 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
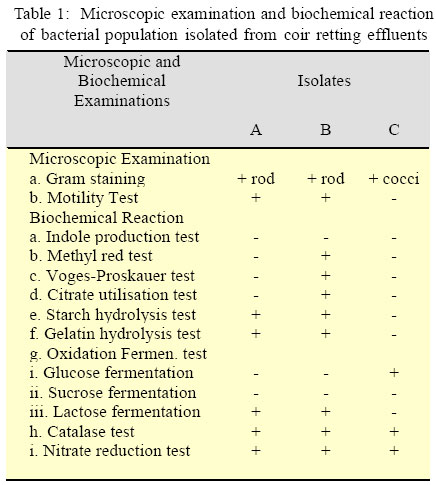
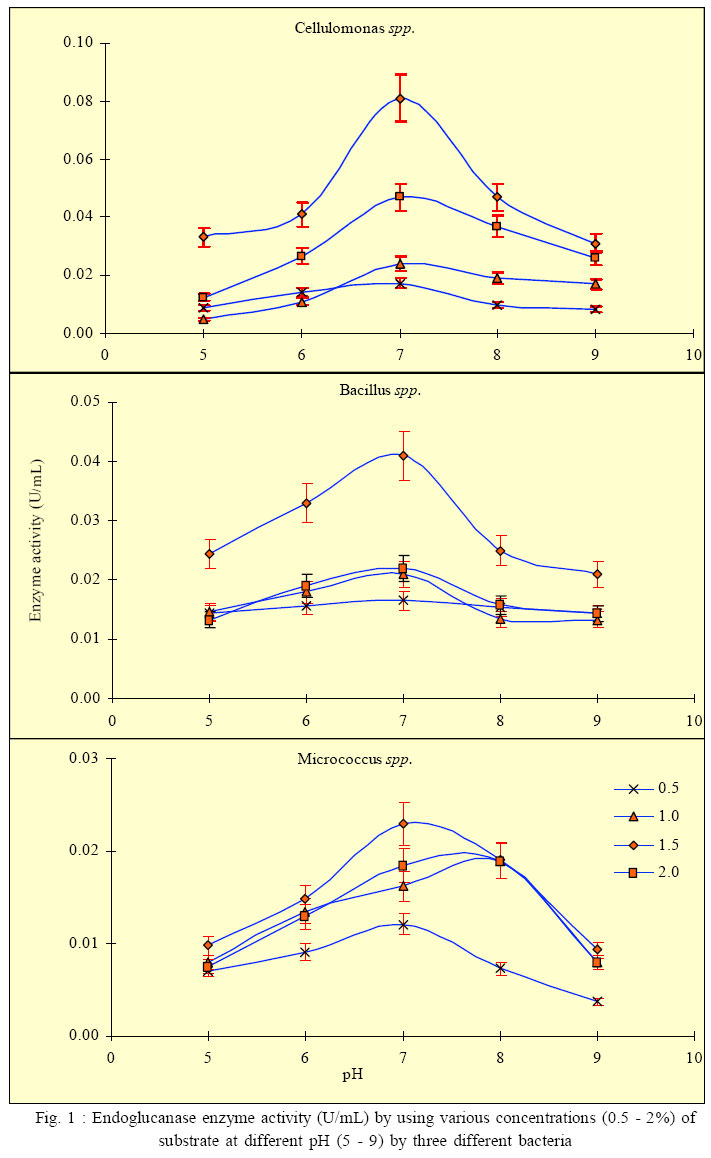
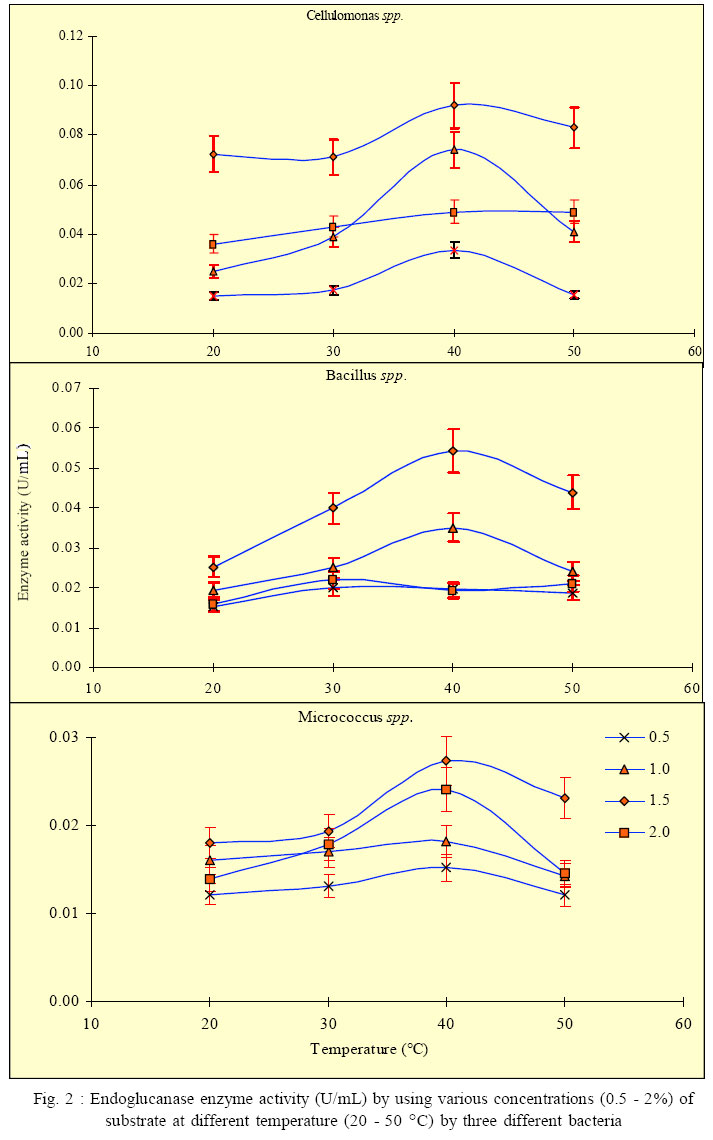
International Journal of Enviornmental Science and Technology, Vol. 3, No. 1, 2006, pp. 25-34 Effect of different growth parameters on endoglucanase enzyme activity by bacteria isolated from coir retting effluents of estuarine environment *G. Immanuel, R. Dhanusha, P. Prema and A. Palavesam Marine Biotechnology Division, Centre for Marine Science and Technology, M. S. University, Rajakkamangalam, Kanyakumari District, Tamilnadu, India *Corresponding Author, E-mail: g_immas@yahoo.com Code Number: st06004 ABSTRACT The cellulolytic enzyme-endoglucanase activity against coir fibre, a major biowaste by bacteria such as Cellulomonas, Bacillus and Micrococcus spp. isolated from coir retting effluents of estuarine environment was studied. The enzyme assay was carried out by using various concentrations (0.5 – 2%) of substrate of coir powder as a carbohydrate in different pH (5 – 9) and temperature (20 – 50 °C). The enzyme activity was minimum in 0.5% substrate concentration at lower pH 5 (0.0087, 0.0143 and 0.0071 U/mL) and at 20 °C temperature (0.0151, 0.0154 and 0.0122 U/mL) by the bacterial strains such as Cellulomonas, Bacillus and Micrococcus spp respectively. Then this level was increased and reached maximum at the neutral pH (0.0172, 0.0165 and 0.0121 U/mL) and at 40 °C (0.0336, 0.0196 and 0.0152 U/mL) by the selected bacterial species. Further increase of pH and temperature, the enzyme activity reduced considerably to 0.0083, 0.0143 and 0.0037 U/mL at pH 9 and 0.0154, 0.0197 and 0.0121 U/mL at 50 °C by the tested bacterial strains. The same trend was also obtained in oth er substrate concentrations such as 1.0, 1.5 and 2.0 %. With in the four substrate concentrations, the endoglucanase enzyme activity was more in 1.5% concentration at the tested pH and temperatures. From the over all result, it was observed that, among the three bacterial strains, the enzyme activity was more in Cellulomonas sp, followed by Bacillus and Micrococcus spp. in varying pH and temperature. Key words: Cellulolytic enzyme, Endoglucanase, coir retting effluent, Cellulomonas, Bacillus, Micrococcus INTRODUCTION Cellulose is probably the most abundant biological compound on terrestrial earth. It is the dominant waste material from agriculture industry in theform ofstalks, stems and husk and is one of the most abundant renewable sources. By means of chemical or bioconversion methods, it is possible to transform this insoluble polymer into glucose, an excellent substrate for industrial fermentation. Mainly bacteria, fungi and actinomycetes achieve bioconversion of these materials. Several studies were carried out to produce cellulolytic enzymes in biowaste degradation process by several microorganisms including fungi such as Trichoderma sp., Penicillium sp., Aspergillus spp. respectively by Mandels (1975), Hoffman and Wood (1985), Brown et al. (1987), Lakshmikant and Mathur (1990) etc. Similarly cellulolytic property of bacterial species like Pseudomonas, Cellulomonas, Cellulovibrio and Sporocytophaga spp. was also reported (Nakamura and Kappamura, 1982). The specific cellulolytic activity