 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
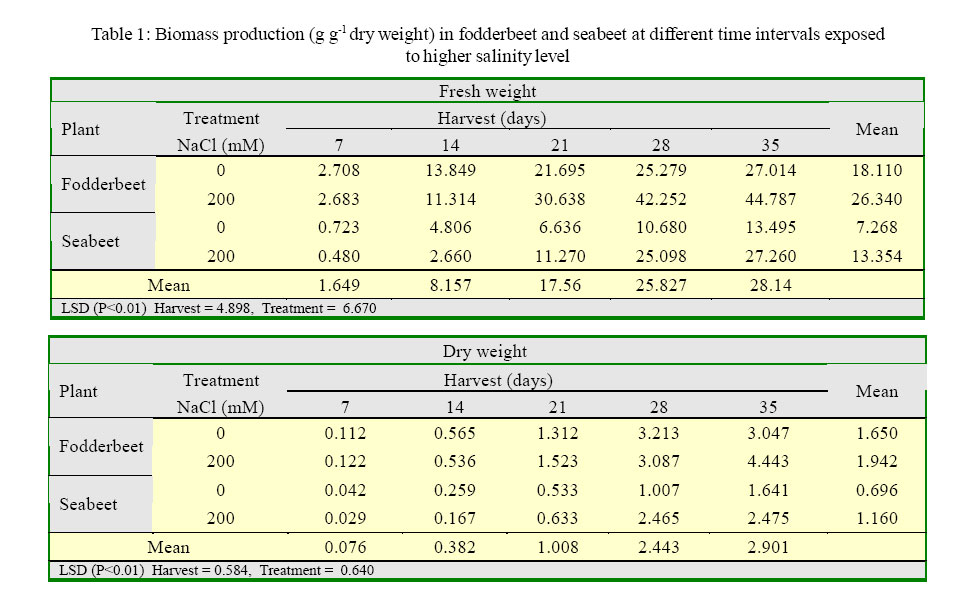
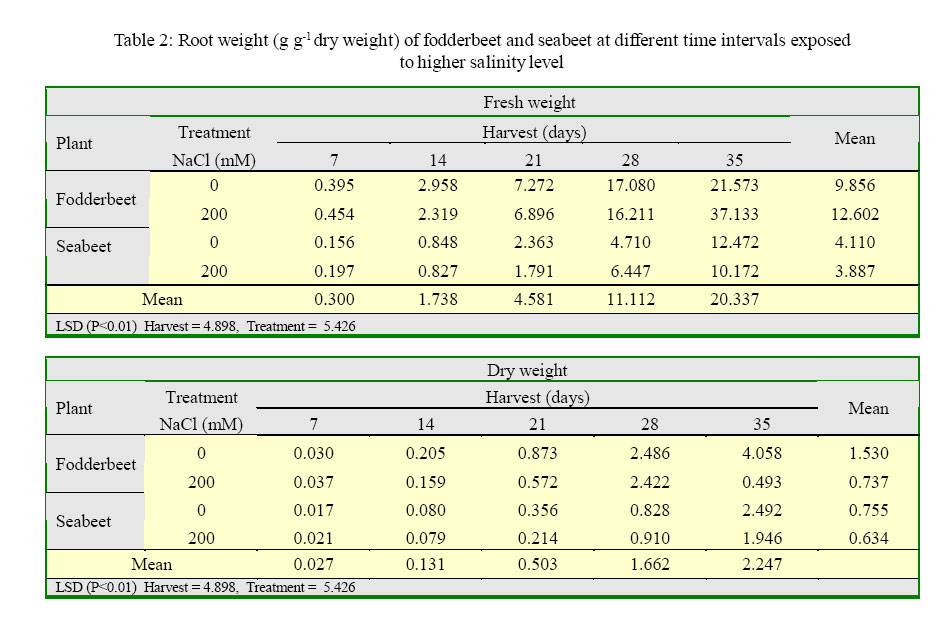
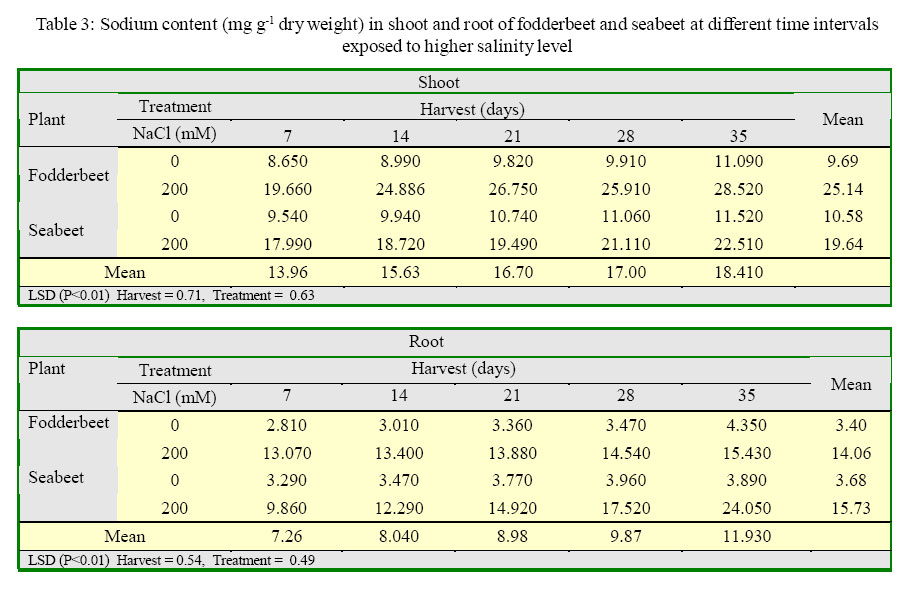
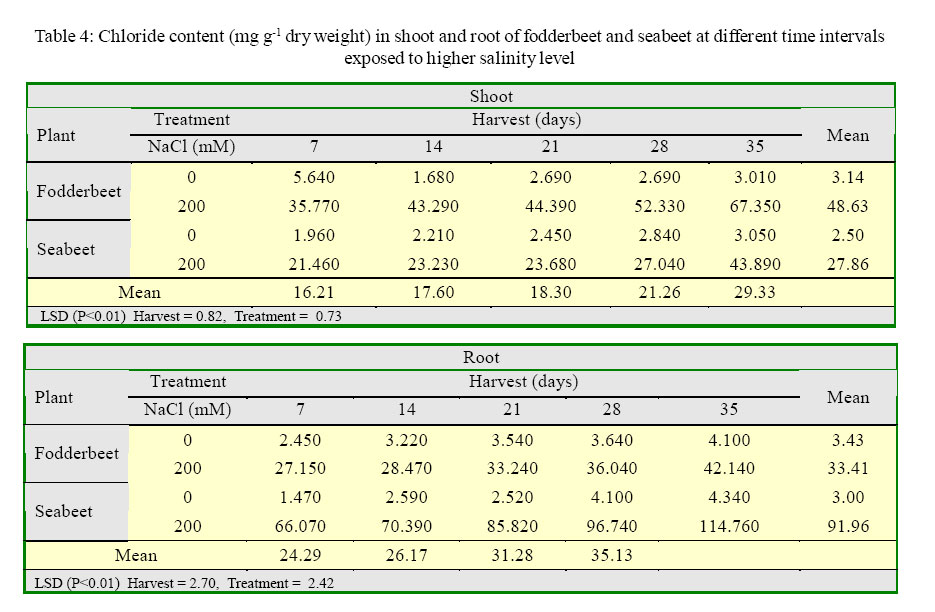
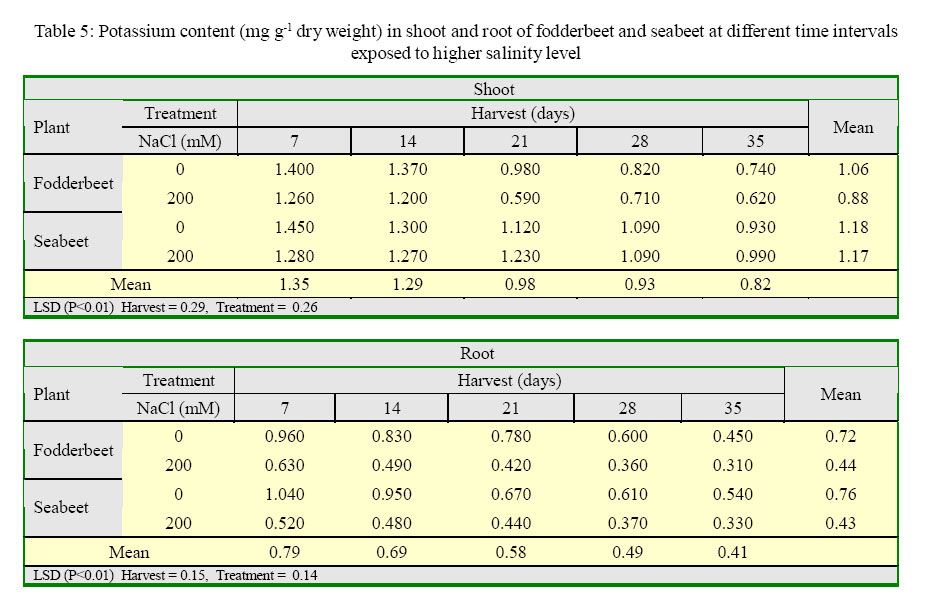
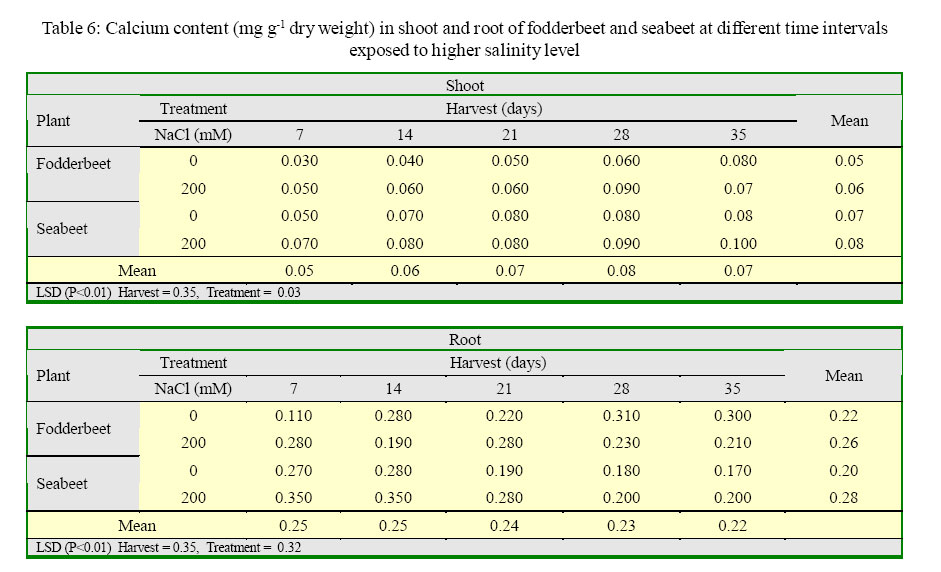
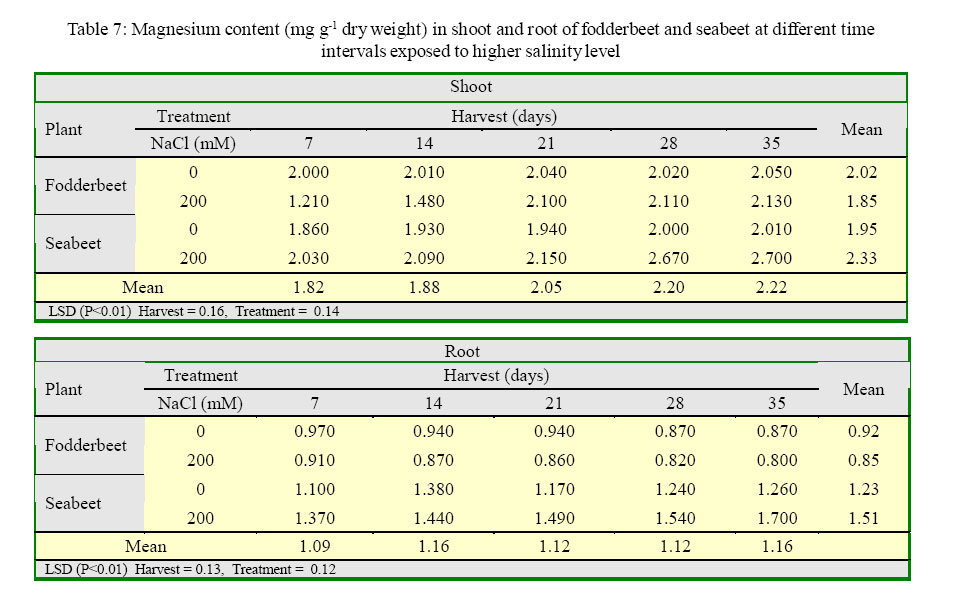
International Journal of Enviornmental Science and Technology, Vol. 2, No. 2, Summer, 2005, pp. 113-120 Growth and ionic relations of fodderbeet and seabeet under saline environments 1B. H. Niazi, 2*M. Athar, 1M. Salim and 3J. Rozema 1Institute of Natural Resources and Environmental Sciences, National Agricultural Research Center, Pakistan Agricultural Research Council, Islamabad, Pakistan Code Number: st05015 Abstract Growth and ionic relations of fodderbeet (Beta vulgaris) and seabeet (B.maritima) were studied in a greenhouse experiment using garden soil salinized with 200 mM NaCl. Both the species tolerated salinity level of 200 mM NaCl, but seabeet performed better than fodderbeet. Fresh weight of shoot increased significantly (P< 0.01) under higher salt concentration in both the species. Fodderbeet accumulated more biomass under saline conditions than seabeet. Salt treated plants accumulated significantly higher Na+ and Cl- content in shoot compared to untreated control plants. Chloride (Cl-) of seabeet root was significantly higher than shoot; however, chloride content of shoot was significantly lower in fodderbeet. The concentration of these ions was comparatively higher in seabeet root than fodderbeet in the later growth period. Potassium K+ content was low and revealed antagonistic effect with that of Na+ content. Calcium (Ca+2) ions were not significantly absorbed by shoot of both species. A significant effect of treatment by the root has been observed in both species. Magnesium (Mg+2) content of shoot were increased with the time. Seabeet has absorbed more Mg+2 than fodderbeet. The fodderbeet showed a significant salt tolerance during five week growth period. Seabeet is comparatively more salt tolerant. Domesticated cultivars of fodderbeet like Majoral have considerable adaptability in the inland salt affected areas of Pakistan. Key words: Sodium chloride, potassium, relative growth rate, fodderbeet, sugarbeet, saline environment