 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
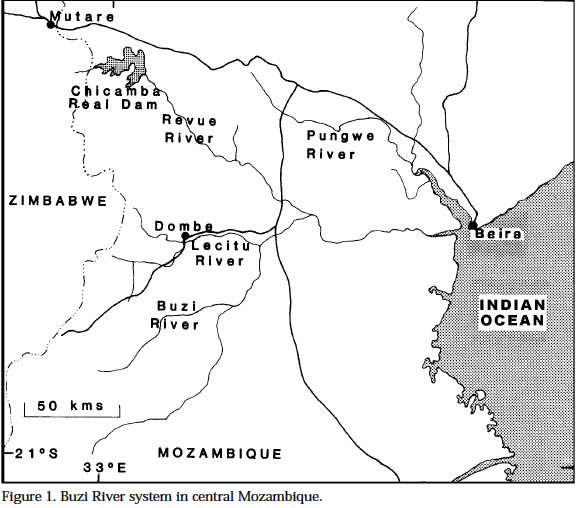
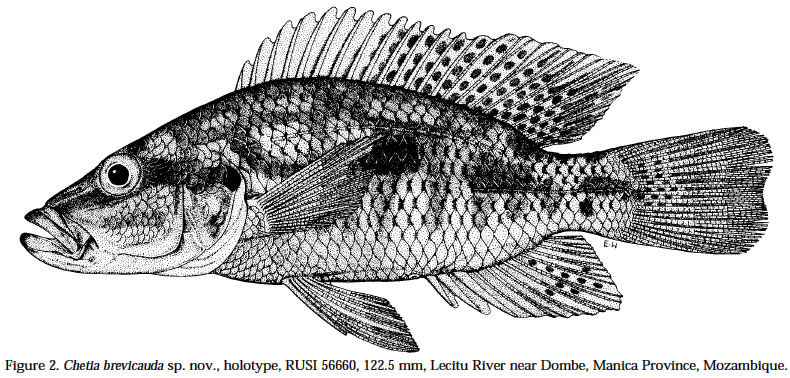
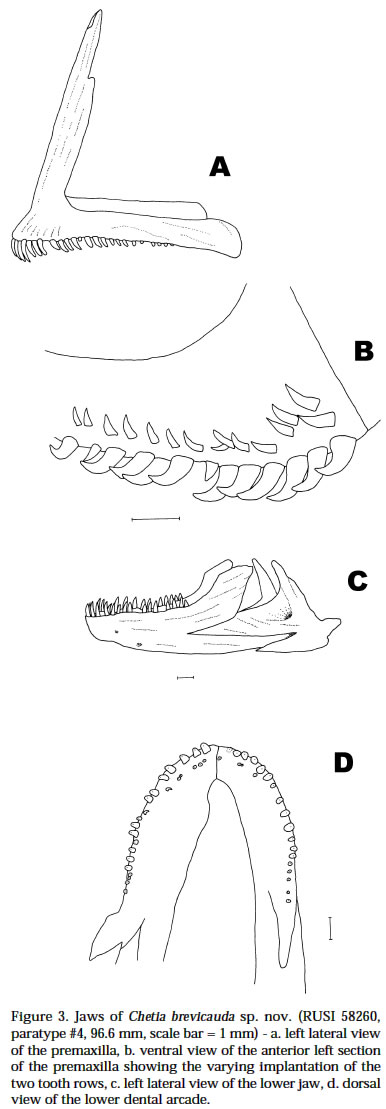
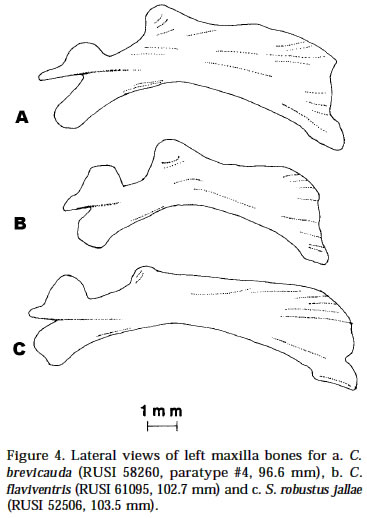
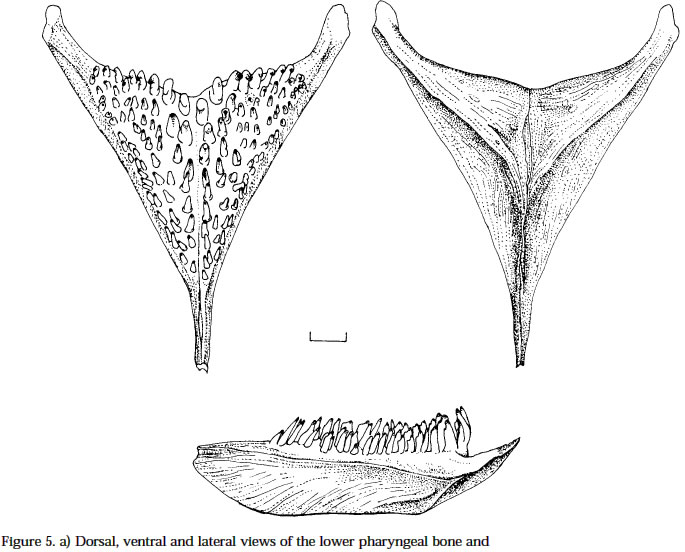
Smithiana, Publications in Aquatic Biodiversity, Special Publication 1 July, 2002, pp. 1-13 A New Species of the Genus Chetia (Teleostei: Cichlidae) from the Lecitu River, Buzi System, Mozambique. I. Roger Bills 1 & Olaf L.F. Weyl 2 1 South African Institute for Aquatic Biodiversity, Private Bag 1015, Somerset Street, Grahamstown, 6140, South Africa. (e-mail: R.Bills@ru.ac.za) - Corresponding author. 2 Department of Ichthyology, Rhodes University, P O Box 94, Grahamstown, 6140, South Africa. Current Address -National Aquatic Resource Management Programme (NARMAP), P O Box 27, Monkey Bay, Malawi. Code Number: sp02001 ABSTRACT A new cichlid species, assigned to the genus Chetia, is described from the Lecitu River (Buzi system) in Manica Province, Mozambique. The diagnostic features for this species are: deep body (37.4-41.6% SL); large head (39.4-41.0% SL); long pectoral fin (25.0-28.4% SL); low number of lateral line scales (31-32); reduced number of caudal vertebrae (14); a short and deep caudal peduncle (length/ depth ratio 1.0). The new species is compared to other members of the genus and some other serranochromines. The value of certain diagnostic features and the distribution of the genus Chetia are discussed. Additions are made to the Buzi River system ichthyofaunal checklist. INTRODUCTIONThe Buzi River system (Fig. 1) in central Mozambique has an essentially lower Zambezi fish fauna (Bell-Cross 1973). The Buzi and Pungwe Rivers are linked in their lower reaches which flood during some wet seasons and the Pungwe itself is linked to the lower Zambezi through a swamp region in the western sector of the Gorongoza National Park. Biogeographically, the Buzi system is of interest because our present knowledge indicates that it forms the southern boundary for several lower Zambezi species e.g. Malapterurus shirensis Roberts 2000, Heterobranchus longifilis Valenciennes 1840, Distichodus schenga Peters 1852, D. mossambicus Peters 1852, Mormyrops anguilloides (Linnaeus 1758) and Hippopotamyrus ansorgii (Boulenger 1905) (Bell-Cross 1972 & 1973, Roberts 2000). These species, while characteristic lower Zambezi fauna, have affinities with species presently found in the Zaire system and are not found in the upper Zambezi and many of its western elements (Okavango and Kafue rivers). In addition, the Buzi River is the type and only known locality for the claroteid catfish Amarginops hildae (Bell-Cross 1973) another species which has close relatives in Zairean and west African river systems (Nichols & Griscom 1917). The serranochromine cichlids (Greenwood 1993) are one of the characteristic upper Zambezi groups and are relatively poorly represented down the east African coast. Serranochromis meridianus Jubb 1967, Chetia flaviventris Trewavas 1961 and Chetia brevis Jubb 1968 occur in South Africa and southern Mozambique but all have restricted distributions even within the systems in which they occur. Except for the Shire River in Malawi, where Serranochromis robustus robustus (Günther 1864) occurs, there are no other Serranochromis Regan 1920 in the lower Zambezi or associated systems. During an angling expedition to the Lecitu River (Buzi system, 19° 55' S 33° 22' E) in January 1997, one of us (OW) collected six specimens of an undescribed serranochromine cichlid. The specimens belong to the genus Chetia Trewawas 1961 as redefined by Greenwood (1993). The purpose of this paper is to describe this new