 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
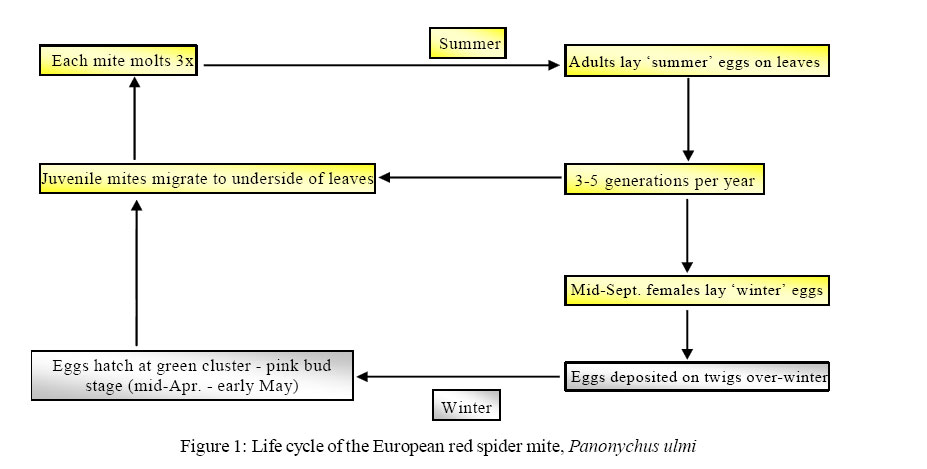
International Journal of Enviornmental Science and Technology, Vol. 2, No. 3, Autumn, 2005, pp. 287-290 Review Paper European red spider mite -an environmental consequence of persistent chemical pesticide application 1*A. G. S. Cuthbertson and 2A. K. Murchie 1Central Science Laboratory, Sand Hutton, York YO41 1LZ, UK Received 18 February 2005; revised 23 June 2005; accepted 22 August 2005; onlined 30 September 2005 Code Number: st05039 The European red spider mite, Panonychus ulmi, is a major pest in almost all fruit growing regions of the world (Hardman et al., 1985). Spread of P. ulmi to most apple-growing areas has probably been caused by the distribution of nursery stock carrying winter eggs. This mite is stated to be an important secondary pest (due to the effects of chemical sprays killing natural enemies) of commercial orchards throughout the United Kingdom (UK) and Europe (Cross andBerrie, 1994).Awide host range includes deciduous bushes and trees belonging to the family Rosaceae, but it is in association with fruit trees such as apple, pear, plum, peach, prune and cherry that it reaches economic importance. In commercial orchards, the potential of P. ulmi to cause severe economic damage necessitates chemical control several times a year (Croft, 1975). Adult females are about 0.4 mm long; body oval, strongly convex and dark red, with long setae arising from light-coloured pinacula (Alford, 1984). The male is similar to the female only smaller. Eggs are orange/ red and about 0.15 mm long. These hatch into six legged larvae with an average length of 0.17 mm (Blair and Groves, 1952). Panonychus ulmi over-winter as diapausing eggs laid on the bark of trees or smaller branches and spurs (Figure 1). During a heavy infestation, areas of the bark may even appear red due to the presence of many eggs. In UK Bramley orchards, and in particular Northern Irish orchards, eggs hatch from mid-April to the end of June depending on climatic conditions (Cuthbertson, 2000). On hatching, juvenile mites move to the underside of the leaves and begin feeding. They reach maturity in approximately three weeks after undergoing three moults. The number of generations per year varies between geographical regions, with for example, in the UK up to five being usual for the south of England (Blair and Groves, 1952) but in the cooler climate of Northern Ireland 2-3 generations is more common (Cuthbertson, 2000). Panonychus ulmi lays two types of eggs. Summer eggs laid on the leaves of the host plant are of the non-diapause type and develop without interruption. The winter eggs are deposited predominantly on the bark (Beament, 1951). Egg type is governed by photoperiod, temperature and nutritional status of the female, as ‘summer’ females exposed to cool conditions lay diapausing eggs (van de Vrie et al., 1972). Mites feeding upon undamaged leaves are well nourished and the incidence of diapause is then determined solely by photoperiod and temperature. The mites feed on plant sap and unless they are crowded they will mostly be found alongside the veins on the underside of the leaves (Blair and Groves, 1952). They puncture plant cells with their stylets and the contents are exuded due to turgor pressure. The depth reached by the stylets is approximately 70-120 µm (Avery and Briggs, 1968). Depth at which injury occurs is related to length of the stylet, the feeding time and population density. Continuous feeding leads to regular spots being formed and when these coalesce characteristic chlorotic areas develop, often referred to as the ‘bronzing’ effect (Jeppson et al., 1975). At high densities P. ulmi reduces the photosynthetic activity of leaves (Mobley and Marini, 1990). This damage can lead to current-year or second-year effects. Current-year damage, depending on the timing, duration and severity, can reduce levels of