Introduction Salinity is a limiting factor to crop production. Yields of most crops are decreased when cultivated in salt affected areas. Generally salinity problems are handled by chemical and biological methods. Chemical methods are usually used to reclaim sodic soils (Qadir, et al., 2000). While cultivation of salt tolerant species on salt-affected lands forms the basis of the biological reclamation. Identification of a wide variety of species with higher salt tolerance is important to achieve more success from this approach. Fodderbeet, is highly salt tolerant during vegetative growth (Niazi, et al., 2000). The sugarbeet and fodderbeet yield is improved by application of sodium chloride as fertilizer (Draycott and Durrant, 1976; Magat and Goh, 1988). This improvement has been related to the increase in root sugar yield with enhanced uptake of Na+ from soil (Hamid and Talibudeen, 1976). Chloride is a essential nutrient for plants (Marschner, 1986). Cell wall extensibility is significantly affected by deposition of Ca2+ in the cell wall, either by forming cross bridges or by inhibiting wall loosening enzymes (Cleland, 1986). The concentration of Ca2+ and Mg2+ in reclaimed soils was reported to be higher at the top (Qadir, et al., 2000 and 2001). Maximum Ca2+ uptake by the plant from these soils can affect the plant growth. Magnesium is an important component of chlorophyll (Marschner, 1986) and presence of magnesium in plant can indirectly improve the growth of plant by more chlorophyll synthesis and better rate of photosynthesis in plant. Although the halophilous nature has been reported in literature (Magat and Goh, 1988), there is a little information about the role of various ions in the growth of fodderbeet. These factors point towards the importance of various ions for the optimum growth of the plant tissue. Fodderbeet and seabeet are the chenopod mainly grown as supplementary stock feed in coastal areas in winter when there is an acute shortage of fodder for the livestock in Pakistan. A series of experiments are underway in our unit to explore the possibilities of improving growth and production of these plants under moderately salinesodic soil conditions (Niazi, et al., 1999, 2000, 2002, 2004a and 2004b). The present study constitutes the results of an experiment