species. Comparisons are made with other members of the genus Chetia and certain characters diagnosing the genus are reviewed. The distribution of the genus raises issues of historical linkages with the Buzi and other southern African river systems. METHODSThe fish were collected by angling using a number four hook with live earthworms as bait. The specimens were fixed in a 10% formalin solution and preserved in 70 % ethyl alcohol. The morphometric measurements and meristic counts follow Barel et al. (1977). Lengths given in the text are SLs unless otherwise stated. All counts were taken from the left side of the fish. Each specimen was radiographed (radiographs are held at the South African Institute for Aquatic Biodiversity, Grahamstown) to determine vertebrae numbers and fin elements. Dorsal and anal fin ray numbers were determined by counting the proximal portions of rays and pterigiophores. Last anal and dorsal fin rays were counted as one ray if they were supported by a single pterigiophore. The abbreviations of institutions follow Leviton et al. (1985). The material examined was from the following institutions: South African Institute for Aquatic Biodiversity, Grahamstown, South Africa (RUSI); the Albany Museum, Grahamstown, South Africa (AMG); Natural History Museum, London, U.K. (BMNH); and the Royal Ontario Museum (ROM), Canada. HOLOTYPE. RUSI 56660, 122.5 mm SL, O. Weyl, Lecitu River at Dombe pontoon (19° 55’S 33° 22’E), Manica Province, Mozambique, 20/January/1997. PARATYPES. RUSI 58260, (5) 82-125 mm SL, collected with holotype. DIAGNOSIS: A member of the genus Chetia distinguished from other members of that genus by the following characteristics. Head large 39.4-41.0% SL vs 34.4-36.0 in C. mola and 33.0-37.0 in C. brevis. A deep bodied species 37.4-41.6% SL vs. 32.4-35.9 in C. mola, 33.3-34.2 in C. welwitschi and 29.0-37.0 in C. brevis. Pectoral fin relatively long 25.0-28.4% SL showing some overlap with C. brevis 18.0-26.0 but greater than C. flaviventris 21.7-24.7, C. mola 22.8-23.3 and C. welwitschi 18.1-20.6. Lateral line scales 31-32 vs 32-34 in C. brevis and C. welwitschi, 34 in C. mola and 34-35 in C. flaviventris. Caudal vertebrae are low in number with 14 in all specimens vs 15 in C. welwitschi and 16-18 in C. mola and C. flaviventris. The caudal peduncle is short and deep with a mean CPL/CPD ratio of 1.0 vs 1.3-1.5 in C. flaviventris and 1.4-1.5 in C. welwitschi. COUNTS AND MEASUREMENTS: Holotype values first followed by range of paratypes in parentheses. Measurements in percent SL: head length 39.4 (39.6-41.0); body depth 37.4 (37.4-41.6); predorsal length 38.9 (39.7-43.0); preanal length 68.2 (68.4-71.3); pectoral fin length 25.0 (26.2-28.4); pelvic fin length 25.6 (26.2-27.8); posterior dorsal spine length 13.5 (13.3-14.7); highest dorsal soft ray 23.1 (17.8-25.1); posterior anal spine length 12.1 (11.9-14.1); highest anal soft ray 21.2 (20.2-23.9); caudal peduncle length 12.7 (12.8-13.9); caudal fin length 27.8 (28.4-30.2). Measurements in percent HL: snout length 32.3 (28.8-34.6); preorbital depth 19.0 (16.1-19.7); eye length 20.3 (20.1-23.6); inter-orbital width 17.2 (17.6-18.6); cheek depth 30.2 (27.5-33.3); lower jaw length 46.8 (44.5-49.3); upper jaw length 37.9 (34.5-39.4). Ratios: caudal peduncle length/ caudal peduncle depth 1.0 (1.0); caudal fin scales/ caudal fin length 0.6 (0.4-0.6). Counts: upper lateral line scales 21 (20-22); lower lateral line scales 11 (10-12) total LL scales 32 (31-32); cheek scale rows 5 (4-5); scales round caudal peduncle 15 (14-15); dorsal fin spinous rays 14 (14-15); dorsal fin branched rays 12 (11-13); total dorsal fin rays 26 (2627); anal fin spinous rays 3 (3); anal fin branched rays 9 (9-10); total