shown by the bacterial species is found to be depending on the source of occurrence (Sexana et al., 1993). The cellulase system of the mesophilic cellulolytic aerobe, Cellulomonas fimi is one of the first studied and has since been one of the most studied bacterial cellulase systems (Whittle et al., 1982; O’Neill et al., 1986; Shen et al., 1995). Many spore-forming bacteria have been isolated from factors that have a feedstock from cattle waste (Sharma and Hobson, 1985), cow manure (Palop et al., 1989), woody biomass (Sleat and Mah, 1984), municipal solid waste(Cailliex et al., 1992) etc. Similarly many strains ofcellulolytic anaerobic bacteria have been reported from various sources as human colon (Skinner, 1960), estuarine sediments (Madden, et al., 1982), freshwater sediments (Leschineand CanaleParola, 1983), decomposing vegetation (Madden, 1983; Petidemange et al., 1984). Because of the common occurrence of these bacteria in various natural environments, they are responsible for vast amount of cellulose. Cellulase yields appear to depend on a complex relationship involving a variety of factors like inoculum size (Carbon source and cellulose quality), pH value, temperature, presence of inducers, medium additives, aeration and growth time etc. The particle size of cellulose can affect cellulase production by microorganisms (Greaves, 1971). Release of enzyme is directly proportional to the particle size of the substrate used, i.e. release of enzyme increases, when the particle size of the substrate decreases. Thus attention has been focused to study the cellulolytic activity and cellulase enzyme production by several microorganisms in various products as well as in various environments. Attempts to increase the production of cellulose enzymes from bacteria including several processes like mutation (Stewart and Leatherwood, 1976; Haggett et al., 1978), protoplast fission (Gokhale et al., 1984), optimization of medium composition and environmental factors (Prasetsan and Doelle, 1987) etc have also been made. The bioconversion of various complex cellulosic waste materials such as bagasse (Kansoh et al., 1999), corncob (Ojumu et al., 2003) and saw dust (Solomon et al., 1999) have been reported. Coir dust and fibres are also major biowastes of coir industries of India, discarded along with coir retting effluent to estuarine environment. Yet literature related to the coir fibre as a carbohydrate source and cellulolytic activity by microorganisms involved in coir retting process is not studied. Hence in the present study, the endoglucanase enzyme activity of bacteria like Cellulomonas, Bacillus and Micrococcus spp. associated with coir retting process against these polymers were studied with varying environmental factors such as pH and temperature. This work have carried out this work during September 2004 to May 2005 in the Department of Marine Microbiology as well as Marine Biotechnology, Centre for Marine Science and Technology in India. MATERIALS AND METHODSCoir retting effluent collection and carbohydrate sample preparation The substrate used for this study was coconut fibre powder collected from coir retting industries in Rajakkamangalam area of K. K. District, South India. The substrate was sun dried for few days to reduce the