conducted to determine the growth response and ionic relations of the fodderbeet and seabeet grown in salinized garden soil in the greenhouse. The resaerch was conducted at National Agricultural Research Center, PARC, Islamabad, Pakistan during 2003. Materials and Methods Seeds of two species of fodderbeet (Beta vulgaris cv. Majoral) and seabeet (Beta maritima) were germinated in sand under greenhouse conditions (temperature: 30, EC: 25 in day/ night, relative humidity 70%, light: 250 mE m-2s-1). Ten days old seedlings were transplanted to pots (one seedling per pot) kept in greenhouse filled with one kg garden soil (Potgrond mengselm, Aalsmeer). Physico-chemical characteristics of the soil are provided elsewhere (Niazi, et al., 2004a). Two treatment levels (0 mM NaCl; (control), and 200 mM NaCl) were maintained in the pots. Salinity was increased stepwise by addition of 50 mM NaCl every two days (twice added on the top of soil and later, to plate placed at the base of pot to keep salinity uniform throughout the soil profile) one week after seedling establishment. There were 20 replications per salinity level. First harvest was done after seven days of maintaining the salinity level in the pots and the subsequent harvests were done with an interval of seven days between each harvest. Plant material was dried at 60 oC in an oven to a constant weight and was analyzed for cations (Na, K, Ca, Mg) using Atomic Absorption Spectrophotometer (Perkin Elmer 2100, USA). Chloride content was measured by Chloro-counter by the method instructed in the instrument operating manual (Marius Instrumenten, Utrecht, The Netherlands). The data were statistically analyzed using three way ANOVA methods described by Sokal and Rohlf (1981). Means were compared with the LSD multiple mean comparison test at significance levels at P< 0.01. Results Biomass production Fresh weight of shoot increased significantly (P< 0.01) under higher salt concentration in both the species. Fodderbeet accumulated more biomass under saline conditions than seabeet. Dry matter accumulation of the shoot increased with the age of the plant and the presence of salt enhanced the dry matter production (Table 1). Fresh weight of root did not increase significantly (P<0.01) for the first 21 days. It increased significantly in the