 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
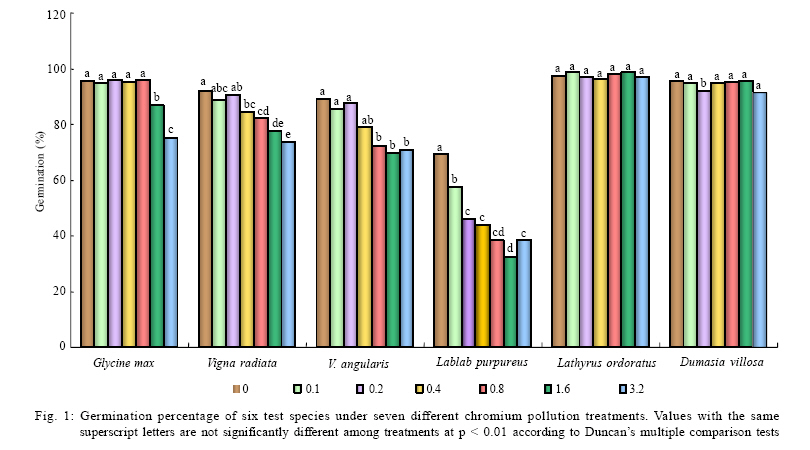
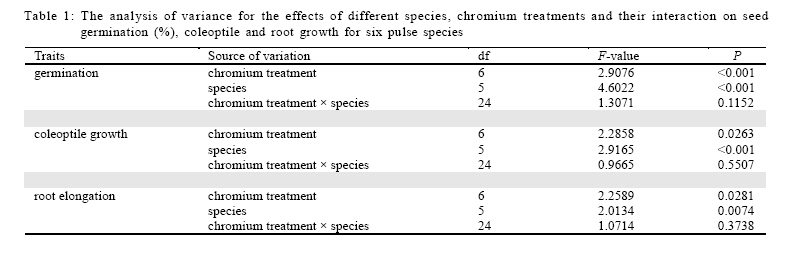
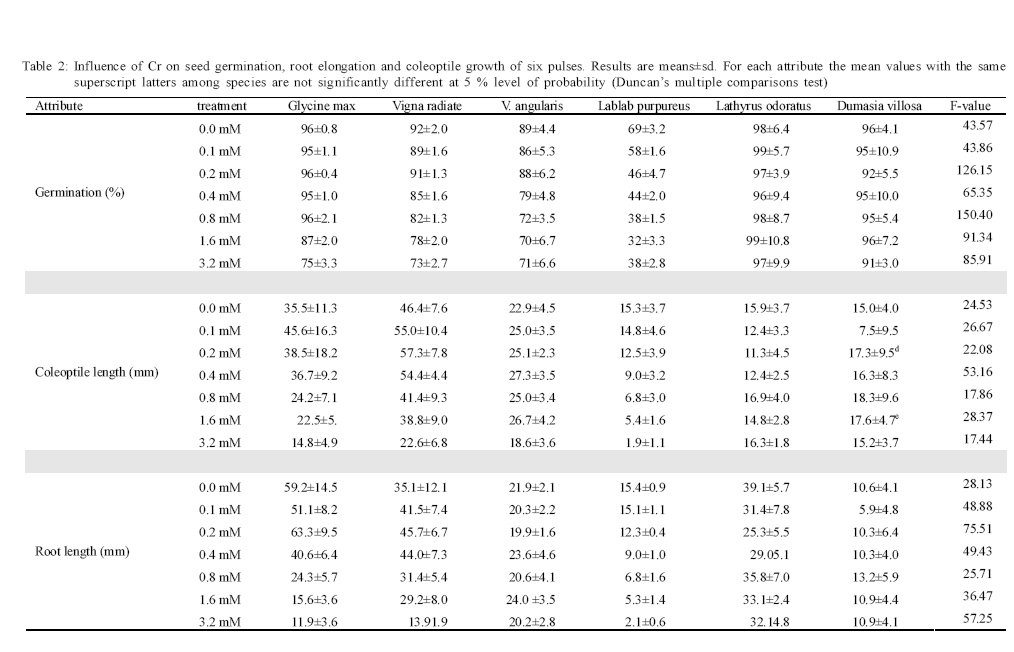
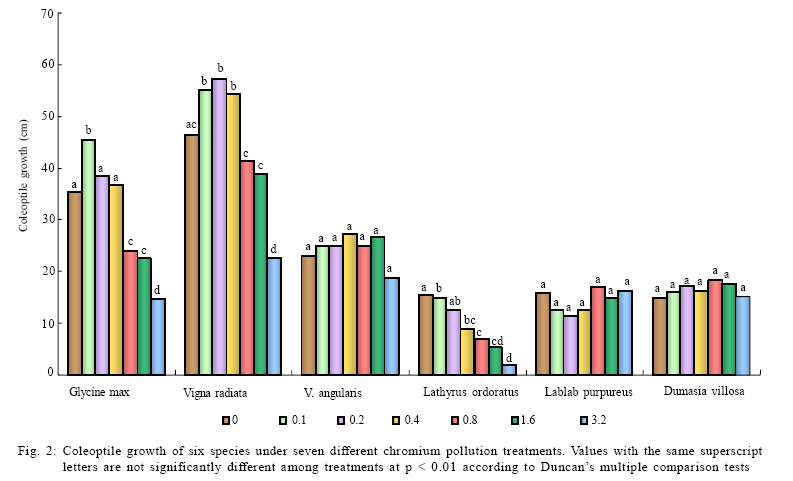
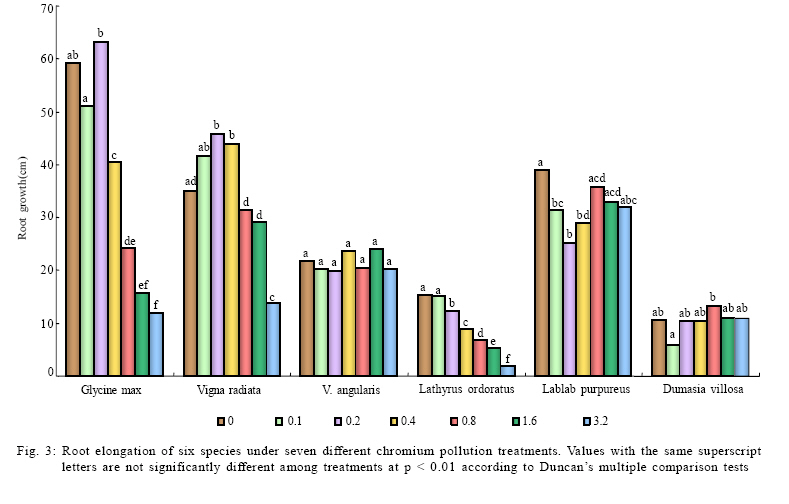
International Journal of Environment Science and Technology, Vol. 6, No. 4, Autumn, 2009, pp. 571-578 Effects of chromium on seed germination, root elongation and coleoptilegrowth in six pulses *R. Jun; T. Ling; Z. Guanghua School of Environmental and Municipal Engineering and Institute of Environmental Ecology, Lanzhou Jiaotong, University, Lanzhou, Lanzhou University, Lanzhou 730070, Gansu, P. R. China *Corresponding Author Email: renjun@mail.lzjtu.cn Tel.: +86931 4938 030; Fax: +86931 7667 661 Received 14 November 2008; revised 16 April 2009; accepted 5 August 2009 Code Number: st09057 ABSTRACT The ecotoxicological effects of Cr2+ on germination and early seedling growth of six pulses were investigated. Seeds of these plants were exposed to seven different concentrations of Cr (0-3.2 mM). The results indicated that root elongation and coleoptile growth of six pulse plants were more sensitive than seed germination for measurement of the toxic of Cr2+ pollutions. Different species show different levels of tolerance to Cr2+ pollution. Lablab purpureus and Glycine max are the most sensitive to Cr2+, their germination percentage, root and coleoptile length were significantly lower than other tested species, by contrast, Lathyrus odoratus and Dumasia villosa are the most resist species, their germination and seedling growth almost were not influenced by Cr2+ pollution significantly comparing the control. There were significantly negatively correlations between seedling growth and increasing concentration of Cr2+ for Glycine max, Vigna radiate and Lablab purpureus. The significantly negative correlations between germination and increasing concentration of Cr2+ were shown for Glycine max, Vigna radiate and V. angularis. Keywords: Chromium; Coleoptile growth; Germination; Pulse; Root elongation Introduction Chromium is important for metallurgical industry. Its salts are used in many industrial processes and product such as, steel production, electroplating, leather tanning, metal finishing, metal corrosion inhibition, textile paints and pigment manufacture, catalysts application, drilling muds, fungicides and nuclear weapons production and hence are present in effluents of those industries and in municipal sewage (Wong et al., 2001; Zayed and Terry, 2003; Nath et al., 2005; Babel and Opiso, 2007; Venkateswaran et al., 2007). Chromium is the second most common heavy-metal contaminant in groundwater, soil and sediments (Wong et al., 2001; Srivastava and Thakur, 2006; Shrestha et al., 2007; Kar et al., 2008; Ogundiran and Afolabi, 2008 ) and although in higher concentration, chromium ions which are more or less insoluble in the soils, can create potentially toxic environments for plant growth (Hasnain and Sabri, 1997; Wong et al., 2001; Srivastava and Thakur, 2006). Chromium phytotoxicity has been considered to be inhibitory for plant growth, its presence in excess amount within the plant can cause stunted shoot growing, root developing poorly (Bishnoi et al., 1993; Citterio et al., 2003; Dube et al., 2003; Faisal and Hasnaim, 