moisture content and pretreated as described by Gharpuray et al., (1983). The resultedpowdery material was used as a carbohydrate source. The coir retting effluent was collected asepticallyfrom the discharging point of Rajakkamangalam estuarine environment in sterile bottles with proper cooling for bacterial isolation. The collected effluent samples were then brought to the laboratory and stored at 4 °C for further studies. Isolation and identification of cellulose degrading bacteria 1 mL of coir retting effluent sample was inoculated into 100 mL of Berg’smedium (NaNO3 2 g/L, MgSO4 0.5 g/L, K2HPO4 0.05 g/L, Fe2SO4 0.01g/L, CaCl2 0.02 g/L, MnSO4 0.02 g/l, pH 7) in a 250 mL conical flask and incubated at 30 °C for 5 days with constant shaking and 1 mL of this culture broth was inoculated in the same manner. This culture was repeated for 3 times and then the culture broth was spreaded on Berg’s agar plates (0.1 mL). The cellulolytic organisms were isolated by overlying plates with cellulose paper (Whatman No: 1) as carbon source. The cellulolytic organisms were identified by the appearance of yellow colour colonies over the filter paper. The same type of yellow colour colonies were streaked on Carboxy Methyl Cellulose (CMC) agarplates (NaNO32 g/L, MgSO4 0.5 g/L, K2HPO4 0.05 g/L, Fe2SO4 0.01 g/L, CaCl2 0.02 g/L, MnSO4 0.02 g/ L, CMC 1%,Agar 2%, pH 7) and incubated for 24 h at 30 °C. The isolatedcolonies on these plateswere maintained on CMC agar slants at 4 °C for further analysis. Screening of cellulase activity The isolated bacterial colonies were further characterized for their morphological and biochemical characters as described by (Cappuccino and Sherman, 1999).The purified colonies were further screened for their cellulase activity. Pure cultures of bacterial colonies weretransferred individually on CMC agarplates. After incubation, the plates were flooded with 1% congored and the plates were allowed to stand for 20 min at room temperature. Then the plates were thoroughly washed with 1 M Sodium chloride solution. A clear zone was evident around the growing colonies of cellulase positive cultures against the dark red background, the unreacted cellulose taking up the stain while the hydrolyzed portion remain colourless. The contrast was further enhanced by treating the plates with 5% acetic acid for 1 to 2 minutes and then washed off the excess acid with distilled water. Then those bacteria that were of good clearance beyond the area of growth were selected for further studies as potential cellulase secretors. The isolated bacterial colonies were further identified as Cellulomonas, Bacillus and Micrococcus spp. These three bacterial cultures were selected for estimation of extracellular cellulase – endoglucanase enzyme secretion. Inoculum preparation A pure culture of Bacillus, Cellulomonas and Micrococcus spp. were maintained as stock culture, from which inocula wereprepared. They were grownon CMC agar slant individually at 30 °C for 5 days and stored at 4 °C with regular subculturing. 