anal fin rays 12 (12-13); abdominal vertebrae 15 (15); caudal vertebrae 14 (14); gill rakers (ceratobranchial) 11 (11-12). DESCRIPTION: Based on six adult and sub-adult specimens. Measurements and counts for the holotype and paratypes are given above. Figure 2 shows body form, fin positions and shapes and preserved colouration pattern. Due to the small number of specimens only a single specimen (RUSI 58260, paratype #4, 96.6mm) was dissected for the description of the jaws and teeth. Body. Compressed and deep with the greatest body depth at the origin of the pelvic fins. Predorsal length short. Head large and negatively allometric with SL. Preorbital depth, snout length and cheek depth positively allometric with SL. Eye large, situated dorsally and negatively allometric with SL. Caudal peduncle short and deep. Upper jaw (Fig. 3a & b). Premaxillary pedicel is slightly longer (103%, 12.5 mm in a specimen 96.6 mm) than the dentigerous arm of the premaxilla. The premaxillary pedicel slopes at an angle of 77 ° from the horizontal. The dental arcade comprises two rows of closely set unicuspid teeth. In the outer row there are (left side 27, right side 26) 53 teeth. These teeth alter in curvature and implantation with position being slightly recurved and erect at the anterior symphysis and changing to more recurved and recumbent posteriorly. The outer row teeth extend 81-83% along the dentigerous arm of the dentary. The inner row teeth are slightly smaller and closer set than those in the outer row and are all recumbent (Figure 3b). The inner row teeth comprise (left side 19, right side 21) 40 teeth and they extend 43% along the dentigerous arm of the dentary. Lower jaw (Fig. 3c & d). Relatively long with a rounded dental arcade. There are two rows of erect, slightly recurved and irregularly spaced unicuspid teeth. The outer row comprised 37 teeth (left side 19, right side 18) and extends 66.2-71.1% of the dentigerous arm of the dentary. The inner row teeth comprise a few anterior teeth (left side 6, right side 4) which extend 22.3-22.8% of the dentigerous arm of the dentary. Maxilla (Fig. 4a). The maxilla is stout (maximum length 12.1 mm) with its maximum width approximately 4/5 along the bone on the posterior flange (3.5 mm). Lower pharyngeal bone & teeth (Fig. 5a). The pharyngeal bone is triangular and not thickened. The dentigerous surface is 62% of the total length and 71% the total width of the bone. The ten posterior-median teeth are slightly enlarged while the remainder are slender and bevelled becoming more slender towards the edges of the bone. There are 10-11 teeth along the centre and 19 along the posterior edge of the bone. Gill rakers (Fig. 5b). Simple, relatively short gill rakers and number 11-12 on the ceratobranchial of the first gill arch. The three to four anterior rakers are reduced to small bumps of tissue. Total gill rakers are 15-16. Scales. Predominantly cycloid with a few weakly ctenoid scales on the lower flanks. Thoracic region fully covered with small, deeply embedded scales that reduce gradually from the larger flank scales. Scales are present on the head up to the interorbital area and cover the cheeks up to the lachrymal bone (usually 4-5 rows which are sometimes not arranged in defined rows). Scales between the orbit and preopercular groove 3-5. Approximately half of the caudal fin is scaled but there are no scales on the other fins. There are 6-7 scales between the pelvic and pectoral fin bases, 5-7 scales between the lateral line and the origin of the dorsal fin. The last two scales in the upper LL are separated from the dorsal fin base by one small and one large scale while anterior to these there are two or more large