later period under study. There was a significant (P<0.01) difference in the fresh weight of species. Salt treatment has no effect on fresh weight. Dry matter yield of root significantly decreased in fodderbeet under saline conditions. However, no significant effect on the dry matter yield of root was observed under higher salt treatment (Table 2). Ion concentration The concentration of Na+ in plants significantly (P< 0.01) increased in the presence of 200 mM NaCl in the root medium. The concentration of Na+ increased with time in control plants (0 mM NaCl) as well. In general, shoot had higher Na content than root (Table 3). Na+ content in root increased with time irrespective of treatment. Fodderbeet had absorbed higher Na+ than seabeet. Effect of high NaCl concentation in growth medium resulted in significantly (P< 0.01) higher Cl-in seabeet root which had higher Cl-than shoot (Table 4). Chloride content in both the shoot and root increased with time. Potassium content of plant generally decreased under saline conditions. Shoot absorbed significantly (P< 0.01) higher K+ content than root (Table 5). Potassium ion concentration decreased with time both in shoot and root. Uptake of K+ by root and shoot was significantly (P< 0.01) low under saline conditions in seabeet than fodderbeet. Plant growth showed a significant correlation (r = -0.72) with K ion uptake by the plant which is a useful trend in the plants grown under saline environments. Uptake of calcium was enhanced by the presence of salt (200 mM NaCl) during early growth period in root (Table 6). Calcium ion (Ca+2) content was significantly (P< 0.01) higher in seabeet root than fodderbeet during first week of growth. Calcium content significantly increased in fodderbeet shoot with the age of the plant. There was significant (P< 0.01) difference in Ca+2 content of seabeet shoot under saline conditions, at the age of 28-35 days. Calcium ion (Ca+2) content of seabeet shoot were increased in the control plants. Magnesium content in fodderbeet shoot decreased under saline conditions. However, Mg+2 content significantly decreased in fodderbeet root and shoot during 28-35 days in the presence of 200 mM NaCl (Table 7). Magnesium content in seabeet shoot Discussion and Conclusion The presence of NaCl in growth medium had enhanced