200 mL of the optimised medium with each strain from a 4 day culture was used as inoculum prepared in a 250 mL conical flask. The inoculum was shaken continuously on an environmentcontrolled incubator shaker (New Brunswick Scientific Co., USA) at 200 rpm and 35 °C for 24 h before it was used for the process. Determination of endoglucanase activity Endoglucanase was assayed as described by Wood and Bhat (1988). The culture broth centrifuged at 5000 rpm for 20 min at 4 °C and the supernatant served as the enzyme source. The enzyme solution was added to 0.5 ml of 1% carbohydrate (coir powder) substrate taken in 50 mM sodium citrate buffer (pH 7). After incubation at40 °Cfor 30 min, the reaction was stopped by the addition of 1 ml dinitrosalicylic acid reagent. After 5 minutes in a boilingwater bath and quick cooling to room temperature, the degreeof enzymatic hydrolysis ofthe cellulose was determinedspectrophotometrically (UV-Vis Spectrophotometer, Techomp Scientific Equipment Ltd., Canada) by measuring the absorbance at 540 nm.The unitofenzymeactivity was expressed as the amount of enzyme required to release1 µmol of reducing sugars as glucose equivalent min/g of the enzyme sample. Optimization of production parameters For the effect of pH, aliquots of the enzyme solution were incubated with 500 μl of different concentrations (0.5, 0.1, 1.5 and 2.0%) of carbohydrate substrate (coir powder) at 37 °C at different pH values (pH 5 and 6 using 50 mM sodium citrate;pH 7 uisng 50 mMsodium phosphate and pH8 and 9 using50 mM Tris/HCl). The reactions were stopped after 30 min by adding dinitrosalicylic acid reagent and the activities were assayed as described above. The temperature optimum for the endoglucanase was investigatedwith different concentrations (0.5, 0.1, 1.5 and 2.0%) of carbohydrate substrate (coir powder), in 50 mM sodium citrate buffer (pH 7), were incubated for 30 minutes at 4 different temperatures in the range 20, 30, 40 and 50 °C. The reducing ends formed were determined using the dinitrosalicylic acid reagent as describedabove. Statistical analysis All experiments were performed in triplicate. The results obtained in the present study were analyzed through Two-way ANOVAtest following Zar (1974). RESULTSBased on the microscopic examination, physical and biochemical reactions, three different cellulosedegrading bacterial genera such as Cellulomonas, Bacillus and Micrococcus spp. were identified from coir retting effluent (Table 1). All the three bacterial strains were characterized for their ability to produce endoglucanase enzymes against coconut fibre powder as a carbon substrate and optimized their enzyme production with various growth factors such as substrate concentration, pH and temperature. Effect of pH on endoglucanase enzyme activity a. Cellulomonas spp. The endoglucanase enzyme activity by Cellulomonas sp at 0.5% substrate concentration at varying pH level showed that the activity was increased from pH 5 to 7 (0.0087, 0.0142 and0.0172U/mL).Further increasein pH level to 8 and 9 resulted in considerable decrease in enzyme activity(0.0097 and 0.0083 U/mL).Similar trend was observed at 1% substrate concentration, i.e. the maximum activity of 0.024 U/mL at pH 7. In 1.5% substrate concentration, the enzyme activity was 0.0331 U/mL at pH5, 0.041 U/mLat pH6, 0.081 U/mLatpH7, 0.047 U/mL at pH 8 and 0.031 U/mL at pH9. In 2% substrate concentration also, the endoglucanase enzyme activity showed by Cellulomonas spp. was minimum atpH 5(0.0125U/mL) and maximum (0.047U/ mL) at pH7 (Fig. 1).Among the four differentsubstrate concentrations, the enzyme activity was more in 1.5% substrate concentration. Statistical analysis by two way ANOVA on endoglucanase enzyme activity by Cellulomonas sp. revealed that the variation due to substrate concentration as well as pH level were statisticallysignificant (F =18.553; P<0.0001 and F = 6.239; P < 0.05). Further it also reveled that the influence of substrate concentration on endoglucanase activity was more than that of the influence exerts by variation in pH. b. Bacillus spp. The endoglucanase enzyme activity by Bacillus spp. at 0.5% substrate concentration was much higher (0.0165 U/mL) on neutral pH 7; whereas it was less in other tested pH levels. Likewise at 1% substrate concentration, the enzyme activity was minimum at pH 5 (0.0146 U/mL) and it showed a marginal increase (0.0180 and 0.0210 U/mL) at pH 6 and 7. Further this activity was greatly reduced to 0.0134 and 0.0132 U/mL at pH 8 and 9 respectively. In 1.5% substrate concentration, the enzyme activity was lesser at pH 5 (0.0243 U/mL) and it was 0.041 U/mL at pH 7 and at pH 8 and 9, it gradually decreased to 0.025 and 0.021 U/mL. The trend observed for the endoglucanase enzyme activity by Bacillus spp. at 2.0% substrate concentration was more or less same with that of observed for 0.5, 1.0 and 1.5% substrate concentrations (Fig. 1). Among the four different substrate concentrations tested, the activity was more at 1.5%. The two-wayANOVA revealed that the endoglucanase enzyme activity due to varying substrate concentrations was statistically more significant (F = 22.477; P < 0.0001) than the variation due to different pH levels (F = 5.703; P < 0.005). c. Micrococcus spp. The endoglucanase activity shown by Micrococcus sp. inferred that, at 0.5% substrate level, the enzyme activity was 0.0071 U/mL in pH 5 and it increased to 0.0091 and 0.0121 U/mL at pH 6 and 7. Then it was decreased to 0.0073 and 0.0037 U/mL at pH 8 and 9. Similar trend was obtained in other substrate concentrations also (1, 1.5 and 2.0%) (Fig. 1). The twowayANOVA revealed that the endoglucanaseenzyme activity by Micrococcus spp. due to the variation of substrate concentrations as well as due to the variation of pH level were statistically more significant (F = 11.116; P < 0.005 and F = 19.671; P < 0.0001). Effect of temperature on endoglucanase enzyme activity a. Cellulomonas spp. At 0.5% substrate concentration, the enzyme activity was more at 40 °C (0.0336 U/mL), whereas it was less in 20 °C (0.0151 U/mL), 30 °C (0.0172 U/mL) and 50 °C (0.0154 U/mL). In 1% substrate concentration also, the enzyme activity was moreat 40°C (0.0740 U/mL), and less (0.025, 0.039 and 0.041 U/mL) at 20,30 and 50 °C.In the case of 1.5 and 2% substrate concentrations, the enzyme activity was 0.0723 and 0.0360 U/mL at 20 °C, 0.0710 and 0.0430 U/mL at 30 °C, 0.0920 and 0.0490 U/ mL at 40 °C and 0.0830 and 0.0490 U/mL at 50 °C (Fig. 2). The statistical analysis (Two-way ANOVA) revealed that the enzyme activity due to different substrate concentrations and due to various temperatures were statistically more significant (F = 39.015; P < 0.0001 and F = 7.587; P < 0.05). b. Bacillus spp. The enzyme activityby Bacillus spp. at 0.5% substrate concentration showed