foliar nitrogen, cause premature leaf fall (Baker, 1984), reduce shoot growth (Briggs and Avery, 1968) and trunk diameter (Chapman et al., 1952) and most importantly lower fruit yields (Hardman et al., 1985; Croft et al., 1983; Baker, 1984). It can also adversely affect fruit quality, for example, size (Hoyt et al., 1979), skin colour, soluble solids, titratable acids and firmness (Ames et al., 1984). Second-year effects of mite damage are reduced bloom with consequent reductions in numbers of apples and yield (Lienk et al., 1956; Baker, 1984). Panonychus ulmi was not considered to be of economic importance prior to the 1940’s (Chant, 1966). After the war, the chlorinated hydrocarbons, particularly DDT, were widely used in orchard pest control (Baker, 1952). These were broad spectrum in action and persisted in the environment. During the 1960’s and 70’s P. ulmi became resistant against many compounds like omethoate and dimethoate (Sterk, 1994). The detrimental effect of chemical spray compounds on the natural enemies of P. ulmi is considered a main contributing factor to the sudden increase in red spider mite numbers across European orchards (Huffaker et al., 1969). In addition, improved management of orchards, including the use of artificial fertilisers, has increased the nutritional value of apple leaves leading to improved mite fecundity (van de Vrie et al., 1972). Panonychus ulmi may therefore be considered a man-made pest. In unsprayed or abandoned orchards where natural enemies are active, damaging levels of European red mite are seldom reached (Croft and Hoying, 1977). Although researchers have attributed the decline of P. ulmi populations to predators that have entered the orchard after toxic sprays are discontinued, the foliage in these orchards may be injured by disease or deficient in nutrients, so making the host plant less favourable for P. ulmi (Walde, 1995). It has been found that severe damage from P. ulmi occurs on fruit such as apples or pears that have a thin cuticle (Garman, 1923). Leaf surface texture and contours are important in the reproductive potential of spider mites. Such features may serve as tough impediments to feeding (van de Vrie et al., 1972). Blair and Groves (1952) stated that Bramley’s Seedling seemed to be the only cultivar which exhibited any degree of resistance. The two layers of palisade mesophyll in the leaves help mask damage caused by the mites. To date, little use has been made of any known host plant resistance in developing commercial apple varieties. Cultivars tend to be selected because of their high yielding potential and adaptability to modern growing systems rather than for their resistance to a particular pest or disease.With ever increasing public concern over the use of chemical pesticides in regard to fruit production, based on fears relating to build-up of chemical residues and also effects on non-target species and the environment, is driving the need for fruit growers to devise new methods of pest control. Integrated pest management is now the leading force behind insect pest control. This involves moving away from the reliance of chemical insecticides and encouraging the natural enemy biodiversity to help control invertebrate pest species populations (Cuthbertson et al., 2003a, b; Cuthbertson, 2004; Cuthbertson and Murchie, 2004a,b; Cuthbertson, 2005; Cuthbertson and Murchie, 2005). This will reduce the build-up of pesticide resistance within pest species and also reduce the amount of chemical pesticides required for their control, therefore helping to form a more environmentally sustainable ecosystem. Acknowledgement Dr. Andrew G. C. Cuthbertson was funded by a Department of agriculture and Rural Development (Northern Irland) Studentship. References
© 2005 Center for Environment and Energy Research and Studies (CEERS) The following images related to this document are available:Photo images[st05039f1.jpg] |
| |||||||||

{kind=link}