the uptake of Na+ by the plant. Control (0 mM NaCl) plants also absorbed available sodium. The concentration of Na+ seemed not very high with plant growth due to rapid division and growth of cells and distribution of Na+ ions within the cells which caused dilution of ion concentration (Grieve and Fujiyama, 1987). Plant absorbed more Na+ under saline conditions from the growth medium than control, thus Na+ concentration of the plant tissue increased during growth until final harvest. Translocation of ions from root to shoot in the presence of saline environment was comparatively higher than the control. Uptake of water by the root caused a regular addition of Na+ from growth medium. Sodium ion concentration of root was probably low to interact with potassium absorption for the first two weeks. A significantly different Na+ accumulation in rice shoot under saline conditions have been described by Flowers and Yeo (1981). Grieve and Fujiyama (1987) noted a 35-40% standard deviation in data related to Na+ concentration in rice cultivars. This variation was related to shoot growth rather than a reduction of net sodium uptake. HighNa+ and Cl-ion accumulation by Beta spp. confirm the family Chenopodiaceae as salt accumulators (Rozema, et al., 1981). Presence of higher NaCl treatments increased Clion accumulation in the root upto the third week. The chloride ion further increased in seabeet until final harvest in the presence of high NaCl concentration. Addition of NaCl to growth medium is followed by increase in pH (probably due to Cl-). Chloride concentration increases in shoot with high pH, while root is little affected (Findenegg, et al., 1989). This condition is hazardous for plant growing under saline soil because of high pH, which will favour Cl concentration in plant and cause reduction in growth. Increase of Cl in shoot is favored by significant transportation of Cl to shoot. A highly significant correlation (fodderbeet r=0.93; seabeet r =0.97) between Na+ and Cl-ions has been recorded in shoot and root of both the species studied. Salinity induced growth reduction in seabeet was comparatively lower (Niazi et al., 1999) which is an indication of its high salt tolerance which may be related to low chloride concentration in shoot of seabeet compared to that of fodderbeet. The