 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
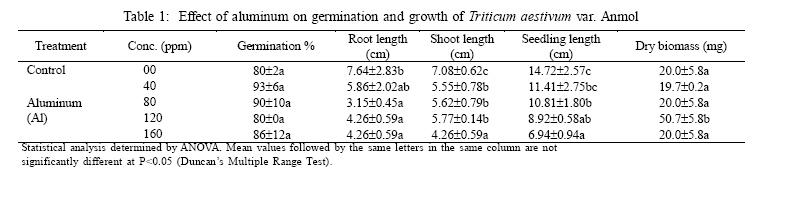
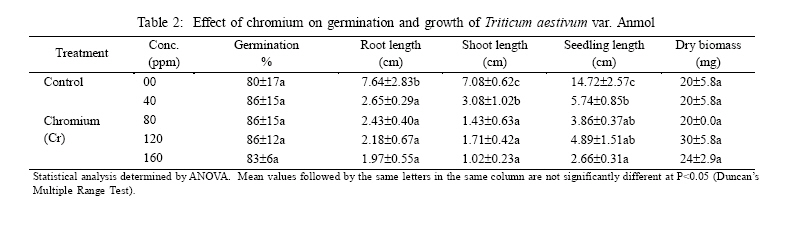
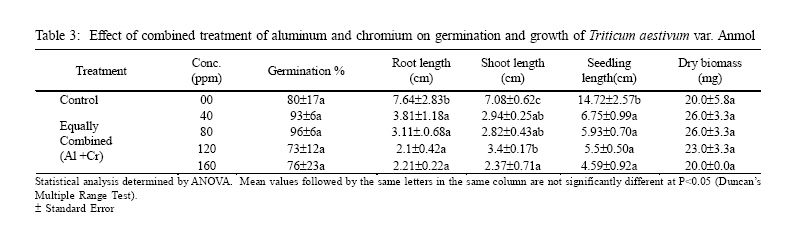
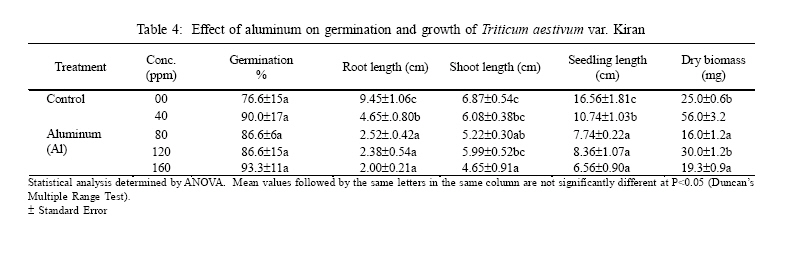
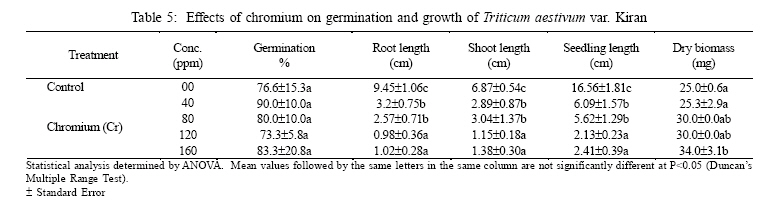
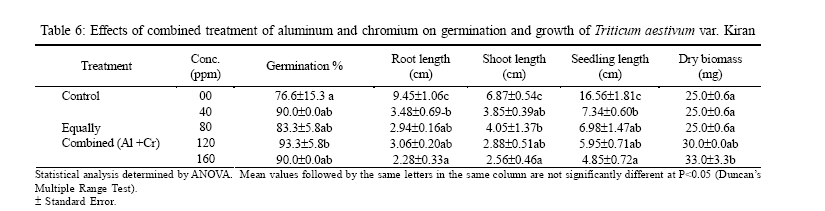
International Journal of Enviornmental Science and Technology, Vol. 3, No. 4, Autumn 2006, pp.411-416 Phytotoxic effect of aluminum and chromium on the germination and early growth of wheat (Triticum aestivum) varieties Anmol and Kiran 1S. N. Jamal, 1M. Z. Iqbal and *2M. Athar 1Department of Botany, University of Karachi, Karachi 75270, Pakistan Received 24 May 2006 Code Number: st06050 ABSTRACT: A greenhouse experiment was conducted to determine the phytotoxic effect of aluminum and chromium on the germination and early growth of two wheat (Triticum aestivum) varieties Anmol and Kiran. Seed were treated with 40, 80, 120 and 160 ppm of aluminum and chromium solution individually and in combined form. Observations were made on seed germination, root, shoot and seedling length, and dry biomass. Seed germination and dry biomass showed no effect of aluminum, chromium and combined treatment. Root, shoot and seedling length of both the varieties showed significant (P<0.05) decrease as compared to control. The growth was also reduced as the concentration of aluminum and chromium increased. Seedling length decreased in both the varieties at all the concentration of different treatment of aluminum, chromium and combined treatment. Attempts are being made in different laboratories to construct novel plants using genetic manipulation technologies that may have a greater tolerance to the presence of toxic metals. The results of the present study may help in understanding the mechanisms involved and their possible use in pytoremediation. Key words: Aluminum, chromium, wheat varieties germination, phytotoxicity INTRODUCTION Heavy metal phytotoxicity is considered to be the main factor limiting plant growth, and thus crop cultivation on acid soils (Foy, 1984, 1988). Acid soils comprise large area of the world’s agricultural regions, particularly in the tropics and subtropics. Soil acidity is an important factor limiting food production in developing countries (Kochian, 1995). Fox (1979) found that yield of maize was reduced at aluminum saturation of 50 %. Excess of aluminum is generally considered to be responsible for inhibiting growth and depressing yields on acid soils. Farina (1980) conducted liming experiment with maize and showed that aluminum toxicity can also be induced by liming reddish brown and ground lateric soils. Aniol (1989) studied the induction of aluminum tolerance in wheat seedling by low doses of aluminum in nutrient solution and found better root elongation. Chromium is considered as a strong toxic element. Chromium ions are tightly bound to humus and clay particles and are more or less insoluble in the soil. Its availability in plants is therefore generally low but mobility and availability are relatively decreased with the increasing pH. Mengel and Kirkby (1987) mentioned that 0.5 ppm of chromium as chromium sulphate stimulated growth in hydroponic experiment with maize. They found that the growth was inhibited at 5 ppm and strongly inhibited at 50 ppm. El-Bassam (1978) reported that low Cr+3 concentrations promote plant growth and also stimulate chlorophyll synthesis and photosynthetic activity. Hexavalent anionic form of chromium is several times