 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
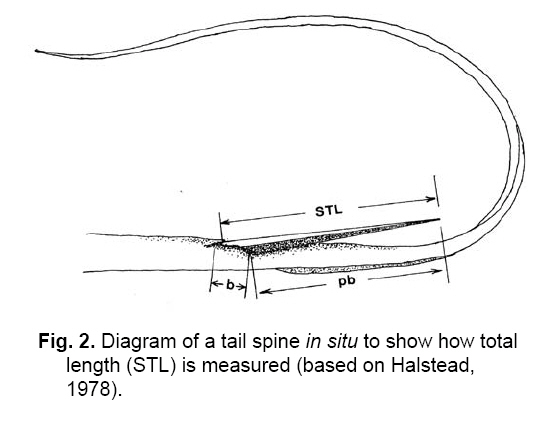
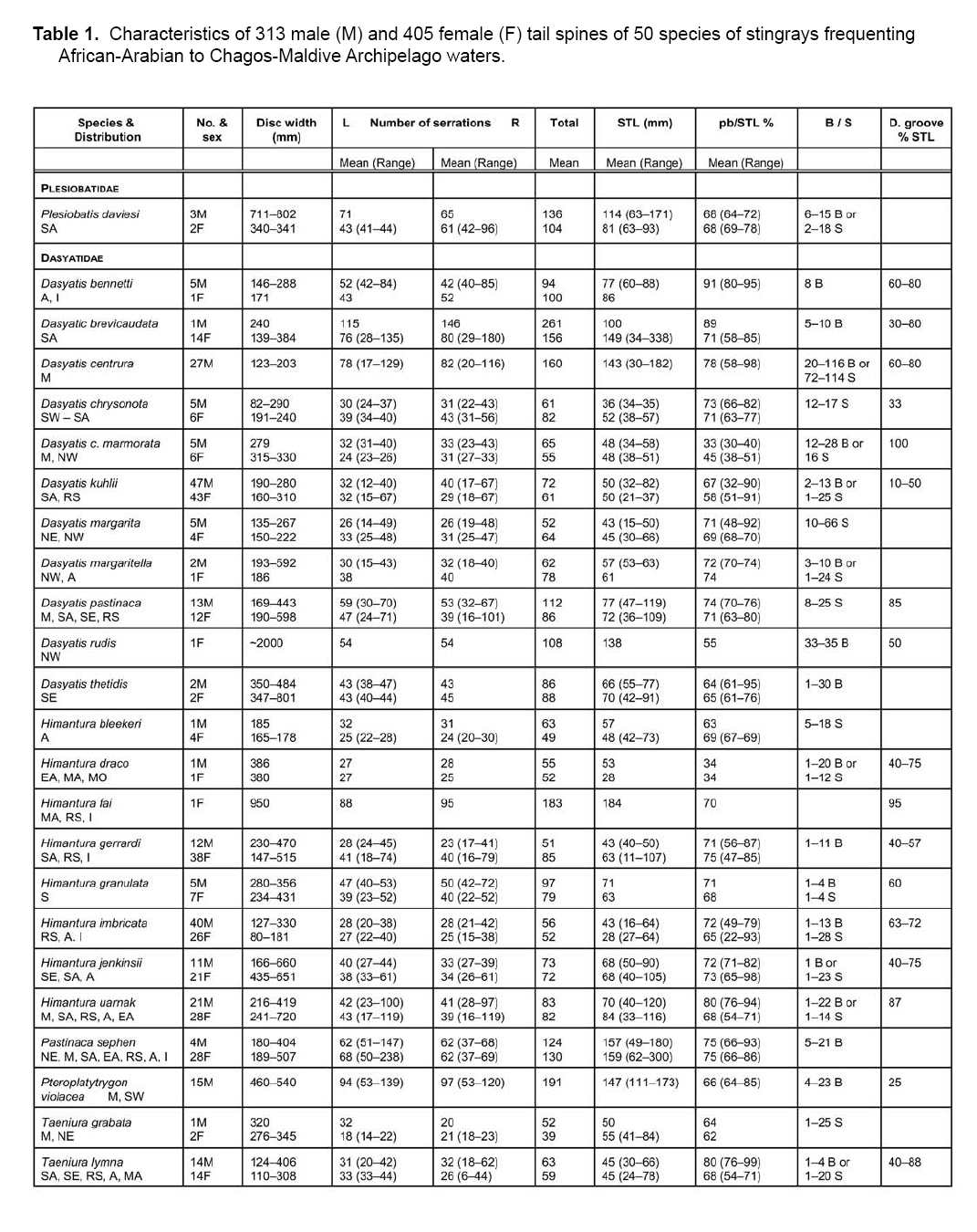
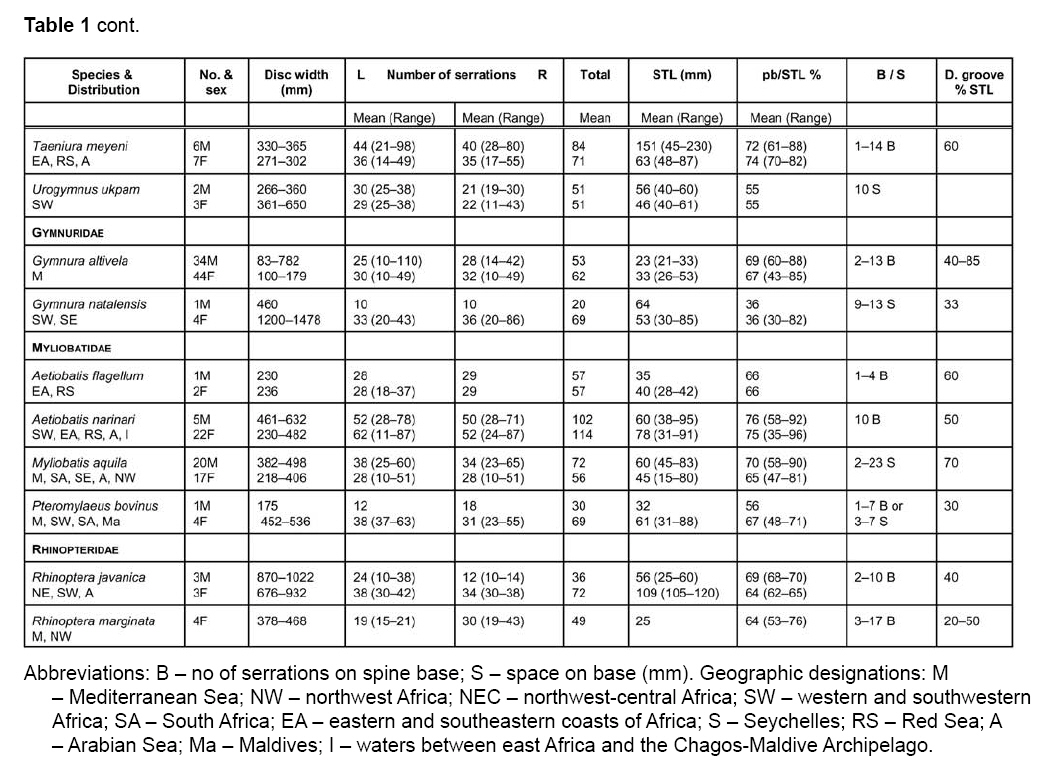
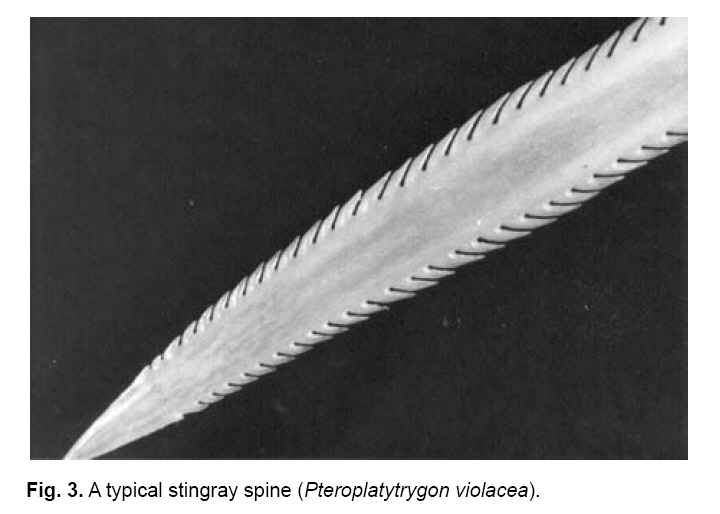
Smithiana, Publications in Aquatic Biodiversity, Bulletin 8, February, 2008, pp. 41-52 A survey of tail spine characteristics of stingrays frequenting African, Arabian to Chagos-Maldive Archipelago waters. Frank J. Schwartz Institute of Marine Sciences, University of North Carolina 3431 Arendell Street, Morehead City, NC USA 28557-3209 Submitted 12 June 2007; accepted 16 November 2007. Code Number: sm08003 ABSTRACT Tail spine characteristics were examined of 51species (including one subspecies) of stingrays frequenting African, Arabian to Chagos-Maldive archipelago waters. Thirty-four species (313 males, 405 females) possessed intact spines, 12 species lacked tail spines. Spine characteristics recorded were: total length, prebase length, serrations (right and left), serrations or spaces on the spine base and if a dorsal groove was present. Spine total serrations ranged 36-261; spine serrations of males were usually more numerous than of females, sometimes twice as many. Total serration count seemed to be associated with stingray habitat and behavior, those of active open water rays have high counts, while benthic or freshwater ray species have low counts. Disk width, spine length, prebase length and presence of a dorsal groove is not correlated with size or type of stingray. Likewise the number of spine serrations does not increase in more derived genera. A combination of spine characteristics does, however, define a species. These features will help scientists, paleontologists and physicians identify a species. KeyWords: stingrays, tail spine serrations, Africa, Arabia INTRODUCTION The public is acutely aware of injuries-even death- resulting from encounters with stingray tail spines. To date no one has examined stingray tail spines for species specific spine characteristics or species-spine-habitat correlations. Schwartz (2005), examined stingrays frequenting the Mediterranean Sea, noting each species could be identified by its spine characteristics, and that the number of spine serrations seemed to be habitat related: two species with high total spine serrations were open water swimmers (Dasyatis centroura, Pteroplatyrygon violacea); five species with intermediate serration counts (Dasyatis pastinaca, Himantura uarnak, Myliobatis aquila, Taeniura meyeni, Dasyatis margarita) were midwater or benthic inhabitants; and three benthic species had low serration counts (Gymnura altavela, Pteromylaeus bovinus, Taeniura grabata). This study reports spine characteristics of 34 stingray species frequenting the eastern Atlantic ocean waters adjacent to Africa, the Mediterranean and Arabian Seas and waters from eastern Africa to the Chagos-Maldive Archipelago, noting spine shape, total length, prebase length, right and left side serration counts and features, base serrations or space features, and presence or absence of a dorsal spine groove. The study area (Fig.1) includes the tropical ocean waters off Africa’s west coast from 10°N to 15°S, the subtropical and cool temperate waters from 15° to the Cape of Good Hope at 25°S; the tropical coastal waters of East Africa from 15°N to the Tropic of Capricorn, and the subtropical and warm temperate south to 25°S (Whitfield 2005). Fish faunal distributions are influenced by ocean currents. Fig.1 shows the prevailing currents in the study area. Two points are worth noting. The first is that during the northwest monsoon (February and March), the wind blows from southeast across the Indian Ocean and across the Indian sub-continent, inducing the development of the North Equatorial Current. A strong countercurrent exists south of the North Equatorial Current at this same time of year. In August and September, during the southwest monsoon, the North Equatorial Current reverses its flow from west to east (and is called the Monsoon Current), and the countercurrent seems to disappear. During this time of year the winds form a major upwelling off the Somali coast (Kindle & Arnone 2001). The second point is that there is no defined current in the Mozambique Channel, but that water is carried down the Channel in a series of slowly rotating eddies. The South Equatorial Current, impinges on the east coast of Madagascar at about 17°S, divides into a south flowing East Madagasar Current and a north flowing current, which flows around the north of Madagascar and contributes to the southward flowing