concentration of K+ in seabeet root significantly (P< 0.01) decreased under saline conditions. This may be related to antagonistic effect of K+ with Na+ seabeet has comparatively lower Na+ content in shoot than fodderbeet and K+ increase in shoot may be due to antagonistic effect with sodium under saline conditions (Plaut and Grieve, 1988). significantly (P< 0.01) increased after 7-35 days in the presence of 200 mM NaCl. A negative correlation (r=-0.77) has been observed in Na shoot/K root in fodderbeet while the seabeet has shown a higher correlation value (r=-0.88). Niazi and Ahmad (1984) in tomato and Ashraf, et al. (2003) in okra have also confirmed antagonistic effect of Na+ and K+ ion during a study on the uptake of ions in under saline soil conditions. Some graminaceous species growing in nutrient cultures containing NaCl as a sole osmoticum, frequently displayed leaf blade deformation and necrosis that are characteristics of severe Cadeficiency. The physiological role of Ca+2 in the cellular system is to stabilize the cell wall structure to maintain membrane integrity, selectivity and to regulate selectivity of ion transport process (Hanson 1983). Membrane permeability may increase by Na+ displaced Ca+2 ions that are associated with the external surface of plasma lemma (Cramer, et al., 1985). In the present study a significant increase in Ca+2 concentration of root was observed in the presence of NaCl, which may form part of a highly salt tolerant behavior of plant. Magnesium (Mg+2) concentration was also not reduced with time in fodderbeet, and even in seabeet an increase in Mg+2 was observed at 200 mM NaCl. The correlation of Na root/Mg shoot was significantly negative (r = -0.77) in fodderbeet, while this same correlation was highly positive (r=0.91) in seabeet. Increase of Ca+2in the leaf resulted in a marked reduction in leaf Mg+ 2. This Ca-Mg antagonism could lead to a disturbance in photosynthesis. Reduced photosynthesis was noted in maize due to Mg+2 deficiency (Peaslee and Moos, 1966). High Ca+2 in leaf may also interfere with CO2 fixation by inhibition of stroma enzymes specially those that are Mg activated (Charles and Halliwell, 1980). The present study proved no disturbance in Ca-Mg imbalance and Mg tended to increase with simultaneous increase in Ca+2 