 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
International Journal of Environment Science and Technology, Vol. 8, No. 1, 2010, pp. 45-56 Biodegradation of cypermethrin by a newly isolated actinomycetes HU-S-01 from wastewater sludge 1Q. S. Lin; 2S.H. Chen; 1*M. Y. Hu; 1M. R. Ul Haq; 1L. Yang; 1H. Li 1Key Laboratory of Natural Pesticide and Chemical Biology, Ministry
of Education, South China Agricultural
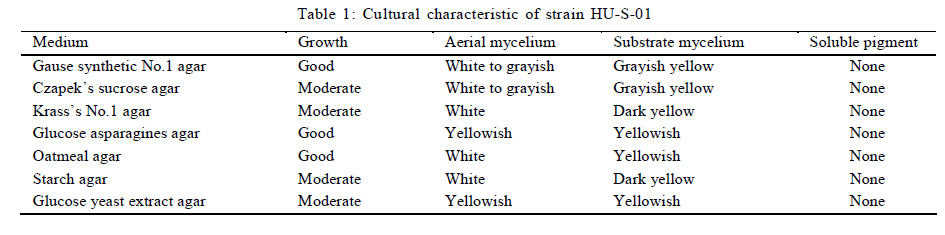
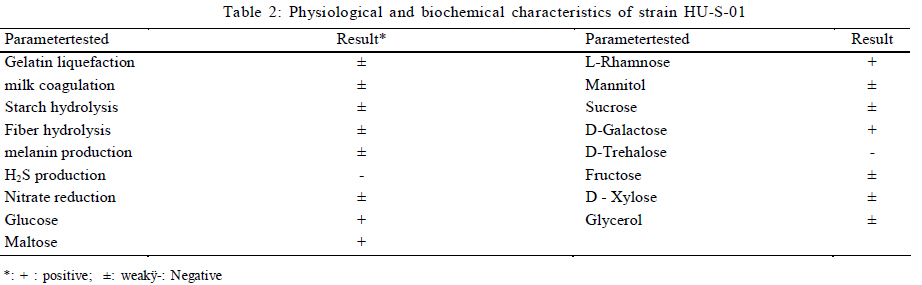
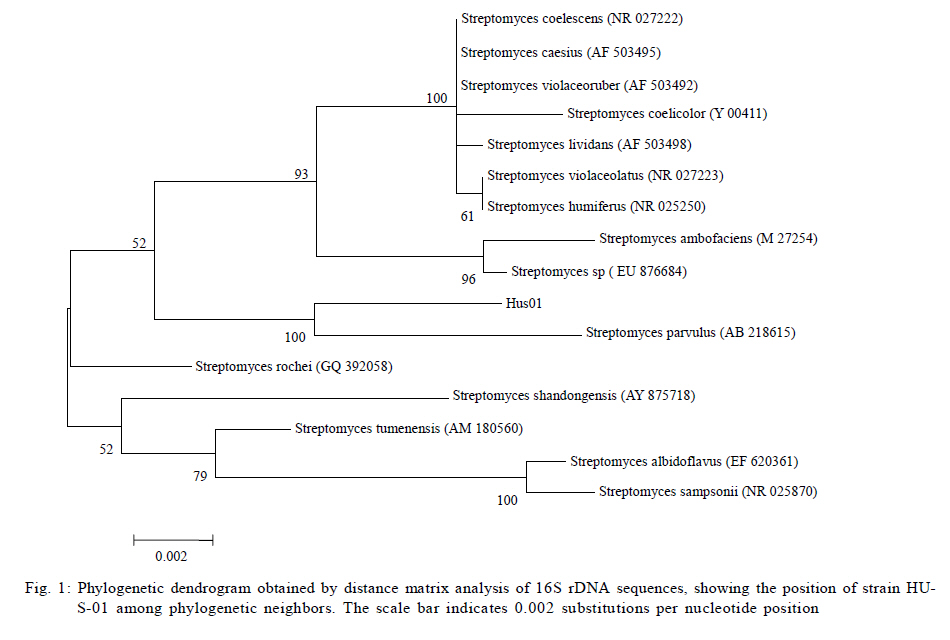
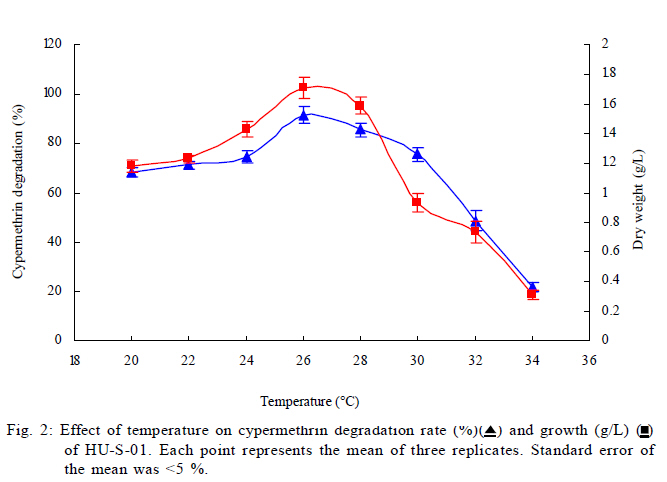
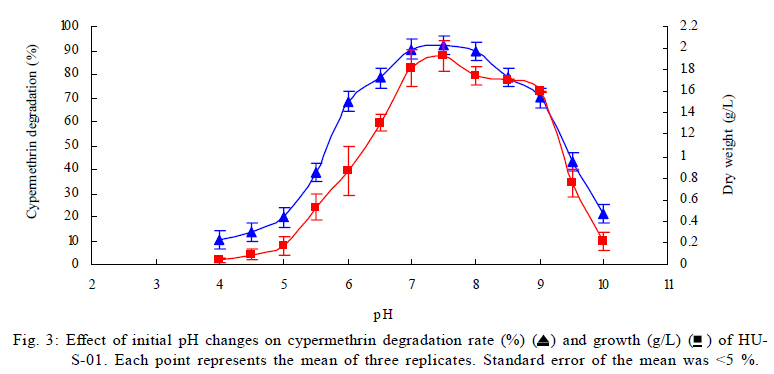
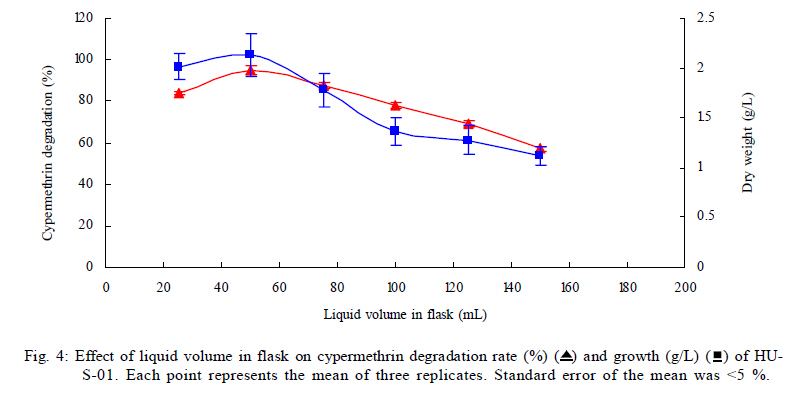
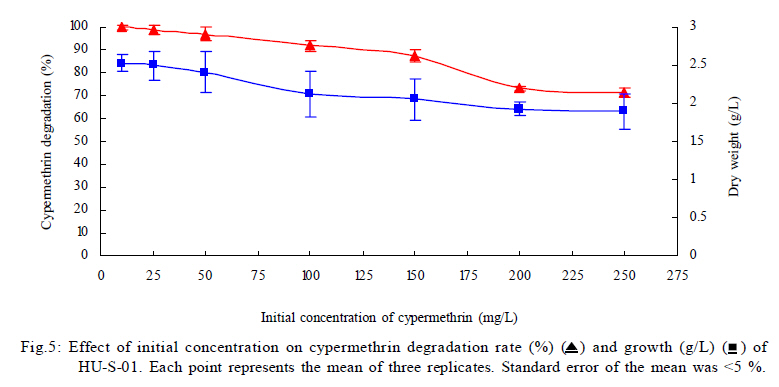
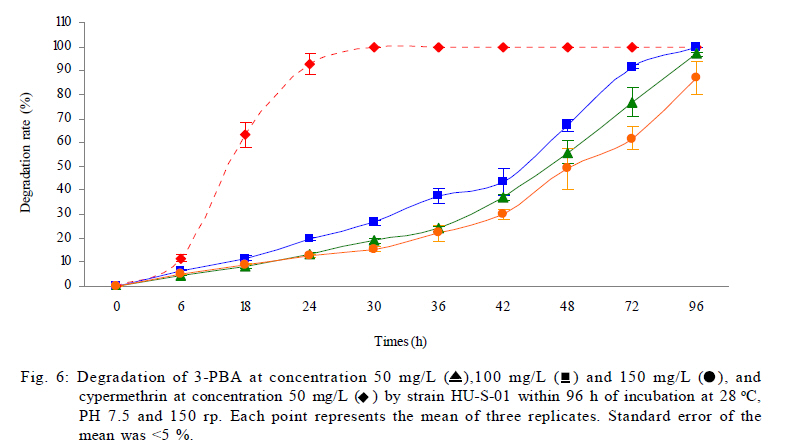
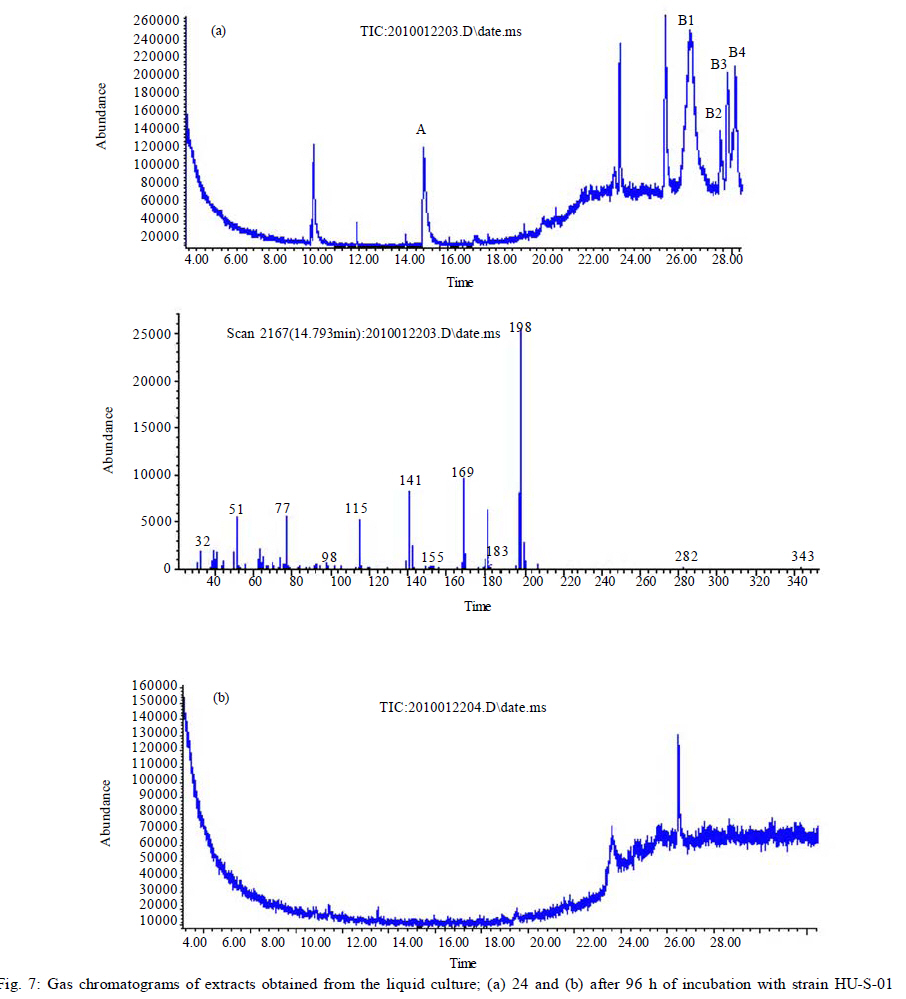
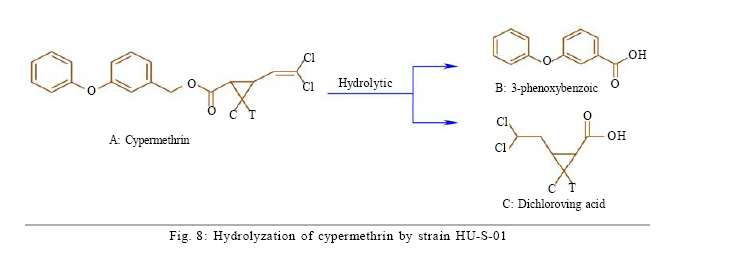
University, Guangzhou, 510640, P. R. China *Corresponding Author Email: humy@scau.edu.cn Tel.: +86 020 85280308; Fax: +86 020 85280308 Received 29 March 2010; revised 5 June 2010; accepted 18 July 2010 Code Number: st11004 ABSTRACT: A new cypermethrin degrading strain was isolated from wastewater sludge using enrichment technique. On the basis of morphological, physiological, biochemical characteristics tests and 16S rDNA sequence analysis, the strain was determined to be a Streptomyces species, probably a strain of Streptomyces parvulus, so it was designated as Streptomyces sp. HU-S-01. The strain Streptomyces sp.HU-S-01 is aerobic and optimum growth temperature for the strain was found to be 26-28 oC with initial pH range 6.0-9.0 and pH 7.5 was found to be the optimum. This strain can also completely degrade 3-phenoxybenzoic acid within 96 h at the concentration of 50 mg/L. The kinetic constants Vmax, Km, Kcat and Kcat/ Km of enzyme for cypermethrin were 1.236 μmol/min, 6.418 μmol/mL, 13.493 min and 2.102 mL/mol.min, respectively. The degradation products of cypermethrin were identified using gas chromatograph-mass spectrometric (GC-MS) analysis. The degradation pathway followed by HU-S-01 involves oxidative as well as hydrolyzing. Biodegradation ability of strain Streptomyces sp. HU-S-01 without toxic byproducts reveals its potential for further study as a biological agent for the remediation of soil, water or crops, contaminated with cypermethrin. Keywords: Biodegradation; Metabolites; Pyrethroid insecticides; Streptomyces sp.; 3-phenoxybenzoic acid INTRODUCTION The synthetic pyrethroid insecticides are analogs of naturally occurring pyrethrins of botanical origin (Elliott, 1980). This group has become a major class of insecticides, used against pests as a replacement for more toxic organophosphorus pesticides (Katsuda, 1999). Cypermethrin [(+/-)-a-cyano-3-phenoxybenzyl (+/-)-cis,trans-3 (2,2-dichlorovinyl)-2,2-dimethylcy clopropanecarboxy-late] as a synthetic pyrethroid pesticide, was widely used to control pests of cotton, fruits and vegetables as well as cockroaches, fleas, termites in houses and other buildings (Tallur, et al., 2008). Earlier, the synthetic pyrethroid insecticides were considered as the safest insecticides because of their quick insecticidal capabilities and generally low mammalian toxicity (Dorman and Beasley, 1991). Recently many studies showed a lot of environmental and health problems due to continuous and excessive use of synthetic pyrethroid insecticides (Cuthbertson and Murchie, 2010; Cuthbertson et al., 2010). For example, cypermethrin has an extremely high toxicity to the aquatic environment