2005; Gbaruko and Friday, 2007). Chromium toxicity in plants also leads to leaves chlorosis, tissue necrosis, decreases enzyme activity, causes membrane damage, diminished photosynthesis and changing of chloroplast (Jain et al., 2000; Parmar et al., 2002; Du et al., 2003; Dube et al., 2003; Zayed and Terry, 2003; Scoccianti et al., 2006). The toxicity of chromium and chromium-content in plants, however, is species specific. Much research has been conducted to determine the toxic effects of chromium on different plant species (Hasnain and Sabri, 1997; Jain et al., 2000; Ren and Gao, 2000; Peralta et al., 2001; Zeid, 2001; Manjappa et al., 2002; Dube et al., 2003; Zhou and Li, 2003; 2004; Faisal and Hasnaim, 2005; Gardea-Torresdey et al., 2005; Jamal et al., 2006; Li and Yang, 2006; Karbassi et al., 2008). However, little information is available on the ecological effects and toxicity of chromium on 6 pulse plants (Leguminosae) which are widely cultivated around the world as crop species. These pulse plants can uptake heavy metal from polluted soil (Bishnoi et al., 1993). The heavy metals accumulated by pulse plants can threat health of human by entering the food chain as food or fodder. The present study was undertaken with a view to find out the toxic effect of chromium on seed germination, root elongation and coleoptile growth of six pulse crops. With the specific aim of determining: 1) What degree chromium inhibits the six pulse crops' seed germination and seedling growth? 2) Which pulses are most sensitive to chromium? This study was carried out on June of 2008 in Institute of Environmental Ecology, Lanzhou Jiaotong University. Materials and methods The effects of several doses of K2Cr2O7 (0, 0.1, 0.2, 0.4, 0.8, 1.6 and 3.2 mM) were studied on six pulse crops. Seeds of soybean (Glycine max (Linn.)), mung bean (Vigna radiata (Linn.) Wilczek.), hyacinth (Lablab purpureus (Linn.) Sweet), sweet pea (Lathyrus odoratus Linn.), adzuki bean (Vigna angularis (Willd.) Ohwietohashi.) and black bean (Dumasia villosa DC.) were offered by Gansu Academy of Agricultural Sciences of China. Seeds were surface sterilized in 0.5 % sodium hypochlorite solution for 20 min and washed thoroughly with distilled water. The seeds were germinated in Petri dishes (diameter=150 mm) with double layer of filter paper soaked in distilled water (control) and 0.1-3.2 mM CrCl2 solutions. The seeds were set under a photoperiod of 12 h and 30 ± 1/25 ± 1 ºC day/night temperature. The seedling was harvested after 96 h and the germination rate was recorded. A 1-mm radical emergence from seeds was considered seed germination. The root and coleoptile length of 10 germinated seeds were measured at 96 h under each treatment. Data are the results from six separate analyses with 50 seeds in each Petri. Statistical analysis was performed based on STATISTICA (Statsoft, 1993). The data were analyzed through one-way analysis of variance (ANOVA) to determine the effect of treatments and least significant difference (LSD) test were performed to determine the statistical significance of the differences between means of treatments. Results and discussion Germination under chromium stress The germination of pulse seeds occurred at all chromium treatments from 0.0 mM to 3.2 mM for each species (Fig. 1). A two-way ANOVA showed that seed