 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
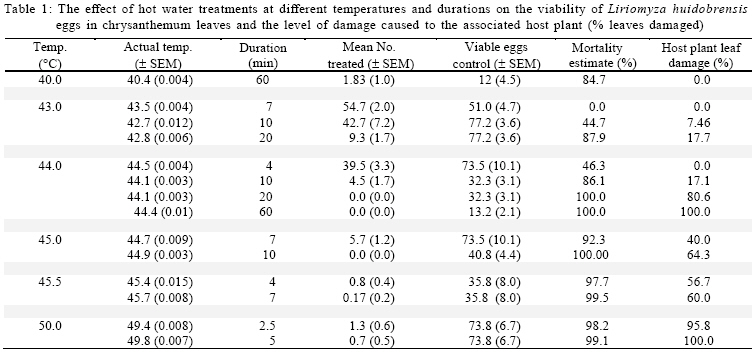
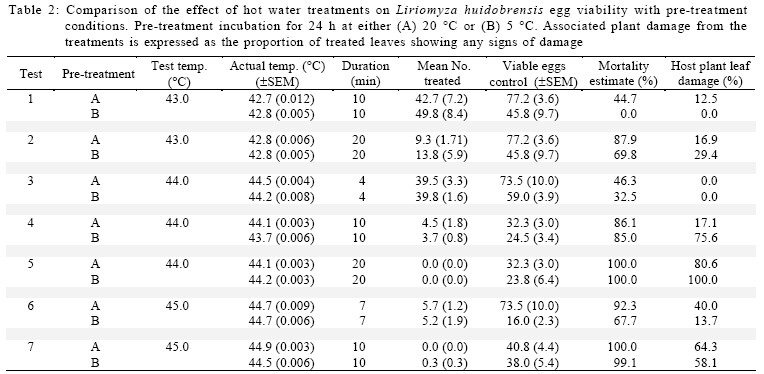
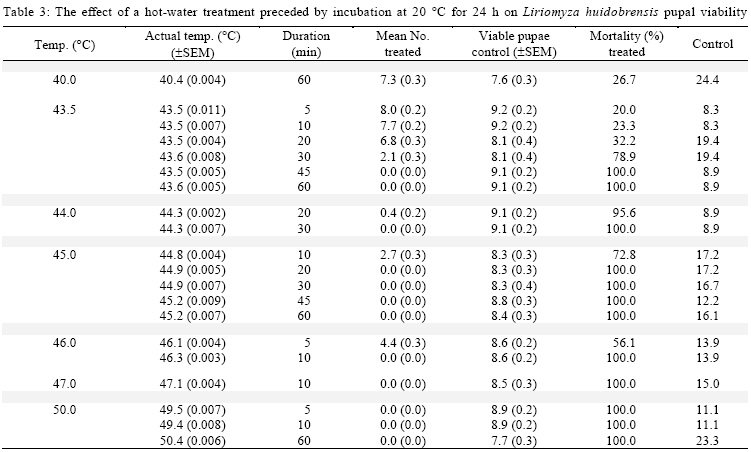
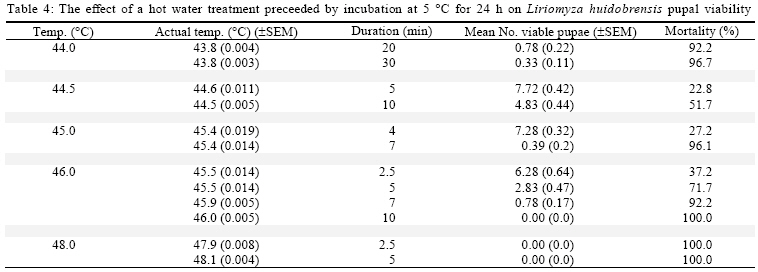
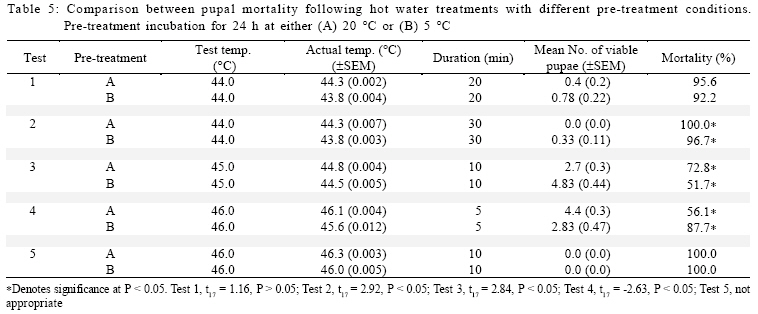
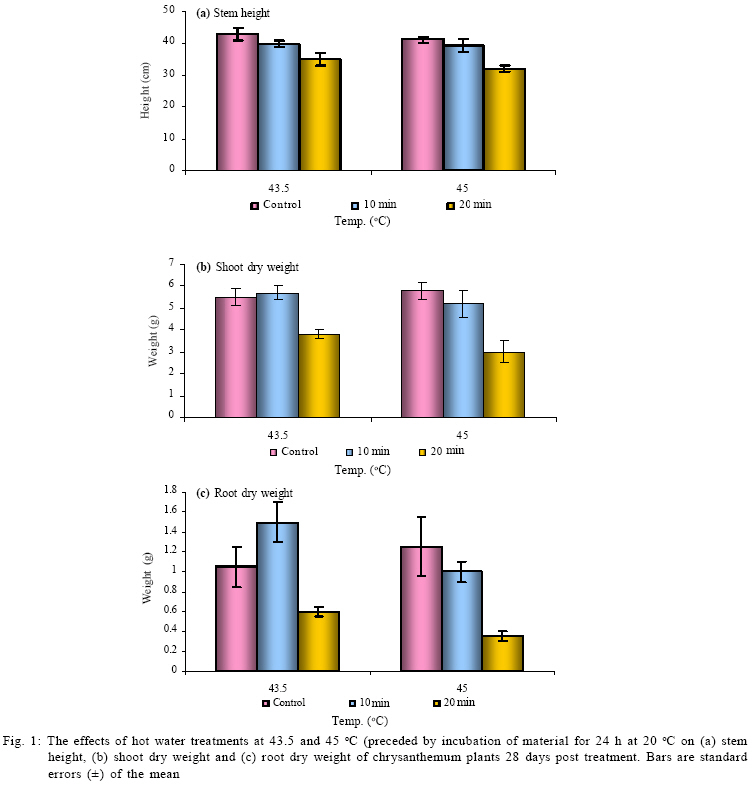
International Journal of Enviornmental Science and Technology, Vol. 6, No. 2, Spring, 2009, pp. 167-174 Environmental evaluation of hot water treatments to control Liriomyza huidobrensis infesting plant material in transit *A. G. S. Cuthbertson; L. F. Blackburn; P. Northing; J. J. Mathers; W. Luo; K. F. A. Walters Central Science Laboratory, York, England, UK Received 12 August 2008; revised 28 November 2008; accepted 20 January 2009; available online 1 March 2009 Code Number: st09017 ABSTRACT Though quarantine disinfestation treatments have been exempt from the methyl bromide phase out, it is still required to research and develop alternative treatments for fumigation of plant material in transit. This study investigated the ability of both the egg and pupal stages of the quarantine pest, South Americian leafminer, Liriomyza huidobrensis to survive submersion in water heated to temperatures between 40 and 50 °C for varying periods of time. Large reductions in egg viability were recorded. However, the treatments also resulted in unacceptable levels of damage to the host plant material. Damage was uneconomically viable. Pupae were all killed exposure to water at 44 °C for 20 min. Incubation of insect and plant material at either 5 or 20 °C for 24 h before submersion in the hot water did not significantly alter the pests’ ability to survive the treatments. The potential of hot water treatments to act as an alternative to methyl bromide fumigation for disinfecting plants in transit is discussed. Keywords: Disinfestations, quarantine, pest management, methyl bromide, pests INTRODUCTION Increased international plant trade is considered as a major mechanism of pest dispersal with numerous pests and diseases being spread around the world (Lenteren and Woets, 1988; Neumann and Elzen, 2004). Within the European Union, a legislation exists that prohibits the movement of specific organisms of plant health importance (Anonymous, 2000; Bartlett, 1993; Cheek and Cannon, 2002). In the UK, various pests including Liriomyza huidobrensis Blanchard (Diptera: Agromyzidae) remain notifiable pests subject to a policy of eradication if found on propagators premises or plants moving in trade, and containment if outbreaks occur at nurseries (Cuthbertson, 2005). To meet these legislatory requirements, effective quarantine disinfestation treatments for traded plant material are essential. The majority of such treatments have relied on methyl bromide (Lopez-Aranda et al., 2003; Walters, 2001). However, although methyl bromide usage for quarantine purposes is exempt from being phased out (Schneider et al., 2003) if no viable alternative is found, there is still a demand for alternatives to methyl bromide for invertebrate pest control to be devised (Giannakou and Karpouzas, 2003; Lopez-Aranda et al., 2003; Slusarski and Pietr, 2003). Various types of post harvest treatments have been evaluated for the disinfestations of fruit and vegetables against a range of pests. These include alternative fumigants such as carbonyl sulphide and methyl iodide (Aung et al., 2001), vapour heat (Hallman, 1990) and hot water immersion (Gould and Sharp, 1992; Hallman, 1994; Hara et al., 1993; 1994; Jang, 1991; Jones et al., 1995; Liquido, 1990; Nascimento et al., 1992; Sharp et al., 1989a,b; Sharp and Picho-Martinez, 1990). However, few studies report the effect of treatments on insects