with concentrations as low as 10 mg/L, destroying aquatic invertebrate life (Pearce, 1997; Virtue and Clayton, 1997; Vinodhini and Narayanan, 2008) and reported to have carcinogenic effects (Cabral and Galendo, 1990; Kakko et al., 2004). Although some scientists did not observe the carcinogenic effects of pyrethroid in the mouse (Cabral et al., 1990) yet Shukla et al., (2002) reported that cypermethrin was found to have carcinogenic activity in both sexes of Swiss albino mice. The carcinogenic potential of pyrethroids has also been discussed in the review by Litchfield (1985) and cypermethrin has been classified as a possible human carcinogen by US Environmental Protection Agency. Thus, an effective and environment friendly treatment method is required to prevent environmental pollution caused by pyrethroid waste. Microbial biodegradation of environmental pollutants is a field of growing importance because of its enormous potential for environmental clean up (Schmid et al., 2001; Dua et al., 2002; Pazos et al., 2003; Nwuche and Ugoji, 2008 and 2010). It is a practical and applicable solution for detoxifying phenols, diesel fuel, azo dyes, anionic surfactants and pesticide in a range of varying conditions whether in fruits, vegetables, river or soil (Grant et al., 2002; Hosseini et al., 2007; Agarry and Solomon, 2008; Babel and Opiso, 2007; Agarry et al.,2009; Kebria et al., 2009; Dhanasekaran et al., 2009; Ghasemi et al., 2010). Many efforts have been undertaken to isolate bacteria, capable of biodegradation of synthetic pyrethroid insecticides and a lot of pyrethroid-degrading microorganisms have been isolated from soil and polluted water (Sakata et al.,1992; Maloeny et al., 1993; Halden et al.,1999; Nirmali et al., 2005; Jilani and Khan, 2006; Tallur et al., 2008; Arulazhagan et al., 2010; Murugesan et al., 2010). Some pyrethroid hydrolase were purified and characterized from fungi, insect and mouse liver (Liang et al., 2005). Genes encoding the pyrethroids-hydrolyzing carboxy-lesterases were also cloned and functionally expressed (Stock et al., 2004; Wu et al., 2006; Wang et al., 2009). With the development of study, microbial degradation has been considerd the most influential and significant cause of pesticides removal. Biodegradation is also considered to be a reliable cost-effective technique for pesticides abatement and a major factor, determining the fate of pyrethroid pesticides in the environment (Horne, 2002). In this study, the isolation and characterization of an actinomycetes strain HU-S-01 from the wastewater sludge has been carried out. The study also reveals its degradation efficiency of cypermethrin and provides useful information for further application of this isolate for bioremediation of cypermethrin contamination on the surface of fruits, vegetables and in the environment. This work was performed in Laboratory of Natural Pesticide and Chemical Biology, Ministry of Education, South China Agricultural University, Guangzhou, P.R. China. from 2008 to 2009. MATERIALS AND METHODS Chemicals and medium Cypermethrin was supplied by Zhongshan Aestar Fine Chemical Inc.Ltd., and 3-phenoxybenzoic acid (3-PBA) was purchased from Sigma-Aldrich, USA. Acetonitrile and methanols were of chromatographic grade while other chemicals were of analytical grade. Czapek-Dox medium was used to enrich the culture and Gause synthetic No.1 media was used to culture the stain HU-S-01. The media were solidiûed with 18 g/L of high-quality agar