scales. Sensory structures. Simple finger-like projections (<1 mm) are present on the lower operculum, inner pelvic fin membranes and on skin on the ventral body directly above the pelvic area near the fin joint. These are possibly tactile sensory structures and indicative of contact with the substrate when foraging. Lateral line sensory canals on the head are branched forming clusters of tubes. Enhanced lateral line development may compensate for poor vision in turbid conditions. Fins. Positions and pigment patterns of fins in preserved specimens are shown in Figure 2. Pelvic fins just reach the origin of the anal fin. Caudal fin slightly emarginate with rounded edges. Colour in alcohol. Preserved material (Fig. 2) has a beige background colour that is pigmented brown by a series of spots and lines. The anterior head is pigmented with what appear to be fragmented nostril, interorbital, supraorbital and nape stripes i.e. paired blotches either side of the head centre line in these positions. Mental and opercular blotches are present. There are three poorly defined and broken lines along the body: mid-lateral, dorsal lateral and dorsal medial bands. The mid-lateral and dorsal medial bands both continue anteriorly up to the eye. There are 6-7 short and indistinct vertical bars on the body. The membranes of the fins are pigmented brown to varying degrees with the darkest section being the anterior dorsal fin. The membranes of the dorsal fin posterior (posterior of spine 9) and caudal fin anterior are covered with dark brown maculae. The centre of the soft rayed part of the anal fin has a series (up to nine) of eggs spots, not quite as large or as darkly pigmented as the caudal and dorsal maculae. The belly and ventral flank are unpigmented. Colour in life. Unknown. Habitat (Fig. 6). All specimens were caught near a submerged tree in a quiet backwater protected from the main river flow by a rocky outcrop. Distribution. Known only from the Lecitu River (Buzi system) at Dombe (19° 55’S 33° 22’E), Mozambique. Etymology. The genus name Chetia is feminine (Eschmeyer 1998). The species name brevicauda, Latin for short and tail respectively (Brown 1979), describes the low number of caudal vertebrae and the short caudal peduncle. Associated fish community. The Dombe pontoon site was rather poorly sampled and only the following species were collected with C. brevicauda: Distichodus mossambicus, Hydrocynus vittatus, Synodontis zambezensis and Glossogobius callidus. REMARKS Generic placement: Greenwood (1993) stated that there is no single diagnostic autapomorphy to distinguish members of the genus Chetia. Rather, there are a suite of characters which exclude them from other riverine haplochromines (Sargochromis Regan 1920, Serranochromis Regan 1920, Pharyngochromis (Steindachner 1866) and Thoracochromis Greenwood 1979). Greenwood considered Chetia to be closely related to the genus Serranochromis based on the following synapomorphic features: two or more postorbital scale rows (from Lippitsch 1990); high numbers of caudal vertebrae (15-17); and an increased number of circumpenduncular scales (16-20). Serranochromis apomorphies not shared with Chetia are: high numbers of abdominal (15-19) and total vertebrae (31-36); high lateral line scale counts (34-41); and unicuspid teeth in juveniles. Consequently, Greenwood suggested the rather unsatisfactory diagnosis for Chetia as “a Serranochromis lacking the apomorphic features of that genus”. The new species has three to five post-orbital scales which clearly places it within the Serranochromis-Chetia “group”but it does not exhibit the other two characteristic features which are high numbers of caudal vertebrae or circumpeduncular