associated with cut flowers and planting material moving in trade. Hansen et al. (1992) evaluated the effect of vapour heat as a potential treatment to disinfest tropical cut flowers and foliage infested with aphids, mealybugs, scale insects and thrips. However, the ability of flowers and foliage to tolerate the various treatments varied greatly, therefore, there is a need to test each specific commodity during the development of a disinfestation method. In the current study, investigations were conducted to determine the effect of immersion in hot water on the egg and pupal stages of the South American leafminer, L. huidobrensis and the associated plant material, chrysanthemum (Dendranthemum grandiflora L. cv. White Fresco). To determine whether the pre-treatment incubation temperature altered the susceptibility of the insects to the hot water treatments, samples were incubated at either 5 or 20 °C for 24 h prior to each treatment. The host plants used in all egg stage investigations were monitored for visual signs of damage caused by the treatments. In addition, a separate series of hot water treatments were conducted with plant material to obtain quantitative assessments of plant growth four weeks post-treatment. The experiments were undertaken in the Plant Health Quarantine Entomology Laboratories based at the Central Science Laboratory, UK, in 2000. MATERIALS AND METHODS Insect cultures Liriomyza huidobrensis was cultured under conditions similar to those used by Cuthbertson et al. (2003; 2005) for culturing Bemisia tabaci. Perspex cages (60 cm × 60 cm × 80 cm) held under quarantine conditions (Cuthbertson et al., 2008a, b) containing a variety of tomato (Lycopersicon esulentum), Chinese brassica (Brassica chinensis) and cucumber (Cucumis sativus) plants at 25 ± 1 °C with a 12:12 h light:dark (L:D) regime and an artificial dawn and dusk were used for culturing L. huidobrensis. Susceptibility of Liriomyza huidobrensis eggs to hot water treatments Batches of twelve chrysanthemum (cv. White Fresco) plants at the 6-8 leaf stage were exposed to 100 mixed sexed L. huidobrensis adults, all of which were more than 24 h post-eclosion and incubated at 20 °C, 16:8 L:D regime, 65 % relative humidity (r.h.). After a 24 h egg-laying period, the flies were removed before the plants were divided into two groups; treatment (n=6) and control (n=6). Plants in the treatment group were submersed into hot water maintained at a specific temperature which was monitored at five second intervals from three locations within the water bath. Leaf temperature was similarly recorded. Following the treatment, the plants were dipped in water at ambient temperature (20 ± 5 °C) for one min before they were incubated with the control group, which had been kept continuously at 20 °C, 16:8 L:D, 65 % r.h. After 7 days, egg survival was recorded as the number of leafminer larvae present in each leaf of each plant. As it was not possible to reliably and non-destructively determine the number of eggs present in each batch of infested, material egg mortality was calculated from the number of larvae emerging in the treated plants compared to those in the control group. The procedure was conducted with water heated to temperatures between 40 and 50 °C for periods ranging from 2.5 to 60 min and preceded by incubation of the experimental material at 20 °C for 24 h in controlled environmental chambers immediately before the hot water treatment (Table 1). In addition, seven tests were repeated to determine the effect of incubation at the lower temperature of 5 °C prior to hot water immersion (Table 2). To record the potential influence of host damage on egg survival, the level