maximum of 0.0190 U/mL at 40 °C, whereas it was less (0.0154, 0.020 and 0.0197 U/mL) at 20, 30 and 50 °C temperatures respectively. In 1% substrate concentration, the enzyme activity was 0.0194 U/mL at 20 °C and it increased to 0.025 and 0.0351 U/mL at 30 and 40 °C, respectively, again the activity decreased to 0.0241 U/mL at 50 °C. In 1.5 and 2% substrate concentrations, more or less a similar trend was observed in the activity of endoglucanase (Fig. 2). The statistical test carried out bytwo-wayANOVA revealed that the endoglucanase enzyme activity due to the variation of substrate concentrations was statistically more significant (F = 14.018; P< 0.005) than the variation due to temperature (F = 4.130; P < 0.05). c. Micrococcus spp. The enzyme activity in Micrococcus spp. at20 °C was 0.0122, 0.161, 0.0181 and 0.0139 U/mL in the tested substrate concentrations (0.5 to 2.0%). Similarly at 30 °C, it ranged from 0.0131 to 0.0194 U/mL at 0.5 to 2% substrate concentrations. But at 40 °C, the enzyme activity waslittlehigh (0.0152, 0.0182,0.0274 and 0.0241 U/mL) at the tested substrate concentrations (0.5 to 2.0%). In 50 °C also, the enzyme activity was less than that of recorded at 40 °C and it ranged from 0.0121 to 0.0231 U/mL at the tested substrate concentrations (0.5 to 2.0%) (Fig. 2). Among the four different temperatures tested, 40 °C is the optimum temperature for maximum enzymeactivity.Similarly 1.5% substrate concentration was also found to be optimum for enzyme activity. The analysis of variance (Two- way) revealed that the endogluconase enzyme activity due to the substrate concentrations was statistically significant (F = 11.223; P < 0.05). But enzyme activity due to the variation in temperature was not statistically significant (F = 6.272; P > 0.01). DISCUSSION AND CONCLUSION In the present study, three bacterial strains such as Cellulomonas, Bacillus and Micrococcus spp. were identified for their ability to produce endoglucanase enzyme against pretreated coir fibre powder, isolated from coir retting effluent. The increase of cellulase activity was studied by optimization of enzyme production at various environmental factors such as temperature and pH. Cellulose serves as a carbon source for the growth of bacteria and also as an inducer for the production of cellulase. Some previous studies reported that the agricultural wastes of lignocellulosics are used as a carbohydrate source to produce commercially important products such as ethanol, glucose and single cell protein (Solomon et al., 1999). Similarly Ojumu et al., (2003) reported some lignocellulosics as carbon source for cellulase enzyme production. They reported 0.0743 U/mL of cellulase enzyme activity from sawdust and 0.0573 and 0.0502 U/mL from bagasse and corncob respectively. In the present study also various concentrations (0.5 - 2%) of pretreated coir fibre powder as a carbon source was used to find out the enzyme activity. Among the tested concentrations 1.5% substrate showed higher enzymatic activity (0.081, 0.041 and 0.023 U/mL at pH 7 for Cellulomonas, Bacillus and Micrococcus spp. respectively). The enzyme with very low activity was accounted from much lower and higher concentrations. Similar reports from few experiments showed that the enzyme activity in