker counts (factor 2), caudal peduncle depth and barbel length (factor 3). There is considerable overlap between L. johnstonii populations and thus they are not outlined in Fig. 13. Middle Shire River specimens, from torrential stretches of river, have more powerful caudal regions than specimens from standing or slow moving waters in Lake Malawi and inflowing streams, and this was reflected in the weight given to caudal peduncle depth in the third principal component. As a result, Lake Malawi specimens (population no. 3 in Fig. 13) are all found in the upper part of the scatter diagram, while the robust Middle Shire specimens (population no. 6) are mainly in the lower part. Labeobarbus codringtonii is a species found in rapids of the Upper Zambezi River system and is distinguished from L. nthuwa by the absence of the ossified last simple dorsal-fin ray and from both L. marequensis and L. nthuwa in having a very tall dorsal fin. DISCUSSION Banister & Clarke (1980) considered L. johnstonii to be 'strikingly similar' to L. marequensis in gross appearance, including the form of the pharyngeal bones and teeth and the scale striations. They cautioned that the characters in question are probably plesiomorphic, but stated that, with no other likely candidates in the region, there are no zoogeographical inconsistencies in the suggestion that the two species are related. Labeobarbus nthuwa is an addition to this group of similar species. In scale counts L. nthuwa is closer to L. marequensis than it is to L. johnstonii. Labeobarbus marequensis occurs in the Luangwa River. The very close proximity of the South Rukuru River to the Luangwa River watershed on the Zambian border for approximately 150 km of its course, and the presence of swamps and evidence of former lakes in the area (Hopkins 1973) suggests river capture has occurred as a result of tectonic warping of the land surface associated with the Lake Malawi rifting. Unconsolidated pebble sheets up to 30 m above the present level of Lake Kazuni (Fig. 1) probably represent the littoral deposits of a once much more extensive lake (Hopkins 1973) created by such tectonic movements. A probable site of river capture is marked on Fig. 1. Prior to this, much of the present South Rukuru River would have been a west-flowing Luangwa River tributary. The Luangwa River in Zambia flows to the Middle Zambezi. It is possible that L. nthuwa is derived from a population of L. marequensis present in the Luangwa tributary captured by the South Rukuru River. Unfortunately no Labeobarbus specimens are available in museum collections of the Luangwa River's eastern tributaries near Malawi to compare with L. nthuwa. An alternative possibility is that L. nthuwa evolved from a population of L. johnstonii present in the South Rukuru before the Wongwe and FuFu Falls became a complete barrier to migration during rifting. Another possible interpretation is that L. johnstonii is derived from Luangwa River L. marequensis, which entered the Lake Malawi system via the South Rukuru River. With the South Rukuru separated from both systems as a result of the river capture to the west and falls to the east, L nthuwa would then have evolved its distinctive characteristics in isolation, while the lake populations in continuous contact with each other evolved into the species now recognized as L. johnstonii. The alternative suggestion that the ancestor of L. johnstonii entered Lake Malawi from the Lower Zambezi via the Shire River (Banister & Clarke, 1980) is untenable because the Shire link is very recent, established only in the Quaternary (Lister 1967). The South Rukuru River is unique among rivers flowing to Lake Malawi as it contains several fish species absent from the other rivers. In addition to the two species described here, there is another small 'Barbus' species with midlateral spots, which is possibly distinct from a similar species, also undescribed, on the lakeshore plain. Genetic study is needed to resolve