as toxic as the trivalent cation Cr+3. Mechanism of resistance in plants continues to remain poorly understood. Such resistance arises either by the ability of the plants to exclude metals from the root or to detoxify metals within the plants (Koachin, 1995; Taylor, 1995). In wheat, malate exudation from root tips seems to be an important mechanism for aluminum detoxification (Delhaize, et al., 1993b). Osawa and Matsumoto (2001) showed that malate efflux started within 5 minutes after the addition of aluminum in wheat. Several economically important cereal grasses such as Hordeum vulgare, Triticum aestivum and Zea mays have been particularly useful in such studies (Farina, 1980; Mossor-Pietraszewska, 2001 and Sharma and Mehrota, 1993).A number of studies have been conducted on relative root growth inhibition to define categories of sensitivity and to identify more tolerant cultivars of wheat (Basu, et al., 1994; Delhaize, et al., 1993a, b). Wheat (Triticum asetivum L.) is the staple food for a large part of the world population including Pakistan. Wheat is grown in Pakistan on 8.141 million hectares with an average yield of 2.28 tons per hectare wit a total production of 18535 thousand tons (Agricultural Statistics of Pakistan, 2003, 2004). The average yield is below than most wheat producing countries like Germany (7.9 tons per hectare), France (6.6 tons per hectare) and Egypt and Germany (6.4 tons per hectare). With increasing population, the over-demanding production of wheat can be increased either by bringing more area under wheat cultivation or by introducing high yielding wheat varieties which are resistant against biotic and abiotic environmental stresses. This study constitutes part of a broader research project to focusing on testing the effect of heavy metals on plant growth (Jamal, et al., 2006a, b, c). The present study was carried out to determine the effect of aluminum and chromium individually and in combination on seed germination and seedling growth of two wheat varieties. MATERIALS AND METHODS The seeds of two wheat (Triticum aestivum) variety Anmol and Kiran were obtained from Pakistan Agricultural Research Council, Islamabad. The seeds were surface sterilized with 1.2 % sodium hypochloride solution. Aluminum was applied in the form of aluminum nitrate and chromium wasapplied in the form ofpotassium dichromate (K2Cr2O7) following methods reported elsewhere (Jamal, et al., 2006a, b). Aqueous solutions of both the metals were prepared at concentration of 40, 80, 120 and 160 ppm. Control was free from these metals. Fifteen seeds of each cultivar of wheat were placed in each petri plates and supplied with 3 mL aluminum and chromium solution applied individually andin combined form. The petri plates were kept in the dark and observed for germination. The seeds were considered germinated with the emergence of radicles. The germinated seeds were then transferred todayand night length of10/14 h respectively illuminated with 160 watt light with temperature regime of about 31±2 oC. The average relative humidity was 75% as recorded by Sling psychrometer in the laboratory. Root, shoot and seedling length were measured after 10 days, and the seedling weight was recorded after drying in an oven at 80 oC for 24 h. Data were analyzed statistically by ANOVA and Duncan’s multiple range tests (Sokal and Rohlf, 1995). RESULTS Seeds of both the wheat varieties Anmol and Kiran germinated well under all the treatments (Tables 1, 2, 3, 4, 5, and 6). All the