 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
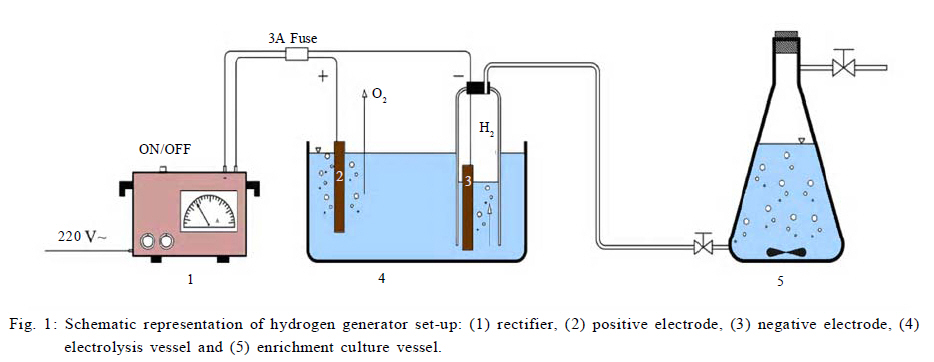
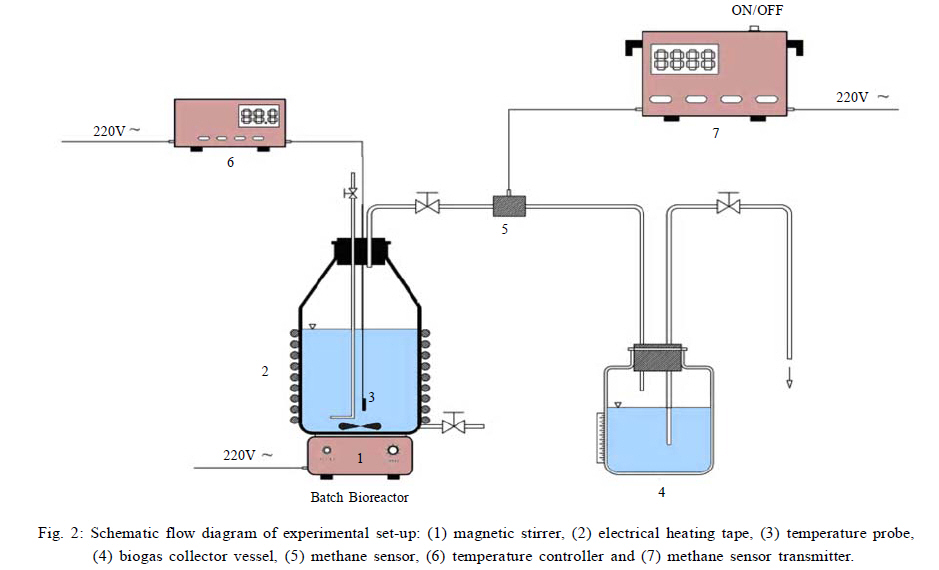
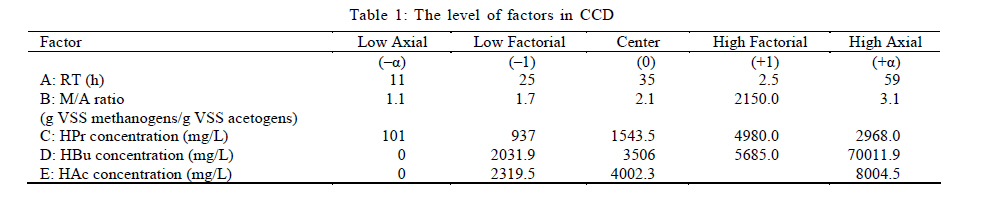
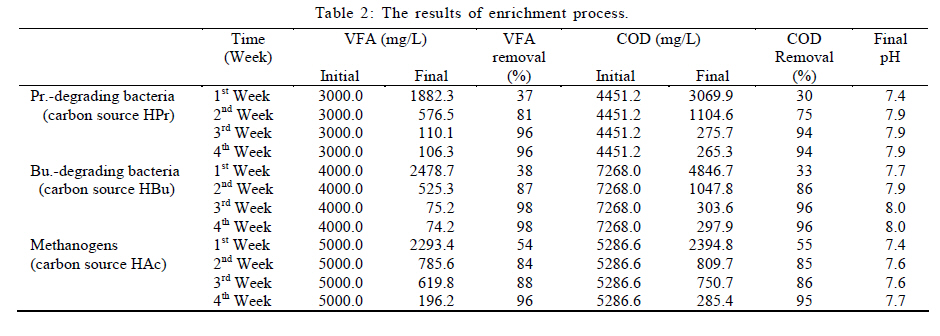
International Journal of Environment Science and Technology, Vol. 8, No. 1, 2010, pp. 83-96 Study of syntrophic anaerobic digestion of volatile fatty acids using enriched cultures at mesophilic conditions 1T. Amani; 1*M. Nosrati; 1S. M. Mousavi; 2R. K. Kermanshahi 1Biotechnology Group, Department of Chemical Engineering, Faculty
of Engineering, Tarbiat Modarres
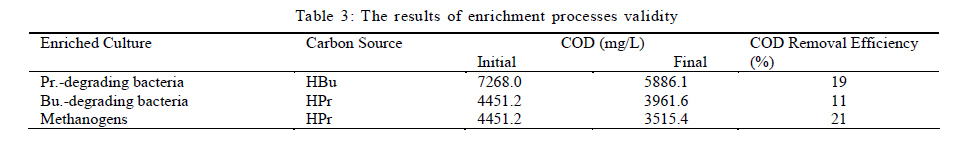
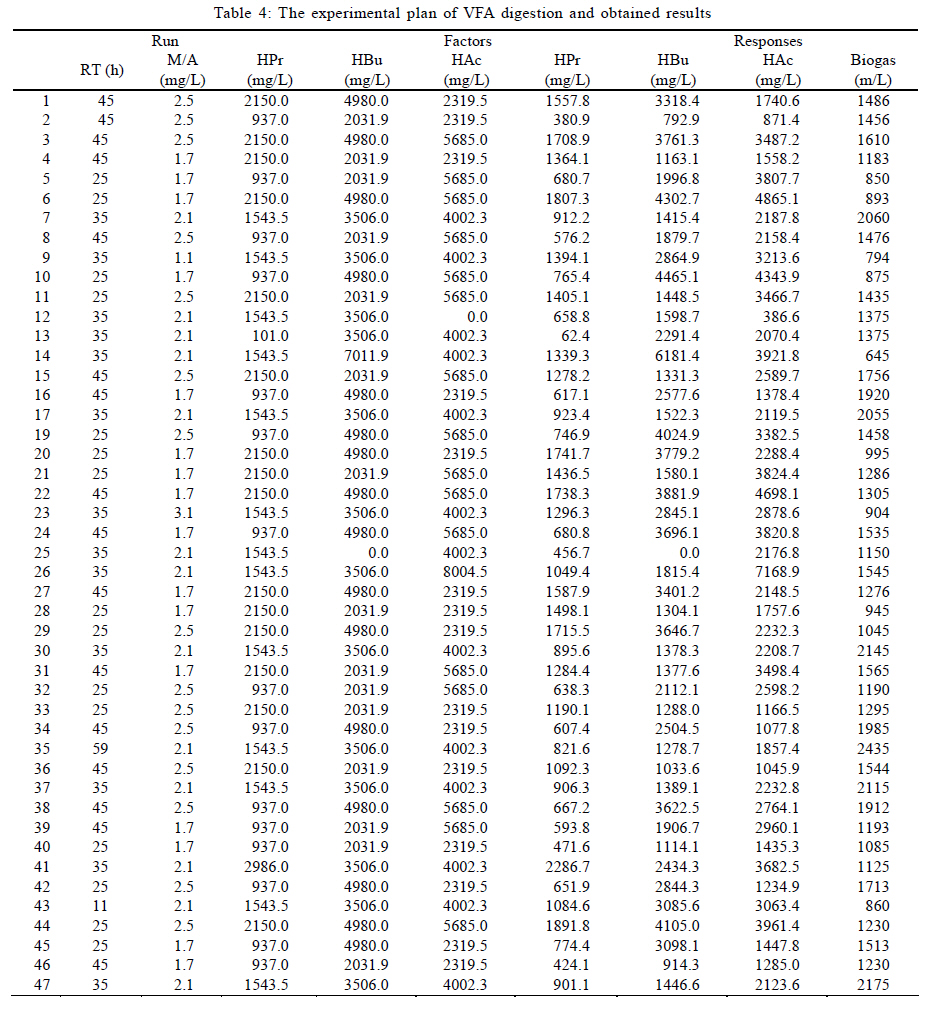
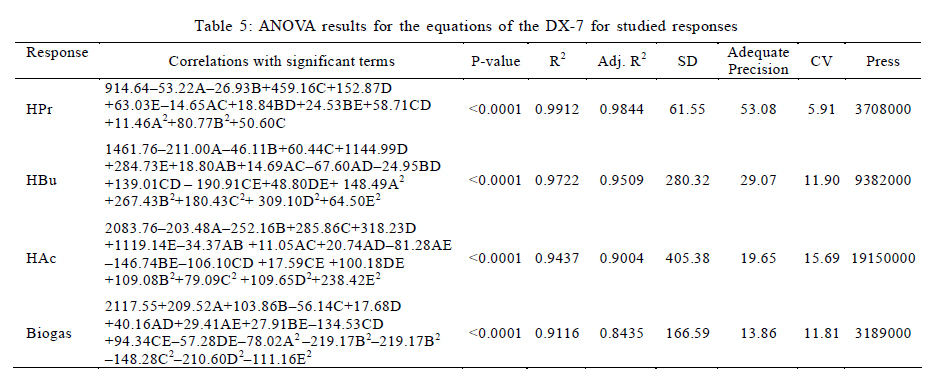
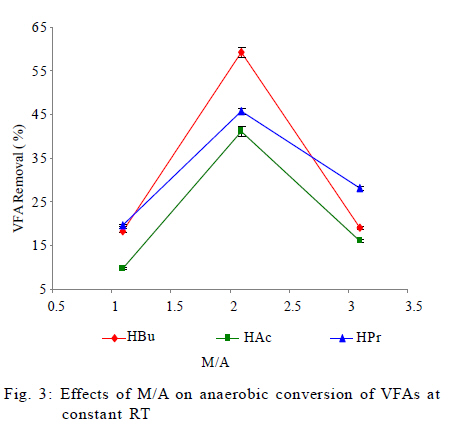
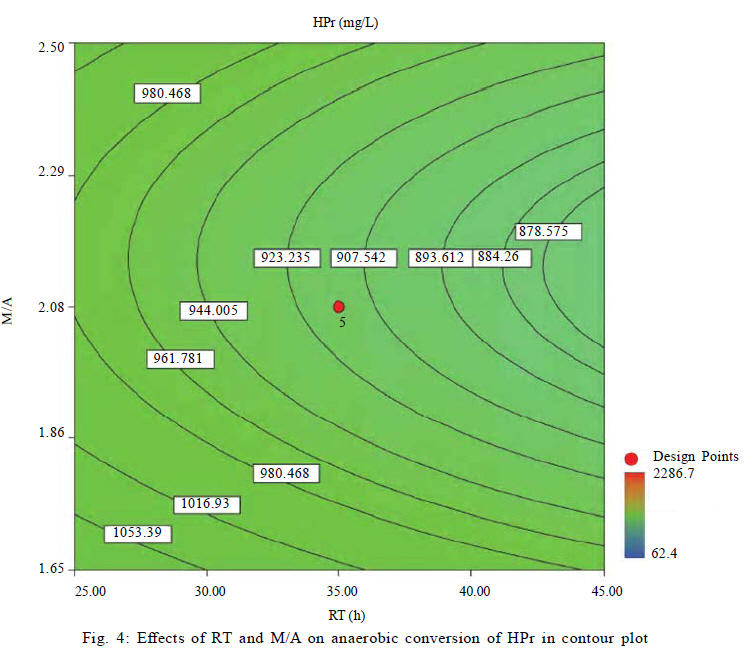
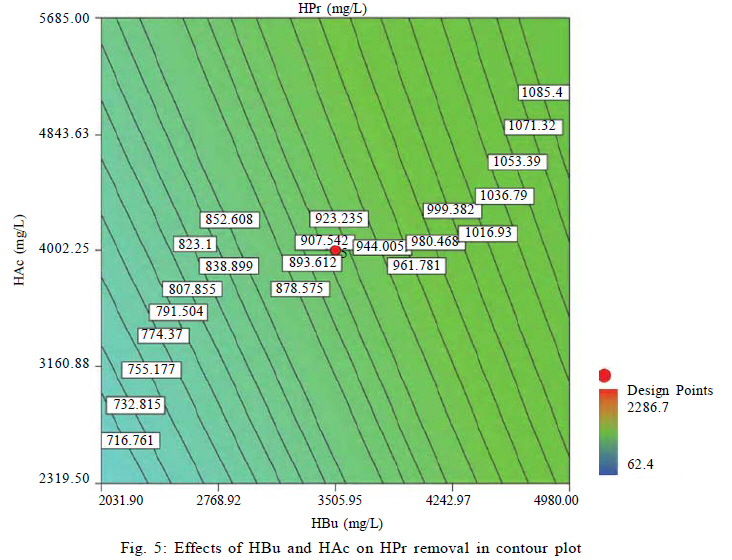
University, Tehran, Iran *Corresponding Author Email: mnosrati20@modares.ac.ir Tel.: +9821 82884372; Fax: +98 21 82883381 Received 30 June 2010; revised 11 September 2010; accepted 1 November 2010; available online 1 December 2010 Code Number: st11008 ABSTRACT: Volatile fatty acids are the most important intermediates in anaerobic digestion, and their degradations are extremely complicated thermodynamically. In this research, syntrophic anaerobic digestion of volatile fatty acids using enriched acetogenic and methanogenic cultures in a batch reactor at mesophilic conditions was investigated. Interactive effects of key microbiological and operating variables (propionic, butyric and acetic acids, retention time and methanogen to acetogen populations ratio) on the anaerobic degradation of volatile fatty acids were analyzed. Acetogenic and methanogenic anaerobes in the granular sludge from an up-flow anaerobic sludge blanket reactor were enriched at mesophilic conditions within a period of four weeks, separately. Enriched cultures were mixed with known proportions and then used in the bioreactor. Experiments were carried out based on central composite design and analyzed using response surface methodology. Four parameters (final concentrations of propionic, butyric and acetic acids and biogas production) were directly measured as response. Also, the optimum conditions for volatile fatty acid degradation were found to be 937.5 mg/L, 3275.5 mg/L, 2319.5 mg/L, 45 h and 2.2 proportions for propionic acid, butyric acid, acetic acid, retention time and methanogen to acetogen populations ratio, respectively (corresponding to maximum volatile fatty acid removal efficiencies and biogas production). The results of the verification experiment and the predicted values from the fitted correlations at the optimum conditions were in close agreement at a 95% confidence interval. The present study provides valuable information about the interrelations of quality and process parameters at different values of microbiological and operating variables. Keywords: Acetogens; Acetogenesis reactions; Methanogens; Methanogenesis reactions INTRODUCTION The anaerobic digestion process is a sequential, complex biochemical process, in which organic compounds are mineralized to biogas (mainly consisting of CH4 and CO2) through a series of reactions mediated by several groups of microorganisms. This process, from the view point of microbiology, follows four major steps: hydrolysis, acidogenesis, acetogenesis and methanogenesis (Gerardi, 2003; Metcalf and Eddy, 2004; Bitton, 2005; Nwuche and Ugoji, 2008; Nwuche and Ugoji, 2010). Of the several intermediate stages, acetogenesis and methanogenesis are the most critical steps, in which acetogens (propionate and butyrate degrading bacteria) and methanogens (hydrogenotrophs and aceticlasts) form special constructions and are interrelated in what is called a "syntrophic interaction" (Schink and Stams, 2005). Propionate and butyrate are the most important intermediates in the syntrophic reactions, and because of thermodynamic