for cell growth on plate. Enrichment and isolation of cypermethrin-degrading strain HU-S-01 Wastewater sludge was collected from severage treatment pool of Zhongshan Aestar Fine Chemical Inc.Ltd. Wastewater sludge enrichment was performed by placing 5g wastewater sludge sample in a 250 mL Erlenmeyer ûask containing 100 mL of Czapek-Dox medium and 20 mg cypermethrin. After 5 days of incubation on a rotary shaker at 140 rpm and 28 oC, the soil suspension (5 mL) was transferred into another Erlenmeyer ûask containing 100 mL fresh Czapek-Dox medium and 30 mg cypermethrin. These steps were repeated five times and the medium containing 50 mg cypermethrin was achieved at the end. A serial dilution of the enrichment culture was carried out and spread on the Czapek-Dox medium agar plate supplemented with 500 mg/L cypermethrin. After 4-6 days of incubation, Individual colonies were subcultured into nutrient agar plates containing 500 mg/L cypermethrin until pure colony was isolated. The pure isolate with highest cypermethrin degradation efficiency was selected for further study. Morphological, physiological and biochemical characteristics The HU-S-01 was identified by morphological, physiological and biochemical characteristics according to the Classification and Identification of Actinomycetes (Yan, 1992). HU-S-01 was grown on Gause synthetic No.1 agar medium and cultural characteristics such as; shape, margin, elevation, surface appearance of the colonies, colour change in the medium because of pigment formation, assimilation of carbon source and odor of the cultures were noted. Morphological features of the cells and spores were also observed with microscope. 16S rDNA sequence and phylogenetic analysis The genomic DNA was extracted from HU-S-01 using a Microbial DNA isolation kit (Tiangen Biotech (Beijing china) co., LTD.). The 16S rDNA gene fragment was ampliûed from the genomic DNA by PCR using the general-purpose primers of prokaryotic 16S rDNA: upstream primer 5AGAGTTTGATCCTGGCTCAG3', and downstream primer 52AAGGAGGT GATCCAGCCGCA32 (Wang et al., 2009). Each 50 mL PCR micro-tube contained 2 µL purified extracted DNA; 1 µL of upstream primer (10 pmol); 1 mL of downstream primer (10 pmol); 5 mL of 10×ExTap Buffer ; 0.5 mL Ex Tap DNA Polymerase(5 µL); 4 µL of dNTP at 2.5 mM and 36.5 µL of sterile MQ water. DNA ampliûcation was performed in a thermal cycler with the following conditions: denaturation for 3 min at 94 oC, 35 cycles at 94 oC for 30s, 55 oC for 1 min and 72 oC for 1.5 min, with extension of 72 oC for 5 min. BLAST search was carried out at the National Center for Biotechnology Information. Phylogenetic analyses of the sequence data was performed using Mega 4.1 software. Optimum conditions for biodegradation of cypermethrin by HU-S-01 The effect of ventilation volume on biodegradation of cypermethrin was studied. 5 mL spore suspension (6.7×108 CFU/mL) of strain HU-S-01 was added in sterile 250 mL Erlenmeyer ûasks containing 25, 50, 75, 100, 125 and 150 mL Gause Synthetic No.1 media. 