Mozambique Eddies. The East Madagascar Current, together with the Mozambique Eddies, form part of the Agulhas Current (Lutjeharms 2007). METHODS Intact tail spines of 34 preserved stingray species and one subspecies (313 males and 405 females) were examined at the following institutions: Natural History Museum (London), Natural History Museum (Paris), Naturhistorisches Musem (Vienna), South African Museum (Cape Town, South Africa), the South African Institute for Aquatic Biodiversity (formerly the J. L. B. Smith Institute of Ichthyology, Grahamstown, South Africa), Natal Sharks Board (Umhlanga Rocks, South Africa), CSIRO Division of Fisheries (Hobart, Tasmania), Australian Museum (Sydney, Australia), United States National Museum (Washington, DC), California Academy of Sciences (San Francisco, California) and Scripps Oceanographic Institute (La Jolla, California). I follow Compagno's (2005) use of stingray common names. Most tail spines were left attached to the tail. The skin surrounding the spine base was teased loose when necessary to reveal serrations or spaces on the spine base sides. The base demarcation point was determined by placing a thin plastic ruler parallel to the spine and pushing it forward until it met the base (Fig.2). Total serrations (right [R] or left [L]), regardless of shape or size of tail spine, were counted from the spine tip to the base. Total spine length (STL) was the length from the spine tip to its posterior dorsal insertion on the tail. Prebase length (pb) was the STL minus the length of the spine from the tip to its base. Base length was the difference of the STL minus the pb. Secondary spines were noted if developing before or behind the primary spine. Disk width (DW) of each specimen was measured in millimeters (mm). Sex was noted as immature or adult. Species' spine serrations are presented alphabetically within a genus in the text and in Table 1a, b. OBSERVATIONS PLESIOBATIDAE - GIANT STINGRAYS Plesiobatis daviesi (Wallace 1967) - giant stingray. Found off southwest Africa, (Wallace 1967). A 270 cm DW species. Spine characteristics: total length averages, males 136 mm, females 81 mm; total serrations averages, males 136, females 104; pb/STL average 68% in both sexes. Six to 15 serrations or a 2-18 mm space on spine base sides; no dorsal groove evident. DASYATIDAE - WHIPTAIL STINGRAYS Dasyatis bennetti (Müller & Henle 1841) - frilltailed stingray. Frequents Persian Gulf and Hormoz Straits (Fowler 1941; Garman 1913; Vossoughi & Vosoughi 1999). A 33 mm DW species. Spine characteristics: total lengths, males 77 mm, females 86 mm; total serrations averages, males 94, females 100; pb/TL averages, males 91%, females 91%. Eight serrations on spine base sides, dorsal groove 60-80% of spine length. Dasyatis brevicaudatus (Hutton 1875) - smooth stingray. Synonyms: Dasyatis schrienei (Garrick 1954) and not D. agulhensis (Wallace 1967). South and southeast coast of South Africa from False Bay to Maputo (Wallace 1967; Compagno & Heemstra 1984; Compagno et al., 1991). A 2.1 m disk width (DW) species. A 33 mm DW species. Spine characteristics: total length averages, males 100 mm, females 149 mm (longest spine measured was of a female at 338 mm STL - see Table 1a, b); total serrations averages, males 261, females 156; pb/STL averages, males 89%, females 71%. Five to 10 serrations on spine base sides; dorsal groove 3080% of spine length. A unique feature was cul-de-sacs located medially between serrations (Fig.3), a feature also possessed by the pelagic stingray Pteroplatytrygon violacea. Dasyatis centroura (Mitchell 1815) - roughtail stingray. Occurs in the eastern Atlantic from Norway to Madeira, Mediterranean and Black Seas (Fowler 1936; Capapé 1977; Compagno & Heemstra 1984; Compagno & Roberts 1984; Dulcic et al. 2003; Golani & Capapé 2004). An active midwater to benthic 2.1 m DW species. Spine characteristics: large, heavy spines, wide at base and tapering to tip; total length averages, males 143 mm, females 159 mm; total serrations averages, males 160, females 178; pb/STL averages, males 78%, females 72%. Eleven to 20 serrations or 72-114 mm space on spine base sides; dorsal extends 60-80% of spine length; secondary spines develop below and behind primary spines. Dasyatis c. chrysonota (Smith 1828) - blue stingray. D. chrysonata is often confused with Dasyatis pastinaca and D.c. marmorata, resolved by Cowley & Compagno (1993). Known from Angola southeast to Cape Agulhas and northeast to St. Lucia, South Africa (Soljan 1948; Wallace 1967; Compagno et al. 1991; Cowley & Compagno 1993; Ebert & Cowley 