concentration. This resulted in better net photosynthesis of fodderbeet in the presence of high NaCl (Niazi, et al., 1999, 2004a and 2004b). The fodderbeet cv. Majoral tested showed a significant salt tolerance during first four week growth period under saline soil conditions. Salt tolerance is not a function of single organ or plant attribute, but it is the product of all the plant attributes (Ali, et al., 2004). Therefore a species exhibiting relative salt tolerance for all the plant attributes may be ideal one. Fodderbeet is halophilous in nature. Seabeet is especially highly tolerant because it grows in the coastal marshy places under natural conditions. The domesticated cultivarsof fodderbeet including cv. Majoral have been tested for adaptation in the inland areas of Pakistan (Niazi, et al., 1999). The better mechanism of salt tolerance may have been achieved by the plant through accumulation of more ions. The salt affected areas could be utilized by cultivation of these species to enhance the production of fodder for livestock as well as utilization of unproductive soils in the resource poor countries like Pakistan. Acknowledgments The financial assistance provided to first author by NUFFIC, The Netherlands, to complete this study is gratefully acknowledged. The first author is also indebted to the staff of the Department of Ecology and Ecotoxicology, Free University, Amsterdam, The Netherlands for technical help and cooperation during this study. The authors are thankful to Dr. Shoaib Ismail, International Center for Biosaline Agriculture, Dubai, UAE for valuable suggestions and helpful criticism on the manuscript. Appreciations are also extended to an anonymous reviewer at University of California, Davis, USA for critical comments that undoubtedly improved the quality of the paper. Authors are also grateful to Dr. Manzoor Qadir, Institut für Pflazenernährung, Giessen, Germany for providing many original reprints that helped in the preparation of this paper. References
© 2005 Center for Environment and Energy Research and Studies (CEERS) The following images related to this document are available:Photo images[st05015t6.jpg] [st05015t3.jpg] [st05015t2.jpg] [st05015t5.jpg] [st05015t7.jpg] [st05015t4.jpg] [st05015t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}