50 mg/L cypermethrin after incubation for 72 h at 28 oC and shaken at 120 rpm was added in the flask. Cypermethrin was extracted for HPLC analysis while the dry weight of mycelium was noted after further culturing for 24 h (Liu and Zhong, 2000). Batch tests were performed in triplicate. The effect of biodegradation was also studied at various initial pH (4.0-10.0 at 28 oC) and different temperatures (20-34 oC at pH=7.0). Hu-S-01 cells were also studied in triplicate for 50 mg/L cypermethrin concentration at 28 oC (pH7.0). Extraction of Cypermethrin for HPLC analysis Samples were collected from 50 mL bacteria solution and extracted three times with petroleum ether mixture (100 mL, 75 mL and 50 mL) by vigorous shaking for 15-20 minute in a separatory funnel. The petroleum ether layer was separated and evaporated to dryness at 50 oC using rotary vacuum evaporator. The dried residue was dissolved in 10 mL chromatographic grade acetone. After gently vortexing and filtering through a 0.2 mm membrane filter, an aliquot of 2 mL was used for HPLC analysis. The HPLC conditions were as follows: HP-1100, Chromatographic column: C18 column (Hypersil ODS2 5 mm, 4.6 mm×250 mm), Flow rate:1 mL/min, Column temperature: 28±1 oC, Mobile phase: acetonitrile: water=90:10, UV detectory 235 nm, Injection volume: 10 mL. Each sample was injected 3 times and the means were calculated. Degradation of 3-phenoxybenzoic acid HU-S-01 was grown for 72 h at 28 °C in 250 mL Erlenmeyer ûasks containing 50 mL Gause Synthetic No.1 media. 2.5, 5.0, 7.5 mg 3-PBA was added to the erlenmeyer ûasks. After 6, 12, 18, 24, 30, 36, 42, 48, 72 and 96 h, 3-PBA was extracted with 60 mL methanol/methylene dichloride (21) +120 µL hydrochloric acid three times. The methylene dichloride layer was separated and evaporated to dryness at 80 °C using rotary vacuum evaporator. The dried residue was dissolved in 10 mL mobile phase of HPLC. After gently vortexing and filtering through a 0.2 mm membrane filter, an aliquot of 2 mL was used for HPLC analysis (Xie et al., 2007). Enzyme activities HU-S-01 was incubated in 250 mL Gause synthetic No.1 media at 28 oC for 96 h and centrifuged at 4000 g for 20 minutes. Mycelium was grinded and the cells were suspended in 50 mmol/L potassium phosphate buffer (pH 7.5), at 4 oC and given 120 strokes each of 2 s with 5 s intervals, by sonication (BILON88-a!, Shanghai Bilon Instruments Co., LTD) at 300 W. Cell debris was removed by centrifugation at 12000 g for 20 minutes. Cell-free supernatant was used for enzyme assays and protein concentration was examined using the Bradford method (Bradford, 1976) with bovine serum as a protein standard. Enzymatic activity was measured spectro-photometrically at 28 °C and pH 7.5. One unit of enzyme activity for cypermethrin and 3-PBA was defined as the amount of enzyme catalyzing the substrates at 1 ìmol/min. Cypermethrin and 3-PBA were detected at 235nm and 210 nm respectively, by the Ultraviolet Spectrophotomete (Shimazu UV mini1240). The kinetic constants of cell extract for cypermethrin and 3-PBA (10 mg/L each) such as maximum reaction rate (Vmax), michaelis constant (Km), catalytic constant (Kcat) and specificity constant (Kcat/ Km) were estimated by Lineweaver-Burk plot. Detection of cypermethrin metabolites Culture filtrate of medium containing cypermethrin used for growth of HU-S-01 was extracted with dichloromethane. The dichloromethane extract was evaporated and the residue dissolved in acetone. The extracts were analyzed by GC-MS (Agilent, USA). The GCMS analyses were performed in electron ionization (EI) mode (70 eV) with a Agilent gas chromatograph, equipped with an MS detector. A HP-1701 capillary column (30 m length × 0.25 mm id × 0.25 ìm film thickness) was used with a temperature program of: 80 °C for 1 min; increased to 200 °C at 8 °C/min and held for 2 min; and finally increased to 260 °C at 15 °C/min and held at 260 °C for 10 min. Nitrogen was used as