2003; Heemstra & Heemstra 2004; Golani & Capapè 2004). A 48 cm DW species. Spine characteristics: total length averages, males 36mm, females 52 mm; total serrations averages, males 61, females 82; pb/STL averages, males 73%, females 71%. Twelve to 17 mm spaces on spine base sides; dorsal groove 33% of spine length. Dasyatis c. marmorata (Steindachner 1892) - blue stingray. A subspecies of D. chrysonata; found off Senegambia, West Africa, Mediterranean and eastern Atlantic from Spanish Sahara south to Congo (Maigret & Ly 1986; Cowley & Compagno 1993; Capapè & Zaouali 1995; Golani & Capapè 2004). A 27 cm DW species found in coastal benthic habitats. Spine characteristics: total length averages, males 48 mm, females 38 mm; total serrations averages, males 65, females 55; pb/STL averages 48% in both sexes. Twelve to 28 serrations or a 16 mm space on base sides; dorsal groove extends length of spine. Dasyatis kuhlii (Müller & Henle 1841) - bluespotted stingray. Found from southeastern Africa north to Red Sea (Fowler 1941; Compagno & Heemstra 1984; Compagno et al. 1991; Goren & Dor 1994; Vossoughi & Vosoughi 1999; Bonfil & Abdullah 2004). A 38 cm DW stingray. Spine characteristics: lengths 50 mm in both sexes; total serrations averages, males 72, females 61; pb/STL averages, males 67%, females 58%. Two to 13 serrations or a 1-25 mm space on base sides. Dorsal groove extends 10-50% spine length. Dasyatis margarita (Günther, 1870) - dwarf stingray. Frequents lagoons, brackish water, mangroves and estuaries from Ivory Coast to Congo, west Africa to Angola (Fowler 1936; Compagno & Roberts 1984; Maigret & Ly 1986; Compagno & Cook 1994). A 65 cm DW species. Spine characteristics: average lengths, males 43 mm, females 45 mm; total serrations averages, males 52, females 64; pb/STL averages, males 71%, females 69%. A 10-12 mm space on the base and spine proper; no dorsal groove. Dasyatis margaritella Compagno and Roberts 1984 - pearl stingray. Frequents marine and inshore waters, even off mouths of rivers, Guinea-Bissau to Congo and West Africa (Fowler 1936; Compagno & Roberts 1984). A 26 cm DW species. Spine characteristics: average lengths males 57 mm, females 61 mm; total serrations averages, males 62, females 78; STL averages, males 72%, females 74%. Three to 10 serrations or a 24 mm space may be present on base sides; no dorsal groove. Dasyatis pastinaca (Linnaeus 1758) - common stingray. Some consider D. tortonese (Capapè 1977; Golani 1996) a separate species, others consider or confuse it with D. pastinaca (Serèt, pers. comm. 2005). Found in the northeast Atlantic to Madeira, Mediterranean and Black Seas, South Africa, Angola to KwaZulu-Natal Coast, and Red Sea (Fowler 1936, 1941; Soljan 1948; Wallace 1967; Capapé 1977; Klimaj 1978; Compagno & Heemstra 1984; Compagno & Roberts 1984; Compagno & Randall 1987; Cowley & Compagno 1993, Dulcic & Lipez 2002; Golani & Capapé 2004). A 60 cm DW inshore species. Spine characteristics: average lengths, males 77 mm, females 72 mm; total serrations averages, males 112; females 86; pb/STL averages, males 74%, females 71%. Eight to 18 mm spaces on base sides; dorsal groove 85% spine length. Dasyatis rudis (Günther, 1870) - smalltooth stingray. Found off coast of Sierra Leone (Springer & Collette 1971; Compagno & Roberts 1984). A benthic 2 m DW stingray. Spine characteristics: length 138 mm; total serrations 108 short and tightly appended to spine proper. Thirty-three to 35 serrations on base sides; dorsal groove 50% of spine length. Dasyatis thetidis (Ogilby 1899) - thorntail stingray. Once described as D. agulhensis or D. lubricus (Wallace 1967). Southeastern Africa (Wallace 1967; Last & Stevens 1984; Compagno & Heemstra 1984). A large, inshore, heavy 180 mm DW species. Spine characteristics: average lengths, males 66 mm, females 70 mm; total serrations averages, males 83, females 88; pb/STL averages, males 64%, females 65%. One to 30 serrations on base sides; no dorsal groove evident. WHIPRAYS Himantura bleekeri (Blyth 1860) - whiptail stingray. Smith's (1945) H. bleekeri specimen may be H. oxyrhincha. Occurs in the Arabian Gulf (Kuronuma & Abe 1986). A 39 cm DW species found over sandy substrates. Spine characteristics: average lengths, males 57 mm, females 48 mm; total serrations, males 63, females 49; pb/STL averages, males 63%, females 69%. Five to 18 mm spaces on base sides; no dorsal groove. Himantura draco (Compagno & Heemstra 1984). No common name. Compagno (2005) considered H. draco a synonym of H. jenkensii. Stehmann (1995) and Eschmeyer (1998) believed H. draco is a valid species. Known