this problem. One of the species described in this paper, 'B.' seymouri, also occurs in the adjacent Kaziwiziwi River, from where many of the type series, including the holotype, were taken. The species may have found its way into the Kaziwiziwi naturally as a result of river capture, or possibly by human transfer when a fish farm on the Nchenachena tributary of the South Rukuru, very close to the Kaziwiziwi, was used to stock fish ponds in the area in the 1950s. The South Rukuru River contains two undescribed species of the small catfish genus Zaireichthys, only one of which occurs in other Lake Malawi rivers, while the other is found in the Zambezi system (D.H. Eccles et al., in prep.). Clarias liocephalus Boulenger occurs in Malawi only in the South Rukuru River. This is a species of the Upper Zambezi, north to the Congo, and through the Central African Great Lakes to the Lake Victoria region (Teugels 1986; Skelton 2001). In addition to these distinct species, there are as yet unquantified differences in the appearance of Amphilius uranoscopus (Pfeffer) from the South Rukuru River compared to other Malawi rivers, and the South Rukuru 'Barbus' paludinosus Peters population grows to a much larger size (15 cm SL) than in other populations elsewhere in its wide range (Jackson et al. 1963; personal observations). The distinctness of the South Rukuru fish fauna shows that it is likely to be derived from a different source than the fauna of other rivers flowing into the lake. It is suggested here that the likely source is highland tributaries of the Luangwa River captured by the South Rukuru during tectonic warping of the rift flanks. The fish fauna of the Lake Malawi catchment as a whole shows close links with both east coast rivers and also with the Zambezi system to the west. The distribution of some of these 'Zambezi' species suggests there may have been more than one source. The following 'Upper Zambezi' species occur along the northern Malawi lakeshore plain, including the Limphasa wetlands (Fig. 1), but not in the south: Pollimyrus castelnaui (Boulenger), 'Barbus' bifrenatus Fowler, Clarias stappersii Boulenger, Tilapia sparrmanii Smith and Pseudocrenilabrus philander (Weber). None of these species occur in the South Rukuru River above the falls. Another 'Upper Zambezi' species in the Hippopotamyrus ansorgii (Boulenger) species complex occurs in the upper reaches of the North Rukuru River, to the north of the Nyika Plateau (W. Kadye, pers. comm., specimens lodged in SAIAB). Thus the relationship between Lake Malawi and the Zambezi system is complex and cannot be explained simply by the single river capture from a Middle Zambezi tributary for which evidence is presented in this paper. Worthington(1933)suggestedlinksbetweennorthern affluents of Lake Malawi and the Chambeshi system in Zambia, which is currently a headwater system of the Congo River system in an area that is flat and largely covered by marshes and large lakes. Genner et al. (2007) also suggested links in this area when discussing the molluscan Melanoides polymorpha (Smith) 'complex' of Lake Malawi. They pointed out that the M. polymorpha complex formed a clade with Melanoides anomala (Dautzenberg and Germain), a taxon now restricted to the southeast of the Congo drainage, and Melanoides mweruensis (Smith), an endemic of Lake Mweru. They stated that the Malawi Basin may have been colonised directly via a direct watershed boundary between the headwaters of the Congo River and the northwest of the Malawi Basin. Here tributaries of the Songwe River are adjacent to tributaries of the Chambeshi River that flows directly into the Bangweulu swamps and onto Lake Mweru. Stankiewicz & de Wit (2006) described historical changes in river directions in this area based on geological evidence. They suggested that in the Miocene the Luangwa and Middle Zambezi rivers changed course from their previous northeasterly flow direction towards an eastward-flowing paleo-Congo