restrictions, their degradations are regarded as the rate limiting steps in the anaerobic digestion. They often accumulate in the anaerobic digester when the process becomes unstable (Kida et al., 1993; de Bok et al., 2004; Wong et al., 2009; Shah et al., 2009; Liu et al., 2010; Hooshyari et al., 2009). The anaerobic oxidations of propionate and butyrate to acetate, CO2 and H2 are highly endergonic (DG°Propionate= +76.1 and DG°Butyrate= +48.1 kJ/mol at 25 °C) and do not occur naturally and thermodynamically, but these reactions can be accomplished by the syntrophic cooperation of propionate and butyrate- oxidizing bacteria and H2/formate-scavenging partners, which maintain a low H2 partial pressure (Schink and Stams, 2005). The anaerobic conversion of butyrate is not as unfavorable thermodynamically as the anaerobic conversion of propionate, and its accumulation rarely has been reported in the literature. Studies on the volatile fatty acid (VFA) conversions have been mainly focused on the syntrophic association in the co-cultivation of acetogens with methanogens (Plugge et al., 2002; de Bok et al, 2005; Kosaka et al., 2006; Babel et al., 2007; Tatara et al., 2008). In these studies, the effects of all VFAs, especially propionate, on the activity of acetogens and methanogens have been considered. Several investigators have described the toxic effects of VFAs in the anaerobic digestion process (Pullammanappallil et al., 2001; Han et al., 2005; Gallert and Winter, 2008; Uneo and Tatara, 2008), but the extent of this inhibition has not been properly studied. The main aim of this research was to investigate, analyze and model the syntrophic anaerobic degradation of VFAs and the performance of the enriched acetogenic and methanogenic microorganisms. It is often necessary to introduce enriched seeding microorganisms to start up anaerobic digesters. The enriched cultures improve the syntrophic reactions. As another contribution in this study, a novel method was applied to enrich the acetogens and methanogens utilizing specific substrates. In the last few years, response surface methodology (RSM) has been applied to analyze, optimize and evaluate interactive effects of independent factors in numerous chemical, biochemical and bioenvironmental processes (Wang et al., 2005; Altaf et al., 2006; Aktaþ et al., 2006; Zinatizadeh et al., 2006; Ghorbani et al., 2008). Analysis and modeling of anaerobic digestion processes, particularly syntrophic anaerobic degradation of VFAs, using RSM has not yet been reported. In this research, the RSM was used to analyze and model the process with respect to the simultaneous effects of five microbiological and operating variables (propionic acid (HPr), butyric acid (HBu), acetic acid (HAc), methanogen to acetogen populations ratio (M/A) and retention time (RT)) and four parameters (HPr, HBu, HAc and biogas production) were assessed as responses. The significant factors and a continuous response surface of the main parameters were developed to provide an optimal region that satisfies the process specifications. This research was carried out in Bio-pilot Laboratory, Biotechnology Group, Chemical Engineering Department, Tarbiat Modares University during November 2009 to February 2010. MATERIALS AND METHODS Enrichment To enrich the acetogens and methanogens, the granular sludge from an up-flow anaerobic sludge blanket (UASB) reactor that is employed in the treatment of dairy wastewater was utilized. The pH of the granular sludge was 7.4, and the concentrations of volatile suspended solid (VSS) and total suspended solid (TSS) were 67.2 g/L and 92.4 g/L, respectively. To enrich the anaerobes, the minimal medium contained the following: KH2PO4 (0.6 g/L), Na2HPO4 (1.33 g/L), NH4Cl (0.3 g/L), CaCl2.H2O (0.11 g/L), MgCl2.6H2O (0.1 g/L), NaHCO3 (4.0 g/L), Na2S.9H2O (0.025 g/L), FeCl2 (0.45 g/L), H3BO4 (0.04 g/L), ZnCl2 (0.032 g/L), CuCl2 (0.0063 g/L), MnCl2 (0.0295 g/L), CoCl2 (0.0305 g/L), NiCl2 (0.0062 g/L), cysteine hydrochloride (0.05 g/L) and the yeast extract (0.05 g/L). The medium cultures were prepared with distilled water. Before inoculation, O2 was removed from all the culture vessels and media by sparging with the N2 gas for 10 min. The enrichment of propionate- and butyrate-degrading bacteria and methanogens was performed at 37±1 °C. They were enriched in three Erlenmeyer flasks with working volumes of 1 liter, independently. The carbon sources for enrichment of propionate-, butyrate-degrading bacteria and aceticlasts were HPr (3000 mg/L), HBu (4000 mg/L) and HAc (5000 mg/L), respectively. To enrich the anaerobic hydrogenotrophs, hydrogen was produced by the electrolysis of tap water. The schematic representation of the hydrogen generator is shown in Fig. 1. This system consists of a rectifier (maximum voltage: 35 V, and maximum current: 15 A), 2 stainless-steel electrodes (dimensions: 2×300×30 mm3) and a vessel containing 8 liters of tap water. Conductivity of water was increased by the addition of 3 to 4 milliliters of concentrated sulfuric acid (98 %). Under these conditions the maximum electrical current passing through the water was 3 A. H2 gas was produced and collected at the negative electrode, followed by venting it to the vessel containing the granular sludge and HAc as the carbon source. Enrichment of hydrogenotrophs with the H2 gas was performed at a flow rate of 95 mL/h. The initial pH was adjusted at 7±0.1 with NaOH. The temperature conditions were controlled by using a water bath. The magnetic stirrer was used to mix contents of the vessels during the enrichment process. Mixing was performed daily with the intensity of 50 rpm and duration of 5 min. The initial enrichment cultures were started by adding 200 mL of the granular sludge to 800 mL of the minimal medium with the carbon source (20 % (v/v) inoculation). Every week, samples of each vessel were drawn, and their pH and VFA concentrations were assayed. The inoculated culture (200 mL) was then transferred to the fresh medium (800 mL). The removal efficiencies of VFAs (more than 95 % removal efficiency) were considered as the criteria to perform the enrichment of the anaerobic microorganisms. After a few transfers, all done according to the defined criteria, the enrichment processes were considered complete and the anaerobic microorganisms continued to remain in the cultures. Substrates The synthetic wastewater, containing pure HPr, HBu and HAc (Merck Inc., Germany), as well as tap water with different concentrations (low to high loads), were used as the major carbon sources and electron donors in this study. To supply adequate nitrogen and phosphorus for the microorganisms, chemical oxygen demand (COD):N:P ratio was maintained at 100:4:1. O2 was removed by N2 sparging for 10 min before feeding to the bioreactor. The pH of the feed and within the reactor was not regulated throughout the experiment (4.55.5). To increase the alkalinity, NaHCO3 (4 g/L) was added to the feed. Batch reactor and operating conditions A flow diagram of the experimental set-up is shown in Fig. 2. The batch reactor was a glass cylinder with a diameter of 10 cm and a height of 15 cm (working volume 1 L). Effluent samples were drawn from the bottom of the reactor using a sampling port. Prior to the experiments, a 200 mL (20 % (v/v) inoculation) mixture of the enriched cultures (methanogens and acetogens) with defined M/A was used to seed the batch reactor. For starting up the bioreactor, the ratio of methanogens to acetogens (M/A) was taken as the relative amount of their VSS concentrations. An electrical heating tape (heating capacity: 40 W/m) was attached to the outside surface of the reactor and a temperature probe was connected to the transmitter. The temperature of the reactor was set to the mesophilic (37±1 °C) condition. To ensure efficient transfer of the intermediates and to release gas bubbles trapped in the medium mixing was performed with an intensity of 50 rpm, duration of 5 min per each 10 h, using a magnetic stirrer. The produced biogas was vented out the top of the bioreactor through a connecting pipe and was collected by the water-displacement method. Generally, concentrations of HPr, HBu and HAc, RT, temperature, mixing, pH and the relative population of syntrophic microorganisms, all affect the anaerobic digestion of the VFAs. To investigate the syntrophic anaerobic digestion of VFAs, concentrations of HPr, HBu and HAc, RT and M/A were selected as the primary factors affecting the study, and central composite design (CCD) was used to design the experiments. The RSM used in the present study was CCD, which is the most popular response surface method (Anderson and Whitcomb, 2000; Myers and Montgomery, 2002; Söteman et al., 2005). The levels of the factors are shown in Table 1. Each factor was varied at five levels, whereas the other parameters were kept constant. Consequently, 47 experiments were conducted; 32 of them organized in a full factorial design, 10 experiments were related to axial points and 5 experiments were done in center points. The remaining five involved repetition of the central idea to get a good estimate of the experimental error. Experiments were designed by Design Expert (DX) Software (State-Ease Inc., version 7.0.0), which is a Windows-compatible software. It provides efficient design of the experiments (DOEs) for identification of the vital factors that affect the process and uses RSM to determine the optimal conditions. Analytical methods Methane concentration in produced biogas was determined with Figaro TGS 2611 methane sensor (made in USA). The sensing element is comprised of a metal oxide semi-conductor layer formed on an alumina substrate of a sensing chip together with an integrated heater. In the presence of a detectable methane, the sensor's conductivity increases depending on the methane concentration. A simple electrical circuit can convert the change in conductivity to an output signal which corresponds to the methane concentration. The liquid samples were first centrifuged at 10,000 rpm for 15 min, followed by the HCl acidification, and finally assayed. The VFAs (HAc, HPr and HBu) were quantified using the Agilent gas chromatograph (model 7890), equipped with an auto-injector (7683 B series), a flame ionization detector (FID) (H2 flow rate: 35 mL/min, air flow rate: 350 mL/min) and a Chrompack Cp-Wax 52 CB fused-silica column (25 m × 0.32 mm i.d. and 0.2 ìm film thickness). The injector and detector temperatures were kept constant at 240 °C and 280°C, respectively. Helium (He) was used as the carrier gas at a flow rate of 3 mL/min and makeup flow rate of 5 mL/min. The oven temperature was programmed at 40 °C for 4 min, raised to 180 °C at 30 °C/min, and then held at 180 °C for 1 min. The VSS, TSS and COD were determined according to the standard methods (APHA, 1992). The pH was measured using the Metrohm 620 pH meter (made in Germany). RESULTS AND DISCCUTION Enrichments Concentrations of COD, VFAs (HPr, HBu and HAc) and pH, during the enrichment processes were measured at the end of each transfer. The initial and ultimate concentrations of COD, VFAs and pH in the enrichment cultures of propionate- and butyrate-degrading bacteria (acetogens) and methanogens are shown in Table 2. The results of COD and VFA removal efficiencies were in good agreement with each other. Increases of HPr, HBu and HAc removal efficiencies or decreases of their ultimate concentrations were stepwise gradual. These results showed that after four transfers (four weeks) VFA removals were greater than 95 %. Thus, according to the defined criteria, it could be concluded that acetogens and methanogens in the granular