germination was significantly affected by different species (F5 = 4.6022, P < 0.001) and chromium treatment (F6 =2.9076, P < 0.001) and the absent of a significant interaction demonstrates that the response of all species to chromium concentration was similar, although percentage of germination varies among the species (Table 1). The germination was significantly different among chromium treatment and control in 5 pulse species (one-way ANOVA: F6,35 = 108.82, p < 0.001 for Glycine max; F6,35 = 9.62, p < 0.001 for Vigna ratiata; F6,35 = 6.20, p < 0.01 for V. angularis; F6,35 = 18.98, p < 0.001 for Lablab purpureus; F6,35 = 5.78, p < 0.01 for Dumasia villosa). Only in the case of Lathyrus odoratus, its germination was not significantly effected by chromium treatment (F6,35 = 0.60, p = 0.73) and it performed higher germination percentage and stronger tolerance to chromium. Compared with the control and lower concentration chromium treatments (0.1 and 0.2 mM), higher concentration chromium treatments (1.6 and 3.2 mM) significantly (p < 0.001) inhabited germination for Glycine max, Vigna ratiata, V. angularis and Lablab purpureus (Fig. 1). The germination was significantly different (p < 0.001) among species at six kinds of chromium treatments and one control (Table 2). At all chromium treatments and control, the germination percentages of Lablab purpureus was significantly lower than other 5 species (p <0.001). At control and 0.1 mM chromium treatment, the germination percentage of V. angularis was significantly higher than Lablab purpureus and lower than other 4 species (p < 0.001). At control and 0.2, 0.4 and 0.8 mM chromium treatment, the germination percentages of Glycine max, Lathyrus odoatus and Dumasia villosa did not performed significantly difference, and were significantly higher than Vigna ratiata, V. angularis and Lablab purpureus (p < 0.001), there was not significant difference in germination percentages between Vigna ratiata and V. angularis. At higher concentration chromium treatment (1.6 and 3.2 mM), the germination percentages of Lathyrus odoatus and Dumasia villosa did not performed significantly difference and were significantly higher than other 4 species (p < 0.001) and there was not significant difference in germination percentages between Glycine max and Vigna ratiata (Table 2). There was a significant negative correlation between the mean percent germination and chromium concentration for Glycine max, Vigna ratiata and V. angularis (p < 0.001) (Table 3). Coleoptile growth under chromium stress A two-way ANOVA showed that coleoptile growth was significantly affected by different species (F5 =2.9165, P < 0.001) and chromium treatment (F6 = 2.2858, P = 0.0263), but not by the interaction between species and chromium treatment (Table 1). The coleoptile length was significantly different among chromium treatments in 3 pulse species (one-way ANOVA: F6,35 = 5.37, p < 0.001 for Glycine max; F6,35 = 13.68, p < 0.001 for Vigna ratiata; F6,35 = 14.68,p < 0.001 for Lablab purpureus). For the other 3 pulse species, the coleoptile growth was not significantly different among chromium treatments (F6,35 = 1.74, p = 0.140 for V. angularis; F6,35 = 2.64, p = 0.052 for Lathyrus odoratus; F6,35 = 1.44, p=0.228 for Dumasia villosa). Compared with the control and shorter concentration