 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
International Journal of Environment Science and Technology, Vol. 8, No. 3, 2011, pp. 461-470 Isolation of hydrocarbonoclastic bacteria from bilge oil contaminated water 1 C. Sivaraman; 1 A. Ganguly; 2 M. Nikolausz; 1 *S. Mutnuri 1Department of Biological Sciences, Birla Institute of Technology and Science - Pilani, Goa Campus, Zuari
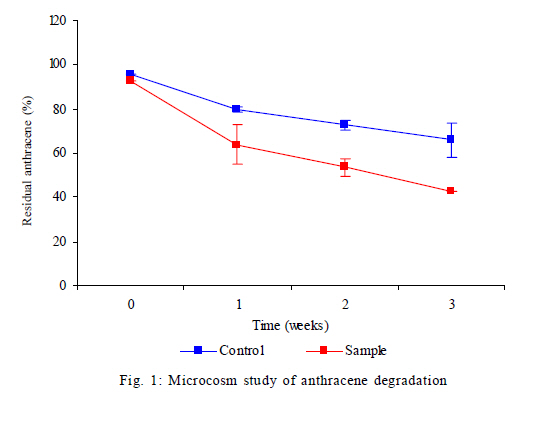
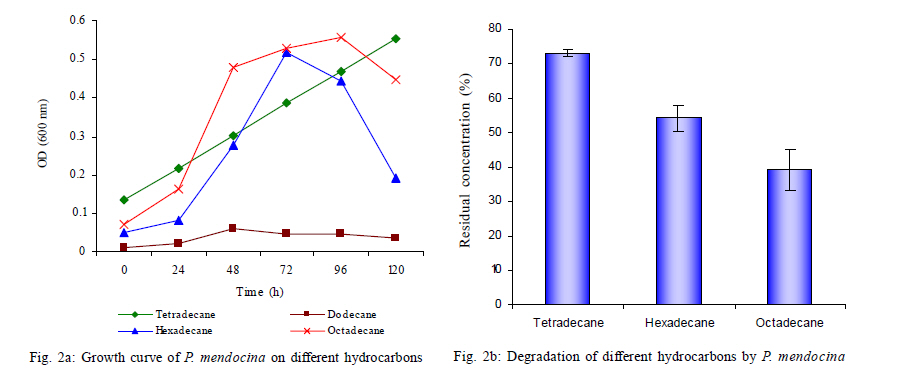