sludge prevailed in the cultures. During the enrichment process the initial granules were broken down gradually, and in the end of the fourth week, almost all of the microorganisms were completely dispersed. The reason is that only one VFA was used for the enrichment of each group of microorganisms and the syntrophic reactions were not complete. Therefore, the enriched cultures obtained by the reaction, especially acetogens, were not granular and were fine or dispersed at the end of the enrichment processes. Enriched cultures (with known ratio) were thickened and then used for the syntrophic anaerobic digestion of VFAs in the bioreactor. The concentrations of the propionate-degrading bacteria in the enriched cultures were VSS=68.4 g/L and TSS=78.2 g/L. For butyrate-degrading bacteria, the concentrations were VSS=71.2 g/L and TSS=81.3 g/L; and for methanogens, they were VSS=74.3 g/L and TSS=84.5 g/L. To verify the performance of each enriched culture against a different feed, the enriched cultures were inoculated with other carbon sources, i.e., HPr was added to the enriched sludge of butyrate-degrading bacteria and to the enriched sludge of methanogens. Moreover, HBu was added to the enriched sludge of propionate-degrading bacteria. The minimal medium, temperature and pH were the same as during the enrichment processes, but the incubation time was 10 days. The concentrations of the initial and ultimate COD are shown in Table 3. The results showed that just before 10 days in culture, low COD removal efficiencies of 19, 11 and 21 percent were observed for the propionate-, butyrate-degrading bacteria and methanogens, respectively. These results again revealed that the enriched cultures predominantly contained the propionate- and butyrate-degrading bacteria, as well as the methanogens. Statistical analysis Forty seven experiments were designed using CCD. The experimental conditions and their responses for the mesophilic anaerobic digestion process are shown in Table 4. Because four responses were investigated in this study, the results were inserted into DX-7 software and were fitted to quadratic correlations and then, correlations were found to be adequate for the prediction of the response variables. The analysis of variance (ANOVA) results for the responses have been summarized in Table 5. The relatively high values of R2 indicate that the quadratic equations for output HPr, HBu, HAc and biogas production are well capable of representing the system under the given experimental domain. As can be observed in Table 5, there is very little difference between R2 and adjusted-R2, which shows there is a good chance that important terms are included and considered in the correlations. The correlation terms in the equations are those essential terms that remained after elimination of insignificant variables and their interactions. As seen in the table, the fitted correlations are significant with a 95 % confidence interval (p-value < 0.0001) for the four responses. The correlation adequacy was tested through lack-of-fit F-tests (Montgomery, 1991). The lack-of-fit F-statistic was not statistically significant because the p-values were greater than 0.05. Adequate precision is a measure of the range of predicted response relative to its associated error or, in other words, a signal-to-noise ratio. Its desired value is 4 or more (Mason et al., 2003). The values were found to be acceptable for the three correlations; see the adequate precision column in Table 5 with values that are much more than 4. Simultaneously, low values of the coefficient of variation (CV) for the responses indicated good accuracy and dependability of the experiments. Effects of M/A and RT on VFA removal Anaerobic digestion of HPr and HBu are highly endergonic and do not occur naturally (from the view point of thermodynamic principles) in anaerobic digesters (Schink and Stams, 