 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
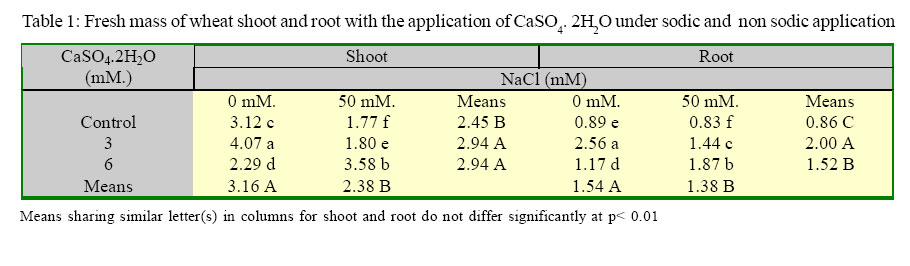
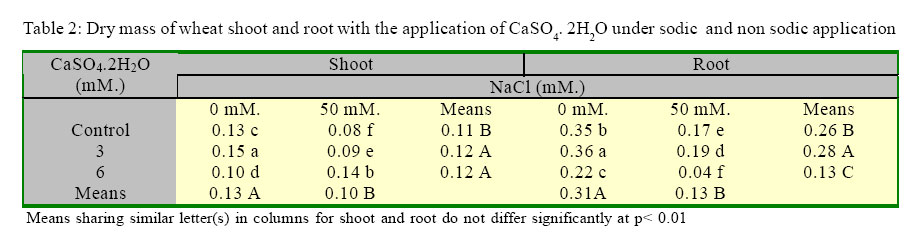
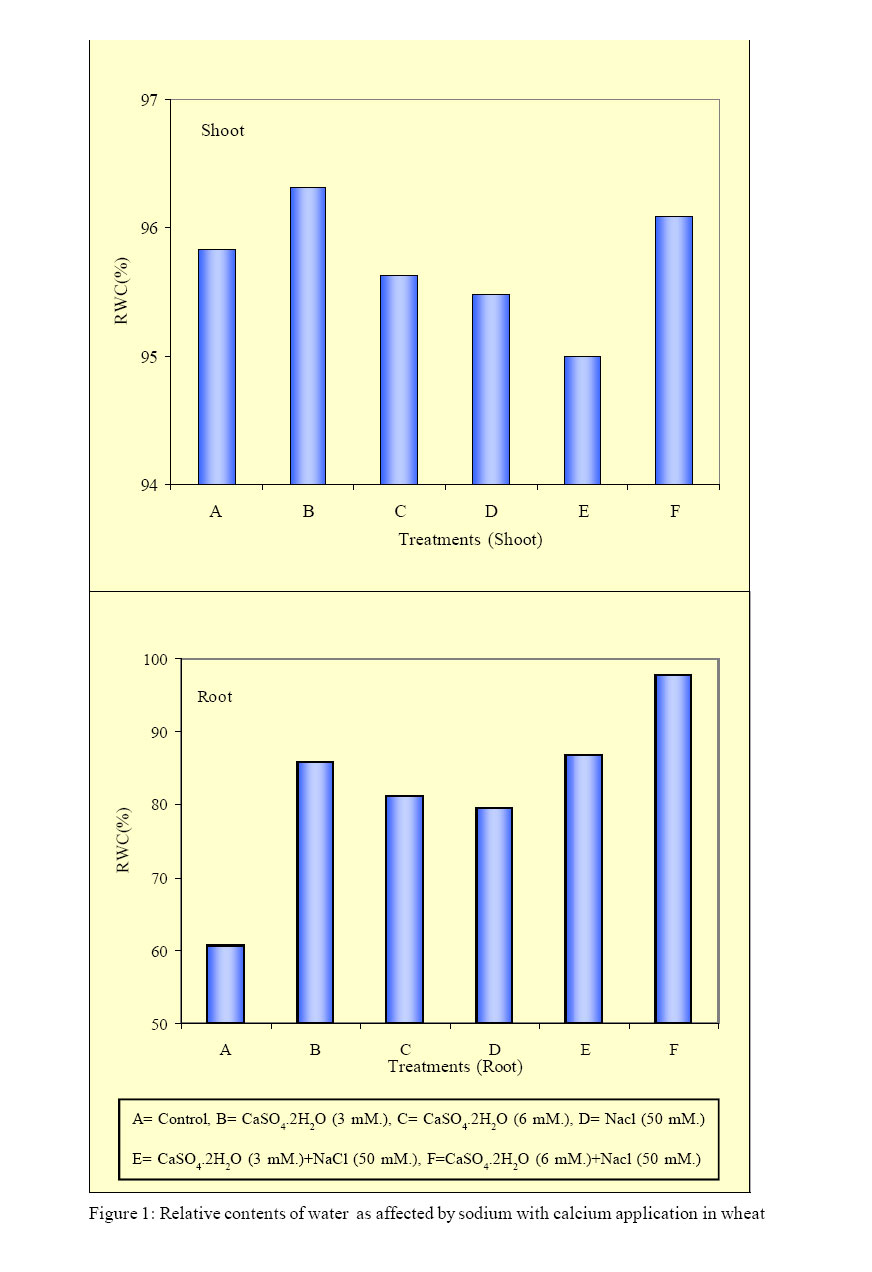
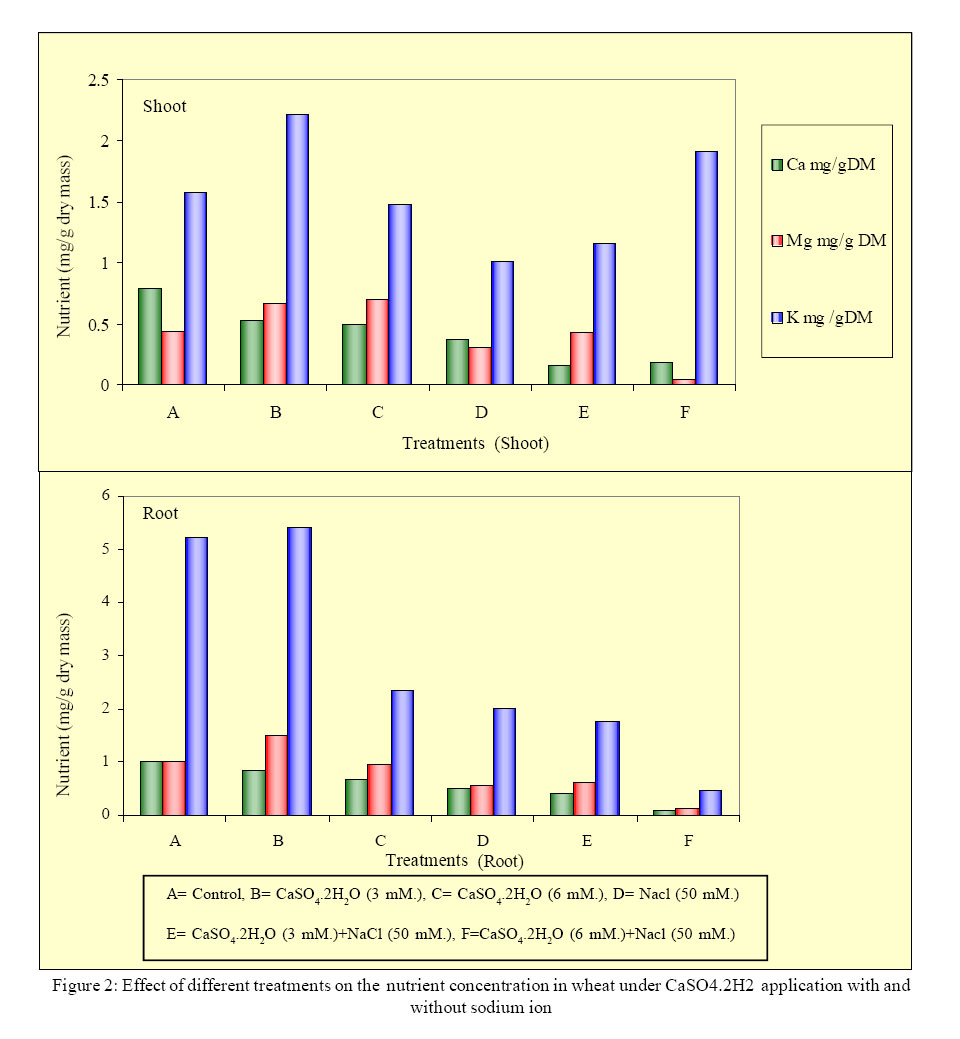
International Journal of Enviornmental Science and Technology, Vol. 2, No. 1, Spring, 2005, pp. 7-12 Response of wheat plants to sodium and calcium ion interaction under saline environment 1B. Zaman, 1B. H. Niazi, 2*M. Athar and 1M. Ahmad 1Institute of Natural Resources and Environmental Sciences, National Agricultural Research Center, Pakistan Agricultural Research Council, Islamabad-45500, Pakistan Code Number: st05002 Abstract Wheat being a glycophyte crop, responds differently to saline-sodic soil environmental conditions. The application of calcium is multidimensional with respect to sodium ion and plant part response. This study was conducted to record the response of shoot and root to sodium and calcium interaction under saline environment. Wheat seed of variety Punjab 85 were raised in quartz sand. Later on the seedlings were transplanted to pots containing Hoagland’s nutrient solution along with NaCl at 0 mM. and 50 mM. Calcium was applied as CaSO4 2H2O at 3 mM. and 6 mM. Under saline conditions shoot showed positive response to sodium ion in the presence of higher calcium. Relative water contents were higher in the root system at 6 mM of CaSO4. 2H2O under saline condition. Growth responses to potassium and Magnesium in the presence of sodium induced salinity with calcium ion interaction remained variable. Key words: Wheat, sodium/calcium ion, interaction, salinity Introduction Wheat being a glycophyte is sensitive to tolerant under saline and saline-sodic soil environmental conditions. Rapid change in climatic conditions and deteriorating water quality are due to anthropoid behavior. This results in the selection of such cultivars of this staple food crop that can not face adverse properties of salinity and sodicity. This may include specific ion toxicity, ion imbalance, water stress and osmotic non-adjustment. Soil salinity is a global problem and results in reeduction in plant growth, reduced yield and, in severe cases, total crop failure(Qadir, et al., 2000). Of all the ions in a saline or saline-sodicmedium Na+ may producedetrimental effects on the availability of water in a root medium as a function of growth period and concentration in the root medium. Source of sodium ion also plays an important role in plant response