River, and may have become landlocked until captured by the Lower Zambezi River. The Chambeshi River also originally flowed north east, either joining the Luangwa or flowing directly into the palaeo-Congo. Its flow was also reversed in the Miocene and it became landlocked, possibly originating Lake Bangweulu. This system was captured first by the Kafue River to become part of the Zambezi River network and then, in the Lower Pleistocene, by the Luapula River to become part of the new west-flowing Congo River network, as it is today. The course of the proto-Luangwa River when flowing northeast cuts across the northern tip of the current Lake Malawi. Thus there have been opportunities for transfer of fish species in the area as proposed by Worthington (1933), explaining the presence of the group of species discussed above in northern Lake Malawi lakeshore streams. Further study is needed on the faunas of Zambian waters between the Upper Zambezi and Lake Malawi, including eastern Luangwa tributaries, to shed more light on these possible links. ACKNOWLEDGEMENTS Department, supported by the British Government (DFID), when conducting the surveys. SAIAB, a national facility of the National Research Foundation in South Africa provided support for the analysis and paper preparation. Martin Villet and Tony Booth of Rhodes University gave statistical advice. Line drawings were LITERATURE CITED
APPENDIX List of specimens of Labeobarbus marequensis and Labeobarbus johnstonii used in the morphometric and meristic comparisons. Labeobarbus johnstonii SAIAB 74-108, 5 specimens, 108.1-144.5 mm SL, Lake Malawi. SAIAB 74-126, 2 specimens, 141.1 &156.7 mm SL, Lake Malawi, Varicorhinus-mouth form. SAIAB 42112, 1 specimen, 213.0 mm SL, South East Arm of Lake Malawi. SAIAB 42111, 2 specimens (large heads only), South East Arm of Lake Malawi. SAIAB 39340, 1 specimen, 133.3 mm SL, Bua River, Lake Malawi system. SAIAB 50089, 9 specimens, 41.9-111.7 mm SL, Bua River, Lake Malawi system. AMG 8401, 3 specimens, 193.0-225.0 mm SL, North Rukuru River, Lake Malawi system. SAIAB 34910, 7 specimens, 41.7-81.2 mm SL, Lufira River, Lake Malawi system. SAIAB 50174, 14 specimens, 40.5-175.0 mm SL, Wankurumadzi River, Middle Shire system. AMG 8403, 3 specimens, 117.9- 170.8 mm SL, Mpatamanga, Middle Shire River. SAIAB 34322, 1 specimen, 120.9 mm SL, Maperera River, Lower Shire system. SAIAB 34854, 8 specimens, 43.9-119.5 mm SL, Maperera River, Lower Shire system. Labeobarbus marequensis SAIAB 34938, 1 specimen, 185.0 mm SL, Chiromo, Ruo/Lower Shire River. SAIAB 51947, 1 specimen, 59.6 mm SL, Sankhulani, Ruo River, Lower Shire system. SAM 8801, 1 specimen 482.0 mm SL, (type specimen of Varicorhinus nasutus), Middle Zambezi below Victoria Falls. AMG 366, 4 specimens, 143.7-156.4 mm SL, Mazoe River, Lower Zambezi system. AMG 8pf) 33, 3 specimens, 50.6-147.7 mm SL, Mazoe River, Lower Zambezi system. AMG 2100, 1 specimen, 368.0 mm SL, Gairezi River, Lower Zambezi system. AMG (pf) 144, 3 specimens, 180.0-195.0 mm SL, Gairezi River, Lower Zambezi system. AMG 30, 6 specimens, 59.4-99.2 mm SL, Gairezi River, Lower Zambezi system. AMG 1249, 3 specimens, 39.6-60.5 mm SL, Gairezi River, Lower Zambezi system. AMG 67, 4 specimens, 70.1-121.2 mm SL, Gairezi River, Lower Zambezi system. AMG (pf) 351, 3 specimens, 83.2-89.1 mm SL, Gairezi River, Lower Zambezi system. AMG (pf) 1322, 6 specimens, 29.2-51.7 mm SL, Luangwa River, Middle Zambezi system. AMG 2022, 10 specimens, 75.6-154.3 mm SL, Kariba, Middle Zambezi system. © 2008 The South African Institute for Aquatic Biodiversity, Grahamstown, South Africa The following images related to this document are available:Photo images[sm08002f4.jpg] [sm08002f3.jpg] [sm08002f10.jpg] [sm08002f11.jpg] [sm08002f13.jpg] [sm08002t2.jpg] [sm08002f5.jpg] [sm08002t1.jpg] [sm08002f8.jpg] [sm08002t3.jpg] [sm08002f1.jpg] [sm08002f12.jpg] [sm08002f6.jpg] [sm08002f9.jpg] [sm08002f2.jpg] [sm08002f7.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}