2005). Small populations of methanogens are not able to metabolize the hydrogen and HAc produced by the acetogens. Increasing the methanogenic population could be used as a method to promote these reactions efficiently (Schmidt and Ahring, 1995). In this study, to investigate the effect of the methanogenic population on the anaerobic conversion of VFAs, the ratio was adjusted within a range of 1 to 3. Fig. 3 shows the effect of M/A on VFA removals. When M/A was increased from 1.1 to 2.1 (when HPr = 1543.5 mg/L, HBu = 2000.8 mg/L, HAc = 2499.9 mg/L and RT = 35 h), the removal efficiencies of HPr, HBu and HAc increased from 10, 18 and 20 to 41, 59 and 46 %, respectively. However, when M/A = 3.1, the removal rates decreased. This showed that in very high M/A ranges, the acetogens (propionate- and butyrate-oxidizing bacteria) were not sufficient to degrade high concentrations of HPr and HBu. In this condition, high levels of HPr and HBu inhibited the acetogenic reactions and suppressed the growth of acetogens; consequently, their degradations slowed down. As seen in Fig. 3, higher M/A also caused HAc removal to decrease; this could be attributed to inhibition of the growth of the methanogenic population in the presence of high levels of HAc and reduced pH. Accordingly, in the high M/A ranges the activities of all anaerobes dropped off significantly. The RT had a positive effect on VFA removal: with increasing RT, anaerobic microorganisms adapted to low pH and removal efficiencies improved. Fig. 4 illustrates the effects of RT and M/A on anaerobic conversion of HPr. From the fitted correlations in Table 5, it can be concluded that the coefficient of RT is more significant than the M/A coefficient for the final concentrations of HPr and HBu. The interaction of RT and M/A is not very significant because the coefficient of the interaction term of the correlations is small. Effects of VFAs on HPr conversion Anaerobic oxidation of HPr is inhibited by VFAs; the extent of this inhibition is dependent on the VFA concentrations and the pH (Siegert and Banks, 2005). Methanogens are extremely sensitive to pH, with optima between 6.8 and 7.4 (Turovskiy and Mathai, 2006), but acetogenic microorganisms are somewhat less sensitive and can function in a wider range of pH. Generally, the presence of VFAs leads to a drop in pH in a digester, and their toxicities are steeper when the pH is below 7 (Hwang et al., 2004). Because acetogenic degradation of HPr and HBu are extremely endergonic, their anaerobic degradations are repressed thermodynamically. Also, degradation of HPr and HBu is inhibited via a pH drop in the digester. On the other hand, methanogenic conversion of HAc is exergonic; thus it could be expected that its inhibitory effect is due to the pH drop, which reduces the growth of methanogens. In fact, methanogenic oxidation of HAc stimulates HPr degradation thermodynamically. Fig. 5 shows the effects of HBu and HAc on anaerobic oxidation of HPr. As seen, when concentrations of HBu and HAc were increased, the HPr removal decreased. The maximum conversion of HPr was obtained when HBu and HAc were diminished. Data from the experiments in Table 4 confirmed these results. Furthermore, because the mechanisms of HBu and HAc inhibition are rather different, their observed inhibitory effects on HPr removal were dissimilar. The coefficient of the HBu term (153) in the fitted correlation in Table 5 was, as expected, greater than that of HAc term (63). Since HAc accumulated when there were high levels of M/A, the relative inhibitory effects of HBu and HAc were not strongly dependent on M/A; therefore, it seemed that the level of HAc inhibition with higher M/A was apparently only a little less than HBu. HPr had the largest inhibitory effect on the HPr removal; on the other hand, its coefficient in the fitted correlation (459) is much higher than that for HBu and HAc.