scales. The new species differs from Serranochromis in that it has a lower LL scale count and lower numbers of abdominal (and consequently total) vertebrae. The new species also has a smaller mouth compared to all other Serranochromis, a feature not referred to by Greenwood, but one which is used as a key distinguishing feature by Skelton (2001). The six specimens comprising the type series are all above 96mm SL. The presence of bicuspid teeth in juveniles, another Chetia characteristic, was therefore not verified. The form of anal egg spots was also considered an important feature by Greenwood (1993). Only preserved material was available at the time of description and, as will be discussed later, the condition of egg spots in live and preserved material changes considerably. The new species exhibits certain features which do not fall within Greenwood’s (1993) definition of Chetia but which do fit Sargochromis and Pharyngochromis characteristics. These features are: low circumpeduncular scales (14); low LL scales (31-32); low caudal (14) and total (29) vertebrae. Consequently, the inclusion of this new species in the genus Chetia, which we believe is correct, does increase the overlap between generic characters for the serranochromine genera (Table 2). Generic characteristics: With the inclusion of Chetia brevicauda sp. nov. the generic definition of the Chetia needs to be altered slightly. The lower limit of the caudal vertebral count is now 14 and the lower limit of the total vertebral count is 29. The range of scales in the lateral line series is now 31-35 and that of gill rakers is 9-12. Table 2 is a summary of the serranochromine genera (from Greenwood 1993) together with the altered characteristics for Chetia. Anal fin spots: The anal fin spot pattern recognised by Greenwood (1993: 34 & 36) for Pharyngochromis, Serranochromis, Sargochromis and Chetia was of small, numerous non-ocellated spots similar to the maculae on the dorsal and caudal fins. This was considered a key character distinguishing these genera from other riverine haplochromines which have ocellated spots i.e. a clear area surrounding the outside of the spot. Examination of live specimens of the type species C. flaviventris, collected in the Roodeplaat Dam near Pretoria, indicated that the pattern of the anal spots in these cichlids was slightly different than that described by Greenwood (1979: 274-275 and 1993: 34). Two specimens, collected and photographed at the same time are illustrated - an immature male and a mature male in breeding colouration (Figure 7). The immature male has ocellated eggs spots, i.e. an orange spot surrounded by a grey/dark circle and then by a clear region. The anal fin is pigmented yellow but is not opaque. In preservation these spots are grey, evenly pigmented and have no clear surround visible. The egg spots of the sexually mature male were yellow anteriorly becoming orange-centered, fading to yellow edges posteriorly and surrounded by a grey/dark ring. No clear area was visible and the whole anal fin was whitish and opaque. In preservation these spots are grey, some solid and some with a darker edges but none with clear surrounds, and the fin is white and opaque. Within this sample of C. flaviventris, RUSI 61095, 26: 55-148 mm, there was considerable variation in the numbers, colour and form (shape and ocellation) of the anal spots. In general, the mature males had larger and more numerous anal spots than immature males. Consequently, we suggest this character warrants a more thorough investigation before it is used for determining relationships within the serranochromine group. The only consistent difference noted in anal fin spot patterns during