the carrier gas at a constant flow of 1.0 ml/min. The samples were analyzed in split mode (1:20) at an injection temperature of 260 °C and an EI source temperature of 230 °C and scanned in the mass range from 50 m/z to 450 m/z. RESULT AND DISCUSSION Enrichment and isolation of cypermethrin-degrading strain HU-S-01 In the present study, 26 different types of bacterial colony isolates were obtained after enrichment with cypermethrin from wastewater sludge collected from severage treatment pool of Zhongshan Aestar Fine Chemical Inc.Ltd. Out of 26 bacterial colonies, only one bacterial strain, named HU-S-01, expressed a comparatively good growth rate and greater cypermethrin degradation efficiency. It degraded about 92.1 % substrate within 24 h and 100 % after 30 h at the concentration of 50 mg/L (cypermethrin) in the liquid minimal medium. Therefore, HU-S-01 was chosen for further study to determine the optimum conditions for cypermethrin biodegradation. Almost all the pyrethroid-degrading strains were isolated from the samples collected from fields, wastewater sludge and sheep dipping facility etc which were seriously contaminated by pyrethroid insecticides and other chemical. For example, the Sphingobium sp. JZ-1 was isolated from a pyrethroid-manufacturing wastewater treatment facility (Wang et al., 2009), Aspergillus nige ZD11 and micrococcus sp. were isolated from the soils where pesticide were heavily used (Liang et al., 2005; Tallur et al., 2008), Acidomonas sp. was isolated from automobile garages within city limits (Mandar et al., 2005), Pseudomonas fluorescens and Serratia plymuthica lived in sheep dipping facility and garden soil (Grant et al., 2002). In these environments, the lives faced high selection pressure from the contaminants, so some living styles evolved ability of detoxification to survive in harsh environments. Some isolates can mineralize the contaminants and be able to utilize them as a sole carbon and energy source. Such as Aspergillus niger ZD11, Micrococcus sp. and Acidomonas sp. can use some pyrethroid insecticide as sole source of carbon and energy. Acinetobacter baumannii CA2, Pseudomonas putida CA16 and Klebsiella sp. CA17 can use 4-Chloroaniline as a sole carbon and nitrogen source without stimulation or co-cultivation with aniline or another co-substrate (Alisa and Wansiri, 2007), Mycobacterium sp. effectively utilized 1, 4-dioxane as a sole carbon and energy source (Kim et al., 2009), Acinetobacter sp. ISTPCP-3 can grow with pentachlorophenol as sole source of carbon and energy (Ashwani et al., 2009). Some other isolates cannot mineralize the contaminants but depend on the availability of other carbon sources and nutrients. For example, Pseudomonas sp. is a synthetic pyrethroid-degrading bacteria that can degrade synthetic pyrethroids in sheep dips, but cannot utilize them as sole carbon and energy source (Grant et al., 2002). In present study, though the strain HU-S-01 was able to degrade cypermethrin and 3-PBA efficiently but like the Pseudomonas sp. cannot utilize them as sole carbon and energy source and depends on the availability of other carbon sources and nutrients such as starch or sucrose to grow and incubate. Identification of HU-S-01 HU-S-01 is aerobic and Gram-positive bacteria. The surface of spore was smooth and white while the color of aerial mycelium and substrate mycelium varied on different media. It developed white to grayish aerial hyphae and grayish yellow substrate mycelium on gause