Bacillus spp. was detected in cultures that contained 1% (w/v) CMC as the growth substrate (Fukumori et al., 1985; Kawai et al., 1988; Shikata et al., 1990). In accordance with these, reports of enzyme activity by Strepetomyces spp. isolated from soil samples with various substrate (CMC) concentrations (0.5-1.5%) showed that maximum activity was achieved at 1% substrate concentration (Hankin and Anagnostakis, 1977). But some new studies on Sinorhizobium fredii, reported that 0.5% of substrate concentration showed better enzyme activity (Po-Jui Chen et al., 2004). Robson and Chambliss (1984) reported that the presence of metabolized carbohydrates in the growth medium stimulated the production of cellulolytic activity. The enzyme hydrolyzes substrate in the pH range
of 4.0 to 9.0, with maximum activity occurring at pH 7.0.
The enzyme maintained their stability over a wide range
of pH 6-8, but it had maximum activity at pH 7. GoKhan
Coral et al. (2002) reported that maximum CMCase
activity was recorded at pH 7.5 by Aspergillus niger
(Z10 wild type strain) when among the tested pH range
between 4-9. Similarly the optimal condition of pH 6-7
for maximum enzyme activity in A. niger also reported
by Akiba et al., (1995). Alkaline pH value (pH 12) also
affected the enzyme activity in Bacillus spp. by 50%
and furthermore, the CMCase in Bacillus spp. is
alkaline, which renders it suitable for use as an effective
laundry detergent additive (Khyami-Horani, 1991).
Some previous studies from Garcia-Martinez et al.
(1980) and Prasetson and Doelle (1987) reported that
pH 7 appears to play a decisive role in cellulose
digestion for maximum enzyme production by
Clostridium thermocellum and Cellulomonas spp. Like pH, temperature is also an important factor that influences the production of enzyme. In the present study, the optimum temperature was found to be around 40 °C and the enzyme activity of 0.092, 0.0543 and 0.0274 U/mL at pH 7 and 1.5% substrate concentration were recorded for Cellulomonas, Bacillus and Micrococcus spps. respectively. Gokhan Coral et al., (2002) reported, the optimum temperature for CMCase activity was 40 °C in A. niger Z10 strain. But these values were lower than that of the commercial cellulase production temperature of 60 °C and the optimum temperature required for enzyme production by bacterial strains as well as fungal strains are entirely differed. Cellulase activities from Trichoderma sp. and other mesophilic cellulolytic fungi are at their optimum when assayed at about 50 °C (Mandels et al., 1974; Tangnu et al., 1981; Kawamori et al., 1987). However, the optimum temperature for Trichoderma sp. A-001 was at least 10 °C higher than the others (Durand et al. 1984; Margaritus and Merchant, 1986). In conclusion, the present study revealed that coconut fibre powder is one of the best carbohydrate substrates for endogluconase enzyme production by the organisms viz. Cellulomonas, Bacillus and Microccocus spp. The organisms were readily utilized this substrate and the present study confirmed that 40°C is the optimum temperature at 1.5% substrate concentration with neutral pH for higher endoglucanase activity. REFERENCES
© 2006 Center for Environment and Energy Research and Studies (CEERS) |

{kind=link}
{kind=link}
{kind=link}