from off Durban, South Africa at 50 m (Compagno & Heemstra 1984). DW 38-58 cm. Spine haracteristics: tail spine long, males 53 mm, females 28 mm. Total serrations are, male right 27, left 28, female right 27, left 25; pb/STL 34% for both the male and female. One to 20 serrations or one-12 mm spaces on base sides; dorsal groove 40-75% of spine length. Himantura fai Jordan & Seale 1906 - pink stingray. Occurs in the Red Sea and Maldives (Capapé 1977; Randall & Anderson 1993; Bonfil & Abdullah 2004; Randall 2005). DW 150 cm. Spine characteristics: long 184 mm tail spine; total serrations (1 female), 88 right, 95 left; pb/STL 70%. No serrations on spine base; dorsal groove 95% of spine length. Himantura gerrardi (Gray, 1851) - sharpnose stingray. Often confused with H. draco or H. jenkinsii (Wallace 1967). Found from South Africa to Red Sea and Arabian Sea (Fowler 1941; Wallace 1967; Compagno & Heemstra 1984; Kuronuma & Abe 1986; Compagno & Randall 1987; Goren & Dor 1994; Stehmann 1995; Randall 1995; Randall & Lim 2000; Bonfil & Abdullah 2004). A 90 cm DW species. Spine characteristics: average lengths, males 43 mm, females 63 mm; total serrations averages, males 51, females 81; pb/STL averages, males 71%, females 75%. One to 11 serrations on base sides; dorsal groove 40-57% of spine length. Two forms may be encountered: tail may be banded or mottled. Himantura granulata (Macleay 1883) -mangrove whipray. Known from Seychelles, Oman and Maldives (Randall & Anderson 1993; Randall 2005). A 90 cm DW stingray. Spine characteristics: average lengths, males 71 mm, females 63 mm; total serrations averages, males 97, females 79; pb/STL averages, males 71%, females 68%. One to four serrations or a one to four mm space on base sides; dorsal groove 60% spine length. Himantura imbricata (Bloch & Schneider 1811) - scaly stingray: Synonyms are Trygonoptera walga and Trygon panopeus. Found in the Red Sea and Arabian Gulf (Fowler 1941; Goren & Dor 1994; Khalaf & Desi 1997; Kuronuma & Abe 1986; Compagno & Randall 1987; Randall 1995; Bonfil & Abdullah 2004). A dwarf species with 23 cm DW. Spine characteristics: average lengths, males 43 mm, females 28 mm; total serrations averages, males 56, females 52; pb/STL averages, males 72%, females 65%. One to 13 serrations or a 1-28 mm space on base sides; dorsal groove 63-72% spine length. Himantura jenkinsii (Annandale 1909) - pointed-nose stingray. Often considered to be Himantura draco or H. gerrardi (Compagno & Heemstra 1984; Stehmann 1995; Randall 1995; Heemstra & Heemstra 2004). A 100 cm DW stingray. Occurs off KwaZulu-Natal, southeastern Africa (Wallace 1967). Spine characteristics: average lengths 68 mm in both sexes; total serrations averages, males 73, females 72; pb/STL averages, males 72%, females 73%. One serration or a 1-23 mm space on base sides. Himantura uarnak (Forsskål 1775) - honeycomb stingray. A systematically confused species often considered to be H. oxyrynchus, H. krempfi or even H. bleekerii (Eschmeyer 1998; Deynat & Fermon 2001). Found in the Mediterranean, Red Sea, Arabian Sea, Persian Gulf and along the east coast of Africa (Fowler 1941; Smith 1955; Smith & Smith 1963; Wallace 1967; Kuronuma & Abe 1980; Compagno & Heemstra 1984; Compagno & Randall 1987; Goren & Dor 1994; Khalaf & Desi 1997; Capapè & Zaouali 1995; Randall 1995; Basusta et al. 1998; Heemstra & Heemstra 2004; Bonfil & Abdullah 2004; Golani & Capapé 2004). A highly active 110 cm DW species. Spine characteristics: average lengths, males 70 mm, females 84 mm; total serrations averages, males 83; females 82; pb/STL averages, males 80%, females 68%. One to 22 serrations or a 1-14 mm space on base sides; dorsal groove 87% of spine length. Body may be spotted or reticulate and tail may be banded or plain. FEATHERTAIL STINGRAYS Pastinachus sephen (Forsskael, 1775) - cowtail stingray. Found off northeast Africa, Red Sea to Chagos Archipelago (Fowler 1941; Smith 1955, 1967; Smith & Smith 1963; Wallace 1967; Kuronuma & Abe 1986; Compagno & Heemstra 1984; Compagno & Randall 1987; Winterbottom et al. 1989; Randall & Anderson 1993; Goren & Dor 1994; Compagno & Cook 1994; Randall 1995; Bonfil & Abdullah 2004). A 180 cm DW stingray. Spine characteristics: average lengths, males 157 mm, females 159 mm; total serrations averages, males 124, females 130; pb/STL averages, males 75%, females 76%. Five to 21 small serrations on base sides, no dorsal groove evident. PELAGIC STINGRAYS Pteroplatytrygon violacea (Bonaparte 1832) - pelagic stingray. Often confused with D. purpureus (Winterbottom et al. 1989). Found in the Mediterranean and