synthetic No.1 media. As shown in Table 1, HU-S-01 developed well on various mediums including Gause synthetic No.1 agar, Glucose asparagines agar and oatmeal agar. It showed moderate growth on Krass's No.1 agar, starch agar, Glucose yeast extract agar and Czapek's sucrose agar and no soluble pigments were produced on any medium. As shown in Table 2, HU-S-01 utilized L-Rhamnose, D-Galactose, Glucose, Maltose positively and utilized Mannitol, Sucrose, Fructose, D-Xylose, Glycerol weakly, but did not utilize D-Trehalose. Acid was not formed from these carbon sources tested. Tests for starch hydrolysis, fiber hydrolysis, melanin production, gelatin liquefaction, milk coagulation, nitrate reduction were positive and H2S production was negative. The sequence of the16S rDNA gene from HU-S-01 showed 100 % similarity to the sequences of the 16S rDNA genes from Streptomyces parvulus (AB 218615) and Streptomyces sp. (EU 876684). The phylogenetic analysis (Fig. 1) based on the 16S rDNA analysis using MEGA 4.1 indicated that the isolate is closely similar to S. parvulus group. Based on the results of morphological, physiological, biochemical and molecular analysis, the isolate was designated as Streptomyces sp. HU-S-01. At present, more than 30 isolates of pyrethroid-degrading strain have been reported and yielded a variety of micro-organisms. Most of them are bacteria including Pseudomonas sp., Serratia plymuthica, Acidomonas sp., Micrococcus sp., Bacillus sp. (Grant et al., 2002; Mandar et al.,2005; Tallur et al., 2008) while few are fungi including Aspergillus niger whereas only one belongs to actinomycetes (Klebsiella sp.) (Liang et al., 2005). To our knowledge, HU-S-01 is the first reported strain in the genus Actinomycetes capable of degrading cypermethrin. Optimum conditions for the biodegradation of cypermethrin by HU-S-01 Fig. 2 depicts the growth of HU-S-01 and degradation of cypermethrin at different temperatures. The results reveal that HU-S-01 is a strain that can't tolerate high temperature. The strain was able to grow well in the temperature range of 20-30 oC with the optimum temperature 26-28 oC but could not tolerate temperature higher than 34 oC. The trend of cypermethrin degradation was consistent with the growth of HU-S-01. In the temperature range of 26-28 oC, HU-S-01 degraded about 90 % cypermethrin within 24 h but degradation was very low (10 %) at the temperature higher than 34 oC. The effects of initial pH on growth of HU-S-01 and degradation of cypermethrin are shown in Fig. 3. The results indicate that the adaptive pH value for HU-S-01 growth was between 6 and 9. The higher cypermethrin degradation was observed between pH 7.0 and 8.0, with the optimum value of pH 7.5. Non significant degradation was observed at extreme pH values of 5.5 and 10.0. As the strain HU-S-01 is aerobic, so the dry weight of the HU-S-01 and the degradation rate of cypermethrin are inversely proportional to the liquid volume. The results shown in Fig. 4, express that at the liquid volume of 50 mL, HU-S-01 had the best growth and maximal degradation rate for cypermethrin whereas Fig.5 shows the removal of different concentrations of cypermethrin by HU-S-01. The strain degraded all the concentrations of cypermethrin lower than 100 mg/L. At the initial cypermethrin concentrations of 20 and 50 mg/L, the strain completely degraded cypermethrin within 24 and 30 h as shown in Fig. 6, but at 150 mg/L to 250 mg/L, degradation was incomplete even after 48 