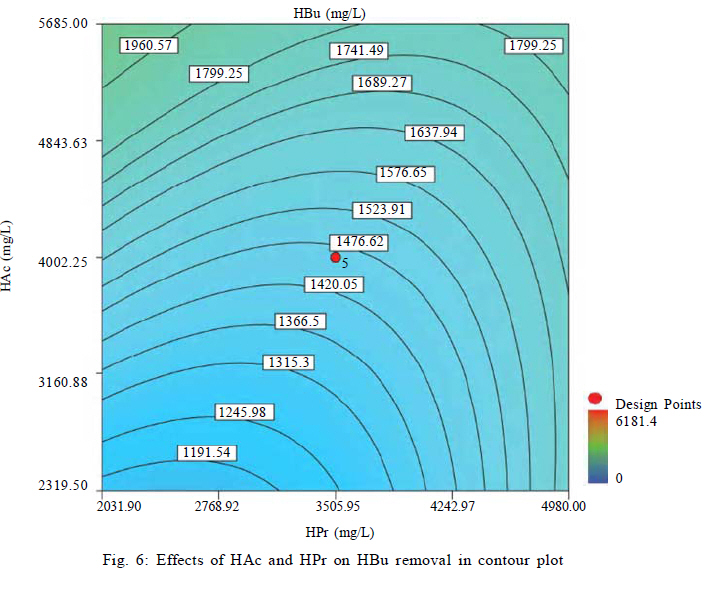
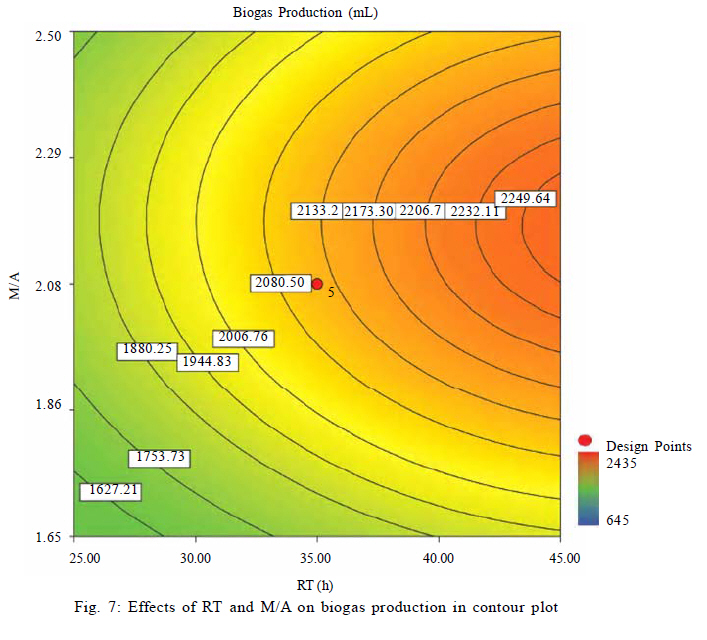
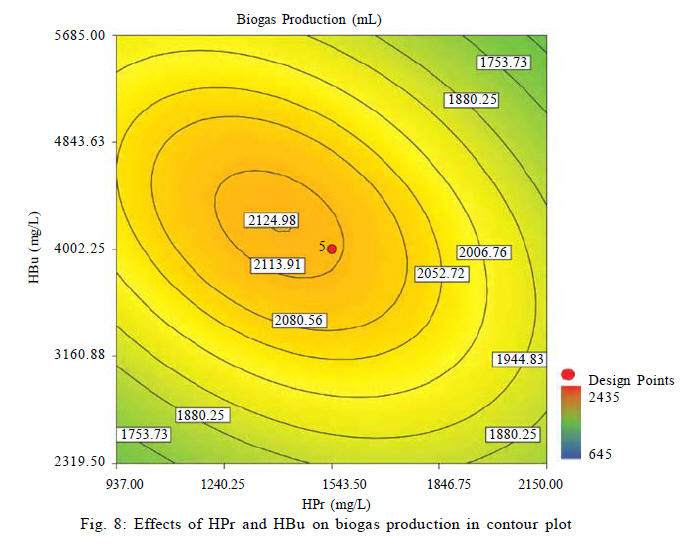
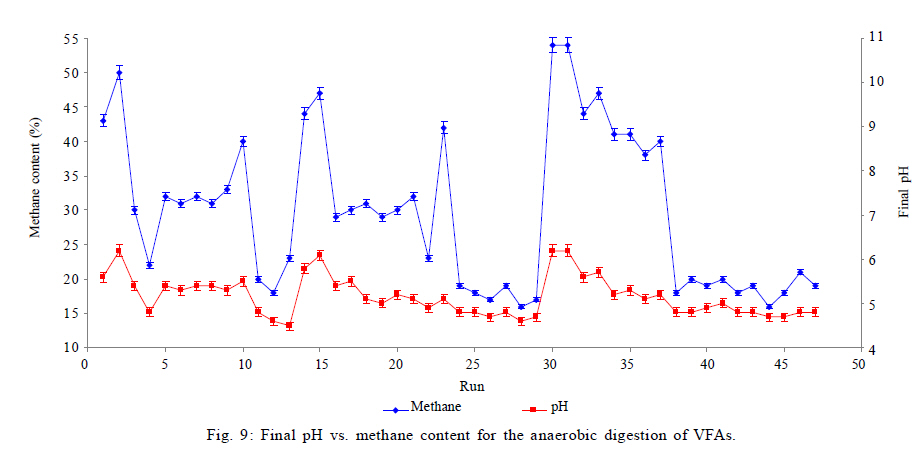
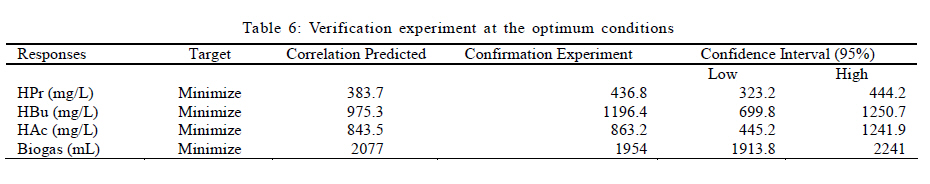
Effects of VFAs on HBu conversion Although anaerobic oxidation of HBu is not as thermodynamically unfavorable as conversion of HPr, the inhibition pattern of VFAs on anaerobic conversion of HBu is similar to HPr inhibition. Both HPr and HAc decrease the pH of the reactor; thus, they can inhibit the growth of acetogenic and methanogenic microorganisms. Thermodynamically HPr hinders, whereas HAc enhances, the anaerobic conversion of HBu. Accordingly, it could be expected that the inhibitory effect of HPr on HBu degradation is much higher than the inhibitory effect of HAc. The effects of HPr and HAc on HBu oxidation is shown in Fig. 6. The results revealed that, in spite of what it would be understood apparently from the above, the inhibitory effect of HAc was greater than that of HPr. The coefficients of their individual terms (60, 285 for HPr and HAc, respectively) in the fitted correlation confirmed this observation. The main reason could be the effect of HAc on pH reduction in the digester, which seriously hindered the growth of anaerobes. Another reason could be attributed to the spatial position or juxtaposition of the syntrophic acetogenic and methanogenic microorganisms (suspended growth). In the syntrophic cultures, diffusion of dissolved intermediates (H2/formate and HAc) from the acetogens (as producers) to the methanogens (as consumers) is the main mechanism; therefore, close microbial proximity plays an important role in this phenomenon. The relative spatial location and mobility of different syntrophic organisms in the bioreactor may play a significant role in controlling the inhibitory effect of HPr and HAc on HBu degradation. Consequently, the movement of butyrate-degrading bacteria probably was inadequate for trouble-free diffusion of intermediates. Similar to HPr, HBu itself had the largest inhibitory effect on HBu removal, because its coefficient in the fitted correlation (1145) is much higher than that for HPr or HAc. Biogas production from VFAs In this study, all variables directly or indirectly affected biogas production in the syntrophic anaerobic digestion of VFAs, and the extents of the influences were dependent on their levels and interrelated interactions. Fig. 7 shows the effects of RT and M/A on the biogas production. Increasing M/A enhanced the methanogenic populations; therefore, the rate of hydrogen consumption was increased, which led to an increase in the biogas production. With higher levels of M/A, the rate of hydrogen production from acetogenesis reactions (because of the reduction of acetogens) was decreased, and consequently biogas production was reduced critically. Thus, there was an optimum M/A for the biogas production, as seen in Fig. 7. The RT had a positive effect on the biogas production; RT and biogas production increased simultaneously. However, in very high RT ranges, the rate of biogas production was decreased and seemed to even stop. The fitted correlation in Table 5 for the biogas production shows that the effect of RT on biogas production was much greater than the effect of M/A, but RT/biogas production interaction was not an important term in the correlation. Fig. 8 illustrates the effects of HPr and HBu on the biogas production. HPr and HBu to some extent enhanced the biogas production, but in their higher levels, they strongly inhibited the growth of anaerobes, which led to reduction in biogas production. As a result, there were optimum amounts of HPr and HBu for the biogas production. The presence of VFAs in the anaerobic digester leads to a reduction in pH and an inhibition of growth in anaerobes, especially methanogens (Turovskiy and Mathai, 