to salinity. Sodium sulphate is less toxic than sodium chloride for the growth of wheat provided potassium and calcium are available in the growth medium (Zaman, et al., 2002a). Elevated level of calcium regulates and maintains potassium concentration in plants (Lauchli, 1990). Therefore the availability of macronutrients in saline or saline-sodic medium is beneficial for the yield of crops. Zaman, et al., (2002b) observed that sodium ion and dry mass are negatively correlated and application of sulphur as sulphate improves nutrient to sodium ion ratios. Salinity stress changes water permeability of the cell membrane. At high salinity level water permeability of coleoptile subepidermal cells is decreased (Mansour and Stadelmann, 1994). No visual symptoms of wilting may appear though water permeability may significantly be decreased by salinity stress. The primary effect may take place in roots due to water deficiency (Flowers and Hajibagheri, 2001). Water stress decreases water potential of tissue (Yordanov, et al., 2003). Siddique, et al. (2000) studied the effects of drought stress on water relations of wheat and observed decrease in water level under stress conditions. Physiological observations associated with the varietals difference in tolerance have been reported (Isla, et al., 1998). Keeping in view the effect of salinity on crop production, a study was conducted to observe the response of wheat plants to sodium and calcium ion interaction under saline conditions. Materials and Methods A hydroponic study was conducted under controlled conditions. Seeds of wheat (Triticum aestivum L. cv. Punjab 85) were germinated in quartz sand irrigating with distilled water. Ten days old seedlings were foam-plugged in lids of plastic pots containing 2.5L of Hoagland’s nutrient solution keeping the pots continuously aerated. The light intensity was 450 µmol m-2 s-1. Photoperiod was adjusted to 16 h. light period and temperature was maintained at 30 ± 2 °C. The pH of the solution was adjusted to 6.0 with HCl or KOH and was regularly monitored. The salinity level was 50 mM. as NaCl, while 0 mM. NaCl served as control. Calcium as CaSO4. 