of plant damage was recorded. This was expressed as the proportion of leaves showing any signs of damage from yellowing to dead material and was recorded by eye. Susceptibility of Liriomyza huidobrensis pupae to hot water treatments Batches of 360 mixed age L. huidobrensis pupae were removed from the culture and groups of 10 were placed into water-tight 0.25 mL plastic tubes. Eighteen tubes were submersed in water at temperatures from 40 to 50 °C for periods ranging between 5 and 60 min. The temperature of the water and the air within the tubes were recorded at 5 sec intervals throughout the treatment. Following the treatment, all pupae were removed from the containers, placed onto damp sand in 90 mm Petri dishes and incubated at 20 °C. Adult emergence was recorded as a measure of pupal survival in the treatment and control groups. The procedure was repeated with water heated to a range of temperatures and preceded by incubation at either 5 or 20 °C for 24 h immediately before the hot water treatments (Tables 3 and 4). Phytotoxicity Batches of six chrysanthemum plants at the 6-8 leaf stage were incubated at either 5 or 20 °C for 24 h before they were subjected to one of four hot water treatments: 43.5 for 10 or 20 min; 45 °C for 7 or 10 min. Following the treatment, the plants were dipped in water at ambient temperature (20 ± 1 °C) for one min before they were incubated for 28 days with the control group, which had been kept continuously at 20 °C, 16:8 L:D regime, 65 % r. h. After this period, plant height along with root and shoot dry weights were recorded. Data from the groups of treated plants were compared against values of an equal number of control plants. Statistical analysis In comparisons with hot water treatments which were preceded by different incubation temperatures percentage mortalities were arcsine square-root transformed before analysis. Paired t-tests were used to compare the means from each set of tests. The plant phytotoxicity tests were analysed using ANOVA and Data were analysed using minitab statistical software. RESULTS AND DISCUSSION Susceptibility of Liriomyza huidobrensis eggs to hot water treatments The introduction of the infested plant material into the hot water resulted in a drop of temperature which returned to the desired treatment temperature within one min. High levels of mortality were recorded following the majority of investigated treatments (Table 1). Exposure to L. huidrobensis eggs to 43 °C for 20 min caused 87.9 % mortality with a low level of damage to the plants. 1 °C rise in water temperature resulted in no larvae emerging from the treated eggs, however, this is likely to be related to the high level of plant damage caused (80.6 % of leaves affected) by the test. A further increase in water temperature to 45.5 °C resulted in 97.7 % egg mortality following a very short exposure of 4 min with signs of plant damage recorded on 56.7 % of the treated leaves. All of the treatments which resulted in total egg mortality also lead to unacceptable levels of visual plant damage. Seven comparative tests were conducted to determine whether incubation at either 5 or 20 °C for 24 h prior to the hot water treatment altered the insects susceptibility to hot water. Egg mortality only differed significantly on one comparative test (Table 2, Test 1). The level of egg mortality was significantly higher in the group incubated at 20 °C, however, as this treatment only resulted in 44.7 % mortality it did not justify further investigation. In addition, the visual assessments revealed that the level of plant damage in all the tests was not consistently reduced following pre-treatment incubation at one particular