along the western and southern African coasts (Soljan 1948; Tortonese 1976; Capapé 1977; Compagno & Roberts 1984; Compagno & Heemstra 1984; Compagno et al. 1991; Basusta et al. 1998; Mollet 2002, Golani & Capapé 2004). A broad (80 cm DW), open ocean stingray, dark dorsally and ventrally, anteriorly arched in profile. Spine characteristics: average lengths, males 147 mm, females 128 mm; total serrations averages, males 191, females 161; pb/STL averages, males 66%, females 78%. Three to 23 serrations or a 7-9 mm space on base sides; dorsal groove 25% of spine length. The even-sided spine is characterized by culde-sacs medially between serrations, similar to Dasyatis brevicaudatus (Fig.3). RIBBONTAIL STINGRAYS Taeniura grabata (Geoffroy St. Hilaire 1817) - round fantail stingray. Known from the Mediterranean Sea, eastern Atlantic and eastern Africa (Fowler 1936, 1941; Maigret & Ly 1986; Biscioto & Wirtz 1994; Compagno & Randall 1987; Dingerkus 1995; Basusta et al. 1998; Serena et al. 1999; Golani & Capapé 2004), erroneously from the Red Sea (Fowler 1936; Compagno & Randall 1987). A ~100 cm DW stingray. Spine characteristics: average lengths, males 50 mm, females 56 mm, tip upturned; total serrations averages, males 52; females 39; pb/STL averages, males 64%, females 85%. One to 25 mm space on base sides, extending onto spine proper; dorsal groove 90% of spine length. New spines often develop ahead of primary attachment end; spine end V-shaped. Taeniura lymma (Forsskål 1775) - blue-spotted stingray. Western Indian Ocean (Fowler 1941; Smith 1955; Smith & Smith 1963; Wallace 1967; Compagno & Heemstra 1984; Goren & Dor 1994; Compagno & Randall 1987; Randall & Anderson 1993; Khalaf & Desi 1997; Basusta et al. 1998; Heemstra & Heemstra 2004; Bonfil & Abdullah 2004). An oval-shaped 30 cm DW stingray. Spine characteristics: average lengths 45 mm STL in both sexes; total serrations, males 63, females 59; pb/STL averages, males 80%, females 68%. One to four serrations or a 1-20 mm space on base sides; dorsal groove 40-88% of spine length. Taeniura meyeni (Müller & Henle 1841) - fantail stingray. Frequents eastern Africa, Madagascar northward to Red Sea, Arabian Sea and Persian Gulf (Fowler 1941; Randall & Anderson 1983; Compagno & Roberts 1984; Bonfil & Abdullah 2004; Randall 2005). A dangerous 180 cm DW species. Spine characteristics: average lengths, males 151 mm, females 63 mm; totals serrations averages, males 84, females 71; pb/STL averages, males 72%, females 74%. One to 10 serrations on base sides; dorsal groove 80% of spine length. PORCUPINE RAYS Urogymnus ukpam (Smith, 1863) - thorny freshwater ray. Often placed in Dasyatis because of tail size or lack of tail spine in some specimens (Compagno & Roberts 1984; Compagno & Cook 1994). Known from freshwaters of west Africa from Cameroon to Congo (Compagno & Roberts 1984; Compagno & Randall 1987). A large 65 cm DW species. Spine characteristics: average lengths, males 56 mm, females 46 mm; total serrations averages 46 in both sexes; pb/STL averages 55% in both sexes. A 10 mm space on base sides; dorsal groove 33% of spine length. GYMNURIDAE - BUTTERFLY RAYS Gymnura altavela (Linneaus 1758) - spiny butterfly stingray. Found in northeast Atlantic from Norway to Madeira, Mediterranean and Black Seas (Fowler 1936; Soljan 1948; Bigelow & Schroeder 1953; Hureau & Monod 1973, Maigret & Ly 1986; Dulcic et al. 2003). A >64 cm DW benthic stingray. Spine characteristics: average lengths, males 23 mm, females 33 mm; total serrations averages, males 53, females 62; pb/STL averages, males 69%, females 67%. Two to 23 serrations on base sides; dorsal groove 40-85% spine length. Gymnura natalensis (Gilchrist & Thompson 1911) - diamond ray. Known from south and eastern South Africa (Wallace 1967; Heemstra & Heemstra 2004). A 182 cm DW stingray. Spine characteristics: average lengths, males 67 mm, females 53 mm; total serrations averages, males 20, females 69; pb/STL averages 36% in both sexes. A 9-13 mm space on base sides; dorsal groove 33% of spine length. MYLIOBATIDAE - BONNETRAYS Aetobatus flagellum (Bloch & Schneider, 1801) - long-headed stingray. Frequents Red Sea (Vossoughi & Vosoughi 1999; Randall & Lim 2000: Bonfil & Adullah 2004). A 62 cm DW stingray. Spine characteristics: average lengths, males 35 mm, females 40 mm; total serrations averages 57 in both sexes; pb/STL average 66% in both sexes. One to four serrations on base sides; dorsal groove 60% spine length. Aetobatis narinari (Euphreson 1790) - spotted eagle ray. Widely distributed along west and east African coasts, in Red Sea and