2H2O was applied at 3 and 6 mM. to the pots. The salinity level (25 mM. ) was imposed 10 days after transplantation by incremental addition of NaCl at 5 mM. NaCl per day. The pots were arranged in complete randomized design in triplicate. The plants were harvested 44 days after sowing and their fresh weight recorded. The plants were rinsed with deionised water and separated into root and shoot portions. Dry weight of each sample was recorded after drying at 65°C to constant mass. The roots and shoots were ground to pass a 40-mesh Wiley Mill and digested in 2:1 perchloric-nitric di-acid mixture. Sodium, potassium, calcium and magnesium ions in the digested material were determined by atomic absorption spectroscopy. The data were statistically analyzed according to CRD and treatment means were compared using LSD test. Results Growth parameters of the plants had a highly significant (P<0.01) effect of the treatments applied (Table 1). In the shoot, fresh weight increased by 30 % with the application of 3 mM. CaSO4. 2H2O. Fresh weight decreased by 27 % with 6 mM. CaSO4.2H2O in treatment without salinity. In the presence of 50 mM. NaCl, fresh weight decreased by 2% with the application of 3 mM. CaSO4. 2H2O and it increased by 102 % with 6 mM. CaSO4. 2H2O. In root it increased by 188% and 31% with application of 3 mM. and 6 mM. CaSO4.2H2O respectively, without salinity. In the presence of 50 mM. NaCl, fresh weight increased by 73 and 125% with application of 3 mM. and 6 mM. CaSO4.2H2O respectively. In the shoot, dry matter (DM) increased by 15% with the application of 3mM. CaSO4.2H2O, and it decreased by 23% with 6 mM. CaSO4.2H2O applied in the plants without salinity (Table 2). In the presence of 50 mM. NaCl, DM increased by 13 and 75% with the application of 3 mM. CaSO4.2H2O and 6 mM. CaSO4.2H2O respectively. In the root, DM increased by 3% with 3 mM. CaSO4.2H2O and decreased by 37% with 6 mM. CaSO4.2H2O without salinity. In the presence of 50 mM. NaCl, DM increased by 12% with 3 mM. CaSO4.2H2O and decreased by 76% with 6 mM. CaSO4.2H2O. As per ascending order, in shoot, the sequence of relative water content (RWC) as given in Figure 1 given as: 3 mM. CaSO4.2H2O with salinity, 3 mM. CaSO4.2H2O without salinity, 6 mM. CaSO4.2H2O with out salinity, salinity, control. Relative water content in root in the same sequence was: control, salinitz, 6 mM. CaSO4.2H2O, 3 mM. CaSO4.2H2O, 3 mM. CaSO4.2H2O with salinity, 6 mM. CaSO4.2H2O with salinity. A comparison between shoot and root showed that at 6 mM CaSO4. 2H2O applied in the presence of salinity RWC was higher in the root system. Expansion of growth was higher in root than shoot at 6 mM CaSO4. 