our superficial study of mostly museum serranochromine material is that between Serranochromis robustus jallae (Boulenger 1896) and all other Serranochromis (including the Lake Malawi form of S. robustus robustus). In all the specimens we have examined of S. robustus jallae the anal spots are confined to the posterior half of the anal fin while in other species eggs spots are distributed over the entire fin with increasing coverage with increasing SL (see Skelton 2001 for colour illustrations). This character may indicate that S. robustus from the upper Zambezi and Lake Malawi are separate species. Dentition: The teeth and their arrangement are typical of both Chetia and Serranochromis (Greenwood 1979: 300). Particularly characteristic is the antero-medial arc of inner teeth on the dentary that comprises less than 10 teeth (Fig. 3d). In other haplochromines, inner row teeth comprise greater numbers of teeth and tooth rows and these extend down the lateral part of the dentary. Slight differences were observed between C. brevicauda and C. flaviventris and S. robustus jallae. The arrangement of the dentary teeth was the same in the three species, but on the premaxilla of C. flaviventris there were three rows of teeth antero-medially reducing laterally to two almost entire rows and the posterior teeth were not as recumbent as in C. brevicauda. The maxilla of C. brevicauda was much larger and deeper than in either C. flaviventris or S. robustus jallae despite the specimens of latter species being slightly longer (Figure 4). Distribution: Members of the genus Chetia exhibit a disjunct distribution pattern (Table 3), similar to several other species in the region such as Mesobola brevianalis (Boulenger 1908) and Barbus bifrenatus Fowler 1935 (Skelton 1994). It adds further support to the widely held hypothesis of connections between the Zambian Congo, Kafue, Upper Zambezi, Kunene, Limpopo, Incomati and Buzi Rivers (Bell-Cross 1972, Gaigher & Pott 1973, Skelton 1994). However, the finer details of river connections remain to be determined and a phylogenetic analysis of these species and species-groups is now needed. Two possible routes for Chetia brevicauda or its ancestor to colonise the Buzi system are: 1) from the north (Zaire system) through the lower Zambezi or east coast rivers (together with many other “Zairean”species); or 2) from the upper Zambezi-Limpopo connection and then north. The limited distributional ranges of each of the five Chetia species is also noteworthy. Three of the species are poorly known, though, the late discovery of Chetia mola Balon & Stewart 1983 and C. brevicauda and the rarity of Chetia welwitschi (Boulenger 1898) seems to indicate they are naturally uncommon or have restricted distributions within their geographical ranges. The two species which are well known, C. flaviventris and C. brevis, also have restricted ranges, the Limpopo and Incomati systems respectively. Buzi River fauna: The discovery of C. brevicauda increases the status of the Buzi system as an area of high endemicity and special scientific (biogeographic) interest as already noted by Skelton (1984). Other endemics include the claroteid catfish A. hildae and the amphiliid catfish Amphilius laticaudatus Skelton (1984). Additional species which have slightly wider distributional ranges, being endemic to the Buzi and Pungwe systems, are Parakneria mossambica Jubb & Bell-Cross 1974 (Kneriidae), Varicorhinus pungweensis Jubb 1959 (Cyprinidae) and Barbus manicensis Pellegrin 1919 (Cyprinidae). New records since Bell-Cross’ 1973 survey have raised the number of species known from the Buzi system to 63. A new checklist is given in Table 4. Names follow Skelton (1993, 2001, 