Arabia Gulf (Fowler 1925, 1936, 1941; Smith 1955; Smith & Smith 1963; Wallace 1967; Klimaj 1978; Kuronuma & Abe 1986; Compagno & Randall 1987; Goren & Dor 1994; Stehmann 1995; Randall 1995; Kahlaf & Desi 1997; Bonfil & Abdullah 2004; Randall 2005). A 330 cm DW active, open water species. Spine characteristics: average lengths, males 60 mm, females 78 mm; total serrations averages, males 102, females 114; pb/STL averages, males 76%, females 75%. Ten serrations on base sides; no dorsal groove evident. Usually spotted dorsally, yet Last and Stevens (1984:448) depict an individual spotted on the rear two-thirds of the dorsum. The same is true for three specimens at Vienna, while one specimen at Hobart (CSIRO) is spotless. May possess up to eight tail spines (Gudger 1914). EAGLE RAY Myliobatis aquila (Linnaeus 1758) - cowtail stingray. Often described as Himantura cervis. Found in the Mediterranean, south and east African coasts and Arabian Sea (Fowler 1936, 1941; Soljan 1948; Wallace 1967; Klimaj 1978; Compagno et al. 1991; Lens & Marin 1993; Heemstra & Heemstra 2004). A 83 cm DW species. Spine characteristics: average lengths, males 60 mm, females 45 mm; total serrations averages, males 72, females 56; pb/STL averages, males 70%, females 65%. Two to 23 mm spaces on base sides and onto spine proper; dorsal groove 70% spine length. BULL RAY Pteromylaeus bovinus (Geoffroy St. Hilaire, 1817) - bullray. Frequents Mediterranean, western Africa to Namibia and South Africa to Mozambique (Fowler 1936, 1941; Wallace 1967; Maigret & Ly 1986; Garman 1913; Heemstra & Heemstra 2004). A 1.1 m DW stingray. Characteristics are: spine lengths are males 32 mm STL, females 61 mm STL. Serrations total: males 30, females 69. pb/STL's average: males 56%, females 67%. One to seven serrations or a 3-7 mm space on base sides. Dorsal groove 30% of spine length. COWNOSE RAYS Rhinoptera javanica (Müller & Henle 1841) - Javanese cownose ray. Synonyms include R. peli Bleeker 1863, R. jayakari (Boulenger 1895) and R. sewelli Misra 1947 (type specimens of last two species are now at the NC State Museum, Raleigh, NC; Schwartz 1990). A midwater to benthic inhabitant from equatorial Africa, southeast Africa, Gulf of Oman and Arabia (Fowler 1934; Smith & Smith 1963; Wallace 1967; Kuronuma & Abe 1986; Schwartz 1990; Vossoughi & Vosoughi 1999). A 195 cm DW species. Spine characteristics: average lengths, males 56 mm, females 109 mm; total serrations averages, males 36, females 72; pb/STL averages, males 69%, females 64%. Two to 10 serrations on spine sides, dorsal groove 40% of spine length. Rhinoptera marginata (Geoffroy Saint-Hilaire 1817) - Lusitanian cownose ray. Synonyms R. peli Cadenat 1960; Quero et al. 1990 and R. bonasus Séret 1990. Found in the Mediterranean and estuaries of northwest African coast (McEachran & Capapé 1984-1986; Ruitart 1998; Schwartz 1990). A 2 m DW pelagic-benthic species. Spine characteristics: average lengths (females) 25 mm; total serrations average, for females 49; pb/STL average 64%. Spine flat with 3-17 serrations on base sides; dorsal groove 20-50% spine length. DISCUSSION Tail spine characteristics are described based for 34 preserved species, one subspecies, and 718 specimens (313 males, 405 females) that possessed tail spines. Spine serration differences existed in relation between a species and the habitat it frequented: species with a total spine serration count of >100 were ocean or pelagic inhabitants (e.g. Dasyatis centroura and Pteroplatytrygon violacea); species with 70-100 total spine serration were midwater swimmers (e.g. Dasyatis pastinaca, Himantura uarnak and Myliobatis aquila); those with 50-70 serrations totals were benthic inhabitants (e.g. Dasyatis chrysonota, Gymnura altavela, Taeniura grabata, Pteromylaeus bovinus), and those with counts of <50 were usually freshwater inhabitants (e.g. Urogymnus ukpam) (Table 1a, b). The spines of Dasyatis brevicaudata and Pteroplatytrygon violacea are distinct, possessing cul-desacs medially between serrations (Fig. 3). No primitive to advanced species-spine serration relationship (as in Dasyatis, Himantura, Gymnura, Myliobatis, Rhinoptera and Mobula (Compagno 2005) was evident. Likewise, Plesiobatis daviesi, considered a primitive species (Compagno 2005), has tail spines with 120 serrations while the spine serrations of 11 species of dasyatids range from 58 to 209, for D. marginata and D. bennetti respectively, and spine serrations of 7 species of Himanturids varied from 54 to183, for H. imbricata and H. fai, respectively. Counts for the other species are summarised in Table 