h. Normally, the reported contaminations of cypermethrin in soil and water were not as high as 250 mg/L, that has been used in this experiment (Zhao et al., 2005; Shen et al., 2006; Chen et al., 2007). So the ability to degrade high cypermethrin concentrations and the conditions for HU-S-01 growth suggested that this strain could be an effective bioaugmentation agent. Degradation of 3-phenoxybenzoic acid 3-PBA is the major metabolite after hydrolysis of cypermethrin in soil and water (Maloeny et al., 1992). It had anti-estrogenic activity in genetically modified yeast cells (Tyler et al., 2000). The degradation of cypermethrin and 3-PBA was carried out by HU-S-01 in the present studies. As shows in Fig. 6, HU-S-01 completely degraded 3-PBA and cypermethrin at the concentration 50 mg/L but the degradation rate of 3-PBA was slower than cypermethrin. It completely degraded the 3-PBA at 50 mg/L in 96 h while to completely degrade the same concentration of cypermethrin just required 30 h. At present a lot of pyrethroid-degrading strains but a few 3-PBA degrading strain have been reported. Edward and Akhtar (1991) find a Pseudomonas strain that could grow on 3-PBA and converts it to phenol. But the degradation of cypermethrin and 3-PBA by the same strain is a rare finding. In this study, HU-S-01 was found to degrade not only cypermethrin but its hydrolysis metabolite 3-PBA as well that could be safer to the environment. Enzyme activities and putative degradation pathway analysis Enzyme activity in cell free extract of HU-S-01 was studied using cypermethrin and 3-PBA as substrate. The kinetic constants of cell extract for cypermethrin and 3-PBA such as maximum reaction rate (Vmax), michaelis constant (Km), catalytic constant (Kcat) and specificity constant (Kcat/ Km) were estimated respectively by Lineweaver-Burk plot. As shown in Table 3, the cell free extract of HU-S-01 degraded cypermethrin and 3-PBA, but the affinity and maximum reaction rate with substrates were different. It infers that the cell free extract has certain specificity to the substrates. The Km, Vmax and Kcat for cypermethrin were higher than 3-PBA, revealing that the degradation efficiency of HU-S-01 enzyme was higher for cypermethrin as compared to 3-PBA. The value of Kcat/ Km are the specificity constant of enzyme, indicating the specificity of an enzyme to a substrate. The high value of Kcat/ Km shows the specificity of enzymes in HU-S-01to cypermethrin and 3-PBA. Degradation products and pathway for cypermethrin The degradation products of cypermethrin by strain HU-S-01 were extracted and identified by GC-MS. The metabolite peaks were identified using documented date from National Institute of Standards and Technology (NIST) library database. The GC chromatograms are shown in Fig. 7 There are four cypermethrin peak (B1, B2, B3, B4) and one peak (A) of metabolism were observed at 24h, the peak A (RT=14.793 min) was identified as 3-Phenoxybenzaldehyde, a hydrolyzate of cypermethrin by strain HU-S-01. However, the peak of cypermethrin and 3-Phenoxybenzaldehyde disappear after 96 h, it suggests that cypermethrin and 3-Phenoxy- benzaldehyde were degraded completely by HU-S-01. Results of the study depict the degradation of cypermethrin by enzymes in Hu-S-01 and probably two kinds of enzymes took part in it. In the molecular structure of pyrethroid insecticides there is an ester bond which is not as firm as other chemical bonds. The first step in the microbial degradation and