seeds germinated next day. Seed germination of both the varieties was elevated at different concentration as compared to control (Tables 1, 2, 3, 4, 5, and 6). Control showed 80% germination and aluminum, chromium and combined treatments showed 90% germination. Results showed that both the aluminum and chromium metals individually andin combined form significantly (P<0.05) affected root, shoot and seedling length of both the varieties as compared to control (Tables 1, 2, 3, 4, 5, and 6). Chromium significantly (P<0.05) reduced the growth of the seedling than aluminum and combined treatments. Root length of variety Kiran was significantly (P<0.05) inhibited at all the treatments of aluminum and chromium as compared to control. Combined treatments showed comparatively better effect on root length of Kiran than the individual chromium treatment. Root length of variety Anmol was slightly larger than root length of variety Kiran. Aluminum treatment showed more or less better effect than chromium and combined treatment as compared to control. Shoot length of both the varieties Anmol and Kiran was inhibited by the individual chromium and combined treatments but aluminum treatment enhanced the shoot growth (Tables 1, 2, 3, 4, 5, and 6). All the treatments showed inhibition in both the varieties as compared to control whereas growth was significantly (P<0.05)reduced bychromium. However, the combined treatment showed the intermediate effect (Tables 1, 2, 3, 4, 5, and 6). Seedling length of both the varieties also showed similar inhibitory effect in all the treatments (Tables 1, 2, 3, 4, 5, and 6). The aluminum treatment showed enhanced and chromium treatment produced reduced growth. Different concentrations showed inhibition as compared to control. The length of the seedlings of both the varieties was decreased with increasing concentration of the metals. The aluminum and chromium treatments did not show any inhibitory or enhanced effect on dry biomass (Tables 1, 2, 3, 4, 5, and 6). Chromium and combined treatment showed similar effect whereas aluminum showed a little increase in biomass in both the wheat varieties. DISCUSSION AND CONCLUSION Both the wheat varieties Anmol and Kiran showed similar germination response when chromium and aluminum were applied individually and in combined form. The germination of both the varieties was not inhibited by chromium and aluminum treatments. These metals have been reported not to inhibit germination but impair the growth of new roots and seedling establishment (Nosko, et al., 1988 and Rellén-Álvarez, et al., 2006). The growth of both the varieties showed inhibitory effect of chromium and aluminum. Chromium was found more toxic affecting the growth of root, shoot and seedling length than aluminum and combined treatments. Chromium is considered strongly toxic because chromium compound in the soil are more or less insoluble as the metal ions are tightly bound to humus and clay particles. Its availability to the plants is generally low as the mobility is relatively decreased with increasing pH. Mengel and Kirkby (1987) reported that 0.5 ppm of chromium as Cr+3 stimulated growth in hydroponic experiment with maize whereas growth was inhibited at 5 ppm and strongly affected at 50 ppm. El-Bassam (1987) observed that low chromium concentration promote plant growth and also stimulate chlorophyll synthesis and photosynthetic activity. Hexavalent ionic form of chromium was as toxic as trivalent cation Cr+3. Aluminum also showed