chromium treatments (0.1 and 0.2 mM), higher concentration chromium treatments (1.6 and 3.2 mM) significantly inhabited coleoptile growth for Glycine max, Vigna ratiata and Lablab purpureus. Lablab purpureus performed shorter coleoptile growth at all chromium treatments (p < 0.001). Comparing with control, for Glycine max, 0.1 mM chromium treatment significantly promoted the coleoptile growth (p < 0.001), 0.2 mM and 0.4 mM chromium treatments had no significant influence on coleoptile growth and for Vigna ratiata, lower chromium treatments (0.1, 0.2 and 0.4 mM) significantly promoted the coleoptile growth (p < 0.001) (Fig. 2). The coleoptile growth was significantly different among species at six kinds of chromium treatments and one control (p < 0.001) (Table 2). The coleoptile growth of Lablab purpureus decreased with addition of chromium concentration (Fig. 2). At control treatment, Glycine max and Vigna ratiat did not performed significant difference in the coleoptile growth, but their coleoptile growth was significantly higher than other 4 species (p < 0.001). At higher chromium treatments (0.4, 0.8, 1.6 and 3.2 mM), Glycine max and Vigna ratiat performed significant difference (p < 0.001) and their coleoptile growth was both significantly higher than other 4 species (p < 0.001) and the coleoptile growth of Lablab purpureus was significantly shorter than other 5 species (p < 0.001). At 1.6 and 3.2 mM chromium treatment, the coleoptile growth of Vigna ratiat was significantly higher than 5 species and Lablab purpureus was significantly shorter than other 5 species (p < 0.001) (Table 2). There was a significant negative correlation between the mean coleoptile length and chromium concentration for three species (Glycine max: y=38.58-0.89x, r2=0.79, F1,5=18.26, p=0.0079; Vigna ratiata: y=53.93-0.92x, r2=0.85, F1,5 = 29.28, p = 0.0029; Lablab purpureus: y = 12.94-0.90x, r2 = 0.81, F1,5 = 21.06, p = 0.0059). Root elongation under chromium stress A two-way ANOVA showed that root elongation was significantly affected by different species (F5 = 2.0134, P = 0.0074) and chromium treatment (F6 = 2.2589, P = 0.0281), but not by the interaction between species and chromium treatment (Table 1). The root length was significantly different among chromium treatments in five pulse species (one-way ANOVA: F6,35 = 39.47, p < 0.001 for Glycine max; F6,35 = 12.81, p < 0.001 for Vigna ratiata; F6,35 = 3.78, p=0.005 for Dumasia villosa; F6,35 = 136.51, p < 0.001 for Lablab purpureus; F6,35 = 3.69, p=0.006 for Lathyrus odoratus). For V. angularis, the root elongation was not significantly different among chromium treatments. Compared with the control and shorter concentration treatments (0.1, 0.2 and 0.4 mM), highest concentration chromium treatments (0.8, 1.6 and 3.2 mM) significantly inhabited root elongation for Glycine max (p < 0.001). For Vigna ratiata, the root length dramatically decreased at 3.2 mM chromium treatment and was significantly shorter than other species (p < 0.001). The root elongation of Lablab purpureus performed decline with increase of chromium concentration. The root elongation of Dumasia villosa was not effected by chromium treatments comparing with the control (Fig. 3). There was significantly difference of root elongation among species (p < 0.001) at six kinds of chromium