2H2O in the presence of salinity. In shoot, in the absence of salinity with the application of 3 mM. CaSO4.2H2O potassium concentration (2.20 DM) was the highest (Figure 2) and its concentration was lowest (1.02 mg/g DM) in the presence of salinity. In root, in the absence of salinity with the application of 3 mM. CaSO4.2H2O, potassium concentration (5.40 mg/gDM) was the highest and on the induction of salinity alone its concentration was lowest (4.76 mg/g DM). In the absence of salinity in shoot, calcium concentration (7.93 mg/g DM) was the highest (Figure 2) and in the presence of salinity its concentration was lowest (1.53 mg/g DM) on the application of 3 mM. CaSO4.2H2O. In root, in the absence of salinity, calcium concentration (1.02 mg/g DM) was the highest and on the application of salinity alone its concentration was lowest (0.10 mg/g DM). In shoot, in the absence of salinity magnesium concentration (7.70 mg/g DM) was highest and its concentration was lowest (0.05 mg/g DM) on the application of 6 mM. CaSO4.2H2O in the presence of salinity. In root, in the absence of salinity and with the application of 3 mM. CaSO4.2H2O, calcium concentration (1.51 mg/g DM) was the highest and its concentration was lowest (1.40 mg/g DM) on the application of 6 mM. CaSO4. 2H2O in the presence of salinity. Discussion and Conclusion Physiologically, growth is a function of different kinds and levels of salts applied in the root medium. The excess of Ca ion in the medium causes retardation in the growth of shoot whereas in the saline medium when sodium ion is available, the abundance of Ca ion is required to check the toxic activities of Na ion. Ebert, et al. (2002) pointed out that cation relations in the shoot tissue such as Ca+2/ Na+ have possessed a strong influence on salt tolerance than absolute sodium levels. Presence of higher level of Ca+2 induced signals and the concentration difference across the plasma membrane results in a very steep electrochemical gradient in favor of Ca+2 influx. Availability of Ca+2 in the external environment ameliorate NaCl stress (Zhong and Lauchli, 1994). Therefore, amount of available Ca+2 is also important. Higher level of calcium application (6 mM. CaSO4.2H2O) contributed towards salinity resistance. The contribution of such salts may not be toxic but play some important role for checking the water potential of plant which ultimately results in reduction of metabolites for the synthesis of biomass. A comparison between shoot and root showed that at 6 mM. CaSO4.2H2O applied in the presence of salinity, RWC was higher in the root system. Expansion of growth was higher in