inhibitory effect as compared to control but overall enhanced the growth than chromium and combined treatments. Root stunning is a consequence of aluminum induced inhibition of root elongation. Roots became stubby and brittle and root tips and lateral roots became thick and turned brown. Such roots are in efficient in absorbing nutrients and water affecting the growth of shoot than the root. Similar observations were made by Chang, et al. (1999) made similar observations for tobacco treated with a combination of aluminum and iron. Mechanism of resistance in plants continues to remain poorly understood. Such resistance arises either by the plant ability to exclude heavy metals in roots or its ability to detoxify heavy metals within the plants(Hall, 2000; Kochian, 1995; Rellén-Álvarez, et al., 2006 and Taylor, 1995). Detoxification also depends upon organic compounds present in soil and inside plants. Wheat plants are very sensitive to these heavy metals as they have very little external ability to detoxify the toxic metals. Wheat seeds were germinated in the petri plates which were devoid of other minerals. The metals applied did not get detoxified and were absorbed by plants which resulted in the inhibition of root, shoot and seedling length. Osawa and Matsumoto (2001) found that in wheat malate efflux started 5 min after the addition of aluminum in wheat. Ma, et al. (1997) observed that in buckwheat, the secretion of oxalic acid occurred within 30 min after the exposure to aluminum. Organic acid extracted from roots has different ability to precipitate aluminum and reduced toxicity. In wheat and maize, aluminum activated anion channels in the plasma membrane mediate the transport of organic acid anions out of the root cells. Plants have evolved several mechanisms to prevent the toxic action of metals. These include reduction of uptake into the root cells by changes in the kinetic properties of transporters and exudation of complexing agents into the rhizosphere (Watanabe and Osaki, 2002). Once metals have entered the plant they induce the synthesis of the glutathione containing peptides phytochelatins and high cysteine containing proteins, the metallothioneins, which are able to bind the metal ions (Cobbet and Goldsbrough, 2002; Hall, 2002; Quartacci, et al., 2000 and Voskaoboinik, et al., 2002). Ultimately the metals are stored in the vacuole in a relatively non-toxic form (William, et al., 2000). There have been a number of attempts by different laboratories to construct novel plants using genetic manipulation technologies that may have a greater tolerance to the presence of toxic metals (Belouchi, et al., 1997; Macnair, et al., 2000; Sasaki, et al., 2004 and Vitorello, et al., 2005). These plants are currently under intensive study to establish the mechanisms involved and for their possible use in pytoremediation. ACKNOWLEDGMENT The research was funded by a grant from the Faculty of Science, University of Karachi, which is gratefully acknowledged. Special gratitude is expressed to the scientists of Plant Genetic Resources, and National Wheat Program of Pakistan Agricultural Research Council, Islamabad for supplying the seeds of wheat varieties Anmol and Kiran. REFERENCES
© 2006 Center for Environment and Energy Research and Studies (CEERS) The following images related to this document are available:Photo images[st06050t5.jpg] [st06050t6.jpg] [st06050t4.jpg] [st06050t3.jpg] [st06050t1.jpg] [st06050t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}