1a, b. Comparing tail spine serrations characteristics of 11 stingray species reported by Schwartz (2005) with this study, three species' spine serrations totals were the same (D. centroura, G. altavela and U. upkam). Of the three, U. upkam enters freshwaters of West Africa. Total tail spine serrations for H. bovinus were lower off Africa than in the Mediterranean Sea (36, this study 44), while serration totals of the remaining seven spines were greater off Africa-Arabia than in the Mediterranean Sea (Schwartz 2005, Table 1a, b). Twelve other species, also found in African-Arabian waters, do not possess tail spines: Aetomylaeus nichofii, A. milvus, A. maculatus, A. vespertilio, Gymnura micrura, Mobula rochebrunei, Mobula topacana, Mobula thurstoni, Mobula kuhlii,Urogymnus asperrimus, Mobula ergoodootenkee and Manta birostris. They represented members of advanced stingray families that exhibited a gamut of swimming and habitat usages, suggesting that advanced stingrays are tending to lose tail spines with evolution. Spines of specimens of Hexatrygon bickelli and Dasyatis garouaensis (the latter an inhabitant of the Niger and Cameroon Rivers, West Africa) were broken and thus not counted. Mobula mobular, Gymnura poecilura and Aetoplatea tentaculata which possess tail spines, were not seen. However, examination of the spines of other Hexatrygon 'species' specimens in Taiwan, Japan and Tasmania (designated species 'I' or 'H' at the time), revealed that there seem to be two species involved. Specimens labelled 'H' have high serration counts (84 or more) and smaller spaced on the base sides, while 'I' specimens have lower counts (64-75) and large spaces on base the sides. CONCLUSIONS Much taxonomic confusion in general still persists regarding the stingrays inhabiting African-Arabian waters. For example, Himantura uarnak is a confusing species complex that Deynat and Fermon (2001) resurrected as H. oxyrhyncha, a species found in Thailand, Sumatra and Borneo, from its synonymy and H. krempfi, a freshwater species from Thailand, Cambodia, Indonesia and Mekong, as a junior synonym. Compagno (2005) listed H. oxyrhynchus as a full species. An H. oxyrhyncha specimen in Paris lacked a spine and three of five H. krempfi specimens possessed tail spines, their characteristics were: spine length 32, 58 and 60 mm STL; serrations numbered 33, 44 and 52 respectively; 4 serrations or a 20 mm space occurred on base sides, and no dorsal groove was evident. While stingrays are large, bulky and feared because of their venomous tail spines that may cause injury or death, stingray spine characteristics are distinct within a species and should not be overlooked when describing a species. Their inclusion in a species diagnosis adds another parameter that helps physicians, paleontologists and geologists with species identity and fossil relationships. ACKNOWLEDGMENTS Thanks are due many who made specimens under their care available or provided valuable information and literature pertaining to the species studied. They are: Drs D. Golani and M. Goren (Israel); Dr B. Séret (Paris); Dr E. Mikschi, H. Wellendorf and staff (Vienna); Dr L.J.V. Compagno (Cape Town, South Africa); J. Wallace (Durban, South Africa); Dr P. Heemstra and A. Bentley (Grahamstown, South Africa); J. Cliff and S. Dudley (Umhlanga Rocks, South Africa); Drs P. Last and J. Stevens (CSIRO, Hobart, Australia); Dr J. Paxton, (Sydney, Australia); Dr J. Randall (Hawaii); Dr P. Hasting, H. Walker and C. Klepedlo (Scripps, La Jolla, California); Drs W. Eschmeyer, T. Iwamoto, D. Cantana and W. Poly (California Academy Sciences, San Francisco, California); Dr J. Williams and S. Jewett (U.S. National Museum, Washington, DC); M. Sabaj and M. Littman (Philadelphia Academy Natural Sciences, Philadelphia, Pennsylvania); K. Hartel (Museum Comparative Zoology, Cambridge, Massachusettes), Dr R. Winterbottom (Royal Ontario Museum, Toronto Canada) and Dr N. P. Parin and P. P. Shershov (Institute Oceanography Moscow, Russia) who measured the 'H. draco' in that collection. Librarians P. Marraro (NOS, Beaufort, NC) and B. Bright (IMS, Morehead City, NC) expedited needed literature references. L. White (IMS, Morehead City, NC) translated my scribbles into the readable text. MATERIAL EXAMINED Institutional abbreviations follow Leviton et al. 1985. Plesiobatis daviesi: AMS 4127, CAS 3350.1, 1104127.1, CSIRO 4107.1, ORI B865 Dasyatis bennetti: MCZ 40418, USNM 13785, 193662, 19781, 21618. Dasyatis brevicaudata: ANSP 167402, 16704, 167405, 167409, CAS 26470, 27449, MCZ 23860, SI 0457, 804861.001, 35710.00, I 35900.02, IB 