temperature. Susceptibility of Liriomyza huidobrensis pupae to hot water treatments Unlike the egg stage, L. huidobrensis pupae are not attached to the plant and therefore are not reliant on the plant material also surviving the treatment. Generally, pupal mortality increased with temperature and the duration of the treatment (Tables 3 and 4). At the higher temperatures, the duration of the treatment required to achieve total mortality was relatively short. For example, at the relatively high temperatures of 45 and 46 °C, treatments lasting 20 and 10 min, respectively, were sufficient to achieve total mortality, whereas a decrease in temperature to 43 required an extended treatment of 45 min. Regression of exposure time (min) and percentage mortality for treatments in water at 43.5 and 45°C estimated that (probit9) 99.9968 % mortality would be achieved after 52.5 and 18.8 min at each of the temperatures. The pre-treatment incubation temperatures of 5 and 20 °C did not consistently alter the susceptibility of the pupae to the hot water treatments (Table 5). In three comparative tests, pupal mortality differed significantly with lower mortality recorded in the groups incubated at 5 °C (tests 2 and 3) and, higher mortality was recorded in test 4. Phytotoxicity Quantitative assessments of chrysanthemum growth were conducted 28 days after hot water treatments and values for treated plants were compared to the control group. The incubation of plants at 20 °C for 24 h followed by a 10 min treatment in water at 43.5 °C did not significantly effect stem height (F = 3.62, d.f. = 1, 10, P > 0.01), shoot dry weight (F = 0.33, d.f. = 1,10, P > 0.01) or root dry weight (F = 3.08, d.f. =1,10, P > 0.01) compared to the control (Fig. 1a-c). Similarly, incubation at 5 °C instead of 20 °C followed by the same hot water treatment did not have an effect (stem height: F = 2.5, d.f.= 1,10, P > 0.05; shoot dry weight: F = 0.44, d.f. = 1, 10, P > 0.05; root dry weight: F = 2.82, d.f. = 1,10, P > 0.05). Doubling the treatment time to 20 min following incubation at 20 °C caused a significant decrease in root dry weight (F = 14.68, d.f. = 1, 10, P < 0.01) and stem height (F = 19.17, d.f. = 1,10, P < 0.01) however, shoot dry weight was not affected (F = 9.56, d.f. = 1,10, P > 0.01). The same regime with pre-treatment incubation at 5 °C also caused a significant reduction in root dry weight (F = 25.71, d.f. = 1, 10, P < 0.05), however, both shoot dry weight and height were not significantly affected (P > 0.05). The seven min treatment at 45 °C following incubation at 20 °C did not significantly reduce shoot and root dry weights (shoot dry weight: F = 0.82, d.f. = 1,10, P > 0.05; root dry weight: F = 2.49, d.f. = 1,10, P > 0.05) however, extending the treatment duration to 10 min resulted in significantly reduced shoot and root dry weights (shoot dry weight: F = 13.93, d.f. = 1,10, P < 0.05; root dry weight: F = 38.81, d.f. = 1,10, P <0.05) (Fig. 1 b and c). Stem heights in both these treatments were not significantly affected, but this may be attributable to the large standard error for height in the control group (7 min at 45, stem height: F = 0.44, d.f. = 1,10, P > 0.05; 10 min at 45 °C, stem height: F= 1.48, d.f. = 1,10, P > 0.05). Decreasing the pre-treatment incubation temperature to 5 °C resulted in significant reductions in all three aspects of monitored growth (45 °C for 7 min, stem height: F = 13.74, d.f.