2006). The methanogenesis at low pH is attributed to the activity of hydrogen-utilizing methanogens that are likely more tolerant to the acidic conditions than the other methanogens (Kim et al., 2004). Because methanogenic activity is decreased in the low levels of pH, the methane yield is decreased significantly. Fig. 9 shows methane content versus final pH in the anaerobic digestion of VFAs. Although the pH in the digester was increasing during anaerobic digestion of VFAs, because of the high activity of enriched cultures, the methane content of the biogas was still low. Maximum VFA removal and biogas production According to the main objectives of this experiment (maximum VFA removals and biogas production), the optimum conditions were obtained to be HPr = 937.1 mg/L, HBu = 3275.5 mg/L, HAc = 2319.5 mg/L, RT = 45 h and M/A = 2.2. To check the accuracy of the fitted correlations of the optimum conditions at the 95 % confidence interval, the batch bioreactor was operated to compare actual responses with predicted responses. Table 6 presents the results of experiments conducted at optimum conditions. The accuracy of the optimum conditions from DOE experiments was checked that the experimental finding was in close agreement with correlation predictions. CONCLUSION Syntrophic anaerobic digestion of VFAs using enriched acetogenic and methanogenic cultures in a batch bioreactor at mesophilic conditions was investigated in this study. Several optimum conditions can be considered, but the main aim of this research was to maximize VFA removal that their anaerobic digestion at high concentrations thermodynamically is intricate or even impossible. The concentrations of HPr, HBu, HAc, M/A and RT were selected as the control factors, and HPr, HBu, HAc and biogas production were the main responses. Analysis of the experimental results using RSM revealed the interrelated interactions of the parameters. RT and M/A had positive effects on VFA removal; however, performance of the process at very high M/A drastically decreased. In the high M/A regions, the number of acetogenic bacteria was not sufficient to oxidize HPr and HBu effectively. The influence of RT on VFA removal and biogas production was more significant than M/A. HPr and HBu inhibited syntrophic oxidation of VFAs thermodynamically as well as via a pH drop. The effect of HBu on VFA removal was more significant, which might be due to spatial proximity or relative positions of suspended syntrophic organisms in the batch bioreactor. According to the fitted correlations, HPr was the most important parameter because it influenced all the responses and reduced the performance of the process. HAc stimulated VFA removal thermodynamically, but its inhibitory effect due to the pH drop was significant. Although syntrophic microorganisms, especially methanogens, are very sensitive to low pH, acetogenesis and methanogenesis occurred efficiently in this research work, and the enriched cultures did not lose their activity even at very low pH. Also, pH was increased during all of the experiments in the anaerobic digestion process. The maximum VFA removals and biogas production occurred when HPr=937.1 mg/L, HBu=3275.5 mg/L, HAc=2319.5 mg/L, RT = 45 h and M/A = 2.2. Syntrophic studies may be more accurate when ratios of M/A become more accurate. This may be achieved by obtaining optimum ratios of propionate-degrading bacteria to butyrate-degrading bacteria in the acetogenic microorganisms bulk. REFERENCES
© IRSEN, CEERS, IAU The following images related to this document are available:Photo images[st11008t1.jpg] [st11008f3.jpg] [st11008t2.jpg] [st11008f5.jpg] [st11008f9.jpg] [st11008t6.jpg] [st11008f8.jpg] [st11008t5.jpg] [st11008f4.jpg] [st11008f6.jpg] [st11008f2.jpg] [st11008t4.jpg] [st11008t3.jpg] [st11008f1.jpg] [st11008f7.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}