2002) and museum accession numbers are given for most new records collected by Weyl and Bills. Three exotics are present in the Buzi system. Two of these (Oreochromis niloticus (Linnaeus 1758) and Micropterus salmoides (LacepPde 1802)) were introduced into Zimbabwean sections of the Buzi and have now moved downstream into Mozambique (Weyl 1998). The third exotic (Cyprinus carpio Linnaeus 1758) was introduced into the Sussundenga Aquaculture station in Manica Province, Mozambique during 1996 (pers. obs., O.W.). Notable absences from the checklist are several species which occur in the lower Zambezi and northern South Africa such as Clarias ngamensis Castelnau 1861, Clarias theodorae Weber 1897, Ctenopoma multispine Peters 1844 and Microctenopoma intermedium (Pellegrin 1920). These are typical floodplain species and are possibly indicative of the fact that the floodplain reaches of the Buzi system are the most poorly known. We anticipate that these and several estuarine species known to enter freshwaters e.g. Elops machnata (ForsskDl 1775) and Acanthopagrus berda (ForsskDl 1775), will be found when thorough surveys are conducted in the lower reaches of the Buzi. ADDITIONAL MATERIAL EXAMINEDChetia flaviventris. Holotype: BMNH 1957.11.16.1, 104.1 mm, Buffelspoort Dam, Sterkstroom, tributary of Crocodile River, Limpopo system, South Africa, collected by Mr S.S. du Plessis. Paratypes: BMNH 1957.11.16.2-3, (2) 72.7-90.9 mm; RUSI 45386, (1) 118.2 mm. Other material: RUSI 28969, (2) 68.5-78.3 mm; RUSI 53629, (6) 47.5-94.5 mm; RUSI 61095, (26) 55-148 mm. Chetia brevis. Holotype. AMG/PF951, 120.5 mm, Lomati River, South Africa, collected I.G. Gaigher, September 1967. Paratypes. AMG/PF952, 73.8-89.3mm. Other material: RUSI 27639, (1) 70.2 mm; RUSI 63749 (5), 66.6-80.7 mm; RUSI 63750 (2), 43.2-59.1 mm. Chetia mola. Holotype. ROM 29825, 117.9 mm, Luongo River above the mouth of the Chambwishitu tributary, Zambia, collected by E.K. Balon, 17 November 1970. Paratypes. ROM 29826, (3) 74.6-132.0 mm. Chetia welwitschi. Holotype. BMNH 1864.7.13.62, 99.7 mm, Fluilla (Huila?), Angola. Thoracochromis buysi. RUSI 63330, (38) 46.8-137.3 mm. Thoracochromis albolabris. RUSI 59465, (6) 35.5-72.7 mm. Pharyngochromis acuticeps: RUSI 39049, (1) 94 mm. Serranochromis thumbergi: RUSI 62392, (1) 200 mm. ACKNOWLEDGEMENTSThe following curators/institutions are thanked for material loans: Royal Ontario Museum, Canada, Ann-Marie Hine, British Museum of Natural History, London, U.K.; Guy Teugels, Central African Museum, Tervuren, Belgium; Jim Cambray and Pat Black, Albany Museum, Grahamstown, South Africa. The following people are thanked for helping collect comparative material: Daksha Naran, Ben van der Waal, Andrew Deacon, Johann Engelbrecht and Kevin Roberts. The staff of the South African Institute for Aquatic Biodiversity are thanked for their assistance in many aspects of this work. In particular, discussions with Paul Skelton on cichlid taxonomy, Sally Terry aided with much of the laboratory work and made all X-rays and Elaine Heemstra drew the figures. Denis Tweddle and Ofer Gon made valuable comments on this work and Jos Snoeks is thanked for improvements made to the final draft. LITERATURE CITED
Copyright 2002 - The South African Institute for Aquatic Biodiversity, Grahamstown, South Africa. Free, full-text also available from http://www.saiab.ru.ac.za/smithiana/index.htm The following images related to this document are available:Photo images[sp02001t1.jpg] [sp02001f7.jpg] [sp02001t2.jpg] [sp02001f5a.jpg] [sp02001t4.jpg] [sp02001f1.jpg] [sp02001f3.jpg] [sp02001t3.jpg] [sp02001f5b.jpg] [sp02001f2.jpg] [sp02001f4.jpg] [sp02001f6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}