7911, BRI 805, CSIRO 1B07911 Dasyatis centroura: MCZ 562, 203759, NMW 50202, 77995a, 88086, 32 uncatalogued. Dasyatis chrysonata: CAS C314, 117, 1334, 41536, 1 uncatalogued, HUJ 13768.12450, RUS 131823. Dasyatis marmorata: NMW 82939, 89680. Dasyatis kuhli: AMS 1B122, ANSP 121539, 127372, 131644, 15851, 171543, 171544, 171545, 171546, 171641, 25843, CAS 104, 270, 278, 280, 310, 21664, 29993, 3237, 60773(3), 61832, 68134, 68137, 68144, 60-252, 60-373, CSIRO 197671, I 11131, 21038.09, H4122.3, H4961, 4926(2), MCZ127372, 22154, 22546, MTUF 2070, NMW 50218, 86966, 90201, 94588, SI 1131, 118180, 19767, 21844, 504, S1982.00, I 1653, 10123, 14878, I1652, 3482.001, 21833.013, USNM 206144, 222533, 222545, 222546. Dasyatis margarita: MNHN 2605(2), 3826, 7954(4), NMW 6241, 50241. Dasyatis margaritella: MCZ 459, 222589, 222592, NMW 3837, 22407, 50200, 50203, 61326, 68326, 78007, 87250, 87273, USNM 004595, 193622, 222589, 222592. Dasyatis pastinaca: ANSP 29998, 350984, CAS 9865, 110560, 12110, 12830, 12845, 19880, NMW 77972, 78931, 79498, 83082, 86608. Dasyatis rudis: USNM 202730. Dasyatis thetidis: CAS 40960, 67575, SI 3047.001, B 6518, H3228.2. Himantura bleekeri: USNM 222606, 222610, 222624(6). Himantura draco: RUIS 1996(2), ZMMU -P 19174. Himantura fai: USNM 051712. Himantura gerrardi: ANSP 172801,BPBM 33201, 33202, 33199, CAS 141, 590, 680, 808, 2437, 2460, 2568, 7888, 20960, 41048, 52984, 63045, 63085, 63086, 63097, 68132, 68138, 68142, 68144, 68145, 68152, 68195, 75869, 12772, 60-28, 60-129, 60-205, 60-326, 60-400, LACM 38130.47, 38113.48, NMW2003.0025, 61774. Himantura granulata: CAS 3097, 41048, 63006, 68144, 68145, 68149, 68152, 75869, LACM 8311, 38130.47, NMW 2003.0025, 60774. Himantura imbricata: ANSP 03108, BPBM 33199, CAS 224(3), 1347, 2223, 2224, 4658, 27435, 48660, 34295, 41675, 41677, 41678, 41680, 41681, 41682, 41683, 41684, CSIRO 20762.001, I 2867, IB 457, MCZ 223, 224(3), C 123(22), SAM 41635, USNM 222529, 222552, 222561, 222564. Himantura jenkensii: CAS 2645, 7855(2), 68131, 68133, 68135, 68136, 68142, 68148. Himantura uarnak: ANSP 25842, 25843, CAS 1060, 2433, 2525, 8807, 63093, 63095, 68111, 68144, 68150, 68151, MCZ 201806, MTUF 25070, NMW 1897, 8972, 22453, 22459, 37325, 60176, 60773, 60776, 65715, 66799, 78972, 81604, 86650, 86659, 87125, 87352, 87329a, 87052a, 87852, USNM 119225, 148100, 201806, 206132, 207045, 220153, 232879, 33700. Pastinaca sephen: ANSP 103715(3), CAS 6155, 65240, 68155, CSIRO 2213.11, H4116.04, 4213.02, 114172.4, MTUF 25108, 26701, 26711, 26712, 26899, 26902, NMW 0711, 2017, 2021, 60759, SI 1908, USNM 147420, 205138, TUFIL 26701, 26711. Ptryoplatytrygon violacea: MTUF 3425, 106471, 11056, 67675, 74436, MZUP 49011, NMW 42672, 88604, 191234, S10, 29-308, 33-35, 58-35, 54-38, 54-93, 63-120, 73-418, 79-51, 76-21, 79-59, 79-275, 79-283(3), 79-219, 79295, 79-320, 79-309, NMW H2672, 88604, 191234, USNM 222566, 222646. Taeniura grabata: BPBM 9204, 9205, 9206, HVJ8359, 12418. Taeniura lymna: CAS 60-160, HV224, MCZ 1251, 40480, NMW 2645, 8791, 50223, 78008, 78020, 78189, 87178, 87189, 87188, 3 uncatalogued, S10, 4568, SI 14564, I 5067.03, 13832, 107687, 197785, USNM 40436, 49326, 222621, 226314, 262618. Taeniura meyeni: CAS 63103, CSIRO I 17314.001, NMW 77976, 89471, SI 17364.000. Urogymnus upkam: CAS 42761, USNM 219780. Gymnura altavela: NMW 2008, 2009, 50231, MZUP 9721, 9914, 9918, 10380, USNM 20261, 20762, 70 uncatalogued. Gymnura natalensis: CAS 14025, 1 uncatalogued, LJVC 682023, 824023, SAM 25019, 33293, SAS 25017. Aetobatis flagellum: USNM 206131, 22684, 256190. Aeotbatis narinari: ANSP 71284, 81715, 10224, 179204, CAS 11675, 62395, 63105, 63111, 68154, 71650, 113807, LJVC 87114, MTUF 4343, 110569, 167675, B9166, 26700616, NMW 6073, 10755, 66725, 87161, 87246, 82829, 88207, 88556, 89551, USNM 63175, 205141, 222689. Myliobatis aquila: MCZ 150-S, 828, 89927, NMW 3857, 6653, 8531, 50236, 50237, 60780, 81159, 87165, 87195, 87366, 87844, 886605, RUSI 04804, 10423, 19823, 444352, SAM 12842, 26452, 26528, 33294, 32575, 333453, 34392, USM 030, 202823, 203457, 22577. Pteromylaeus bovinus: NMW 89825, USNM 202709, 202763, 222709, 232329. Rhinoptera javanica: China 34916, MCZ 316, NCSM 2 uncatalogued types, ORI 0347, B646, SCB 98006. Rhinoptera marginata: MNHN 2605(2), 3824, 7954(4), NMW 50241, 56241, 59055. LITERATURE CITED
© 2008 The South African Institute for Aquatic Biodiversity, Grahamstown, South Africa The following images related to this document are available:Photo images[sm08003f2.jpg] [sm08003f3.jpg] [sm08003t1a.jpg] [sm08003f1.jpg] [sm08003t1b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}