treatments and one control (Table 2). At control treatment and lower chromium concentration treatments (0.1 and 0.2 mM), the root length of Glycine max was significantly longer than other 5 species (p < 0.001); the root lengths of Lablab purpureus and Dumasia villosa were significantly shorter than other 4 species (p < 0.001); the root lengths of Vigna ratiata was significantly shorter than Glycine max and longer than other 4 species (p < 0.001). At higher chromium concentration treatments (1.6 and 3.2 mM), there was not significant difference between Glycine max and Dumasia villosa. The root length of Lablab purpureus was significantly shorter than other 5 species (Table 2). There was a significant negative correlation between the mean coleoptile length and chromium concentration for three species (Glycine max: y = 51.85-15.41x, r2 = 0.85, F1,5 = 12.69, p = 0.0162; Vigna ratiata: y = 42.25-8.71x, r2 = 0.83, F1,5 = 24.92, p = 0.0041; Lablab purpureus: y = 12.96-3.93x, r2 = 0.80, F1,5 = 19.42, p = 0.0070). Chromium is very important for the normal growth of plants, although stimulating effects to the additions of Cr on plant growth have been observed by several researchers (Lintschinger et al., 1997), but excessive amount can leads to toxicity. The Cr2+ showed very strong inhibits effect to germination, root and coleoptile growth of the test species, especially at high chromium concentrations and the root and shoot growth were much more sensitive than the germination by this study. High levels of Cr2+ supply can inhibit seed germination and subsequent seedling growth (Zayed and Terry, 2003). In another study, Cr (VI) concentrations up to 2 mM supplied as K2Cr2O7 did not affect germination of pea seeds significantly, this was similar results for Vigna angularis and Lathyrus ordoratus by this study. However, growth of root and shoot was suppressed at chromium concentrations of 0.5mM. The seed yield was reduced by 75-80 % and even at 0.1 mM Cr seed yield was reduced by about 70 % At Cr(VI) concentration of 0.2 mM (Bishnoi et al., 1993). Munzuroglu and Geckil (2002) demonstrate that inhibition of root elongation is considered to be the first evident effect metal toxicity in plants, may be it's because of the seed coat is able to reduce the amount of Cr2+ entering the seed, but after the seed germination, chromium is considered strongly toxic because chromium compound in the soil are more or less insoluble as the metal ions are tightly bound to humus and clay particles. Shoot and root after germination have no barrier to protect. Some other studies have the same conclusion (Rout et al., 2000). Conclusion This study concluded that seed germination of all the test species were not so sensitive like root and coleoptile elongation to Cr2+, root and coleoptile growth of the test species were more accurate to measure the toxic of Cr2+ pollutions and different species show different levels of tolerance to Cr2+. Lablab purpureus and Glycine max are the most sensitive to Cr2+. By contrast, Lathyrus odoratus and Dumasia villosa are the most resist species. REFERENCES
© IRSEN, CEERS, IAU The following images related to this document are available:Photo images[st09063f2.jpg] [st09063t2.jpg] [st09063t1.jpg] [st09063f3.jpg] [st09063t4.jpg] [st09063f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}