detoxification of pyrethroid compounds is the hydrolysis of carboxyl ester linkage (Liang et al., 2007). When the cypermethrin was metabolized by micrococcus sp., it was first hydrolyzed into two parts followed by the further oxidation and dehydrogenation of 3-PBA (Tallur et al., 2008). The metabolites of allethrin degradation by Acidomonas sp. were cyclopropanecarboxylic acid, 2,2-dimethyl-3-(2-methyl-1-propenyl), 2-ethyl-1,3-dimethyl-cyclopent-2-ene-carboxylic acid, chrys-anthemic acid and allethrolone [2-cyclopenten-l-one, 4-hydroxy-3-methyl-2(-2-propenyl)], so the proposed pathway for allethrin degradation by Acidomonas sp. is through a hydrolytic pathway followed by oxidation and dehydrogenation (Mandar et al., 2005). Like other pyrethroid insecticides, the ester bond in the cypermethrin is the most fragile site so the hydrolytic enzyme might cause the cypermethrin to hydrolyze into 3-PBA and dichloroving acid, as shown in fig.8 This might be the main way of primary metabolism by HU-S-01. As the C and T sites of cypermethrin and some sites of 3-PBA are also very fragile and can be oxidized by oxidase. So, Oxidation on these sites might be the other possible pathway of cypermethrin degradation by HU-S-01. CONCLUSION In the present study, the cypermethrin-degrading strain Streptomyces sp. HU-S-01 was isolated from wastewater sludge collected from severage treatment pool of Zhongshan Aestar Fine Chemical Inc. Ltd. It revealed that temperature, initial pH, initial cypermethrin concentration and ventilation volume affected the growth of HU-S-01 and its degradation ability of cypermethrin. AS the HU-S-01 is aerobic strain, so under aerobic conditions, the optimum growth conditions were found to be 18-30 oC and 6.0-9.0 pH. It was unable to tolerate the temperature higher than 34 oC. The optimum growth temperature and initial pH for the strain was 26-28 oC and 7.5, respectively. At the optimum growth conditions, HU-S-01 efficiently degraded 10-250 mg/L cypermethrin within 48 h and completely degraded cypermethrin within 30 h at the concentration lower than 50 mg/L. This is an important feature of an organism to be utilized for biodegradation of cypermethrin residue in environment because the reported contamination of cypermethrin in soil or water is not as high as 250 mg/L. Degrading ability of 3-PBA is another important aspect of this strain. Degradation of cypermethrin and 3-PBA by the same strain is very important as the 3-PBA is the major metabolite of cypermethrin hydrolysis in soil and water. It could accumulate in the culture medium and effect the survival of wildlife, endanger human health, hence degrading the parent compound through catabolite repression if not degraded properly. Keeping in view the advantages, one could also visualize the development of cell-free enzymes for similar applications in the detoxification of cypermethrin residues on the surface of fruits and vegetables as well.
ACKNOWLEDGEMENTS The authors gratefully acknowledge the grants from the National Natural Science Foundation of China (No. 30871660), Project of Scientific Technological Planning of Guangdong Province (No. 2009B020310005). REFERENCES
© IRSEN, CEERS, IAU The following images related to this document are available:Photo images[st11004t2.jpg] [st11004f2.jpg] [st11004t3.jpg] [st11004f7.jpg] [st11004f5.jpg] [st11004t1.jpg] [st11004f1.jpg] [st11004f3.jpg] [st11004f6.jpg] [st11004f4.jpg] [st11004f8.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}