root than shoot at 6 mM. CaSO4.2H2O in the presence of salinity. This indicates that the presence of excessive amount of Ca ions contribute to decrease the toxic effects of salinity by enhancing tissue growth. The leaf water potential evaluates the water-stress intensity sensed by leaves (Hsiao, 1973). An important factor limiting the potential tolerance of all wheat varieties may be the high unidirectional influx of Na+ into the root, which is very similar in both tolerant and sensitive varieties (Davis, 1984 and Davenport, et al., 1997). Salt tolerance has been associated with preferential accumulation of K+ against high Na+ (Badr, et al., 2002a). Selectivity of K+/Na+ is improved by the presence of Ca+2 (Cramer, et al., 1985). Preference of K+/Na+ may be due to genetic factor (Greenway and Munns, 1980). The results show that organ preference for K+ over Na+ is also dependent on Ca+2 in the growth medium. Minhas, (1996) suggested that adverse effects of high sodium on the physical and chemical properties of soils can be mitigated by the use of different amendments, which contain soluble calcium e.g., gypsum. It is also suggested by Leopold and Willing (1984) that application of external calcium has been known to ameliorate salinity stress symptoms in many plant species. Lynch and Lauchi (1985) pointed out that Na+ concentration in root medium may induce calcium deficiency in plants grown with low calcium. Kinraide (1998) reported that main effect of Ca+2 on the Na+ uptake at high Ca+2 concentration was the electrostatic displacement of Na+ from plasma membrane surface. Maathuis and Amtmaan (1999) concluded that direct effect of Ca+2 on the movement of Na+ is manifested in voltage independent channels. Positive correlations were also reported for species tolerant to stresses that have a dehydrative component such as salt stress (Galvez, et al., 1993; Moons, et al., 1995). In the presence of salinity with the application of Ca+2 ion, the nature of salinity and the intensity changes that may cause a change to RWC as well as nutrient status in aerial and non aerial wheat plant parts. In wheat at 50 mM. of sodium induced salinity, the application of 6 mM. CaSO4.2H2O reduces the osmotic effects. References
© 2005 Center for Environment and Energy Research and Studies (CEERS) The following images related to this document are available:Photo images[st05002f2.jpg] [st05002t2.jpg] [st05002t1.jpg] [st05002f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}