=1,10, P < 0.05; shoot dry weight: F = 27.16, d.f. = 1,10, P < 0.05; root dry weight: F= 13.92, d.f. = 1,10, P < 0.05; 45 °C for 10 min, stem height: F = 81.05, d.f. = 1,10, P <0.05; shoot dry weight: F = 60.14, d.f. = 1,10, P < 0.05; root dry weight: F = 67.04, d.f. = 1,10, P < 0.05). CONCLUSION The duration of treatments in water heated to temperatures between 43 and 45 °C is critical in terms of the mortality of specific life stages of L. huidobrensis and the associated level of damage caused to the host plant, chrysanthemum. The egg stage of L. huidobrensis was highly susceptible to the treatments and slight increases in water temperature above 43 °C greatly reduced the exposure time necessary to achieve high levels of mortality. However, total egg mortality was only recorded following treatments which proved to be very damaging to the host plant. Total pupal mortality was recorded after treatments of 45 min at 43.5 °C, 20 min at 45 °C and 5 min emersion in water at 50 °C. Regression equations were determined for pupae treated at the two temperatures 43.5 and 45 °C and used to estimate the duration of exposure to achieve total mortality 52.5 and 18.6 min, respectively. These values within the region of probit estimates have been reported for immature stages of another dipteran species, Bactocera tryoin when immersed in hot water (Heard et al., 1991). However, treatments, which killed all pupae, proved to be very damaging to chrysanthemum foliage when tested against the egg stage. The incubation of the insect stages at either 5 or 20 °C prior to a hot water treatment did not consistently alter the susceptibility of the egg or pupal stages of the pest to the hot water treatments. Incubation at higher temperatures (28-35 °C) was shown to increase the ability of the light brown apple moth, Epiphyas postrittana, to tolerate exposure to hot water at 43 °C (Lester and Greenwood, 1997). Storage at 20 °C as in the current study did not elicit such a response. None of the tests obtained very high levels of egg mortality without damaging the foliage to warrant the further development of these treatments for the disinfestations of plant material harbouring quarantine insect species. Additionally, the treatments which produced high pupal mortality resulted in unacceptable levels of visual damage to the chrysanthemum foliage. The quantitative assessments of plant growth did not identify any treatment as stimulating plant shoot and root development. Visual assessments of plant damage are of primary importance when developing treatments and chrysanthemum proved very susceptible to damage. The quantitative assessments demonstrated that subsequent growth by the rooted plants was impaired by the treatments and no significant increases in plant development were detected. At 49.4 - 49.8 °C, an exposure of only 5 min caused all leaves to be damaged. The identification of a treatment which reliably attains very high levels of mortality to both egg and pupal stages without causing unacceptable levels of damage to chrysanthemum plant material was not possible. However, other plant types are likely to be more tolerant of the treatments which caused high levels of pest mortality and therefore screening of other relevant commodities could identify useful treatments for the disinfestations of particular commodities moving within the international plant trade. ACKNOWLEDGEMENTS This work was funded by the European Union, Project number FAIR CT984259. The authors thank Mr Richard Natt (CSL Horticultural Manager) and his team for the provision of plant material. Liriomyza huidobrensis were held under quarantine license number: PHL 251B/5328 (02/2006) Amended (04/2006). REFERENCES
© IRSEN, CEERS, IAU The following images related to this document are available:Photo images[st09017t2.jpg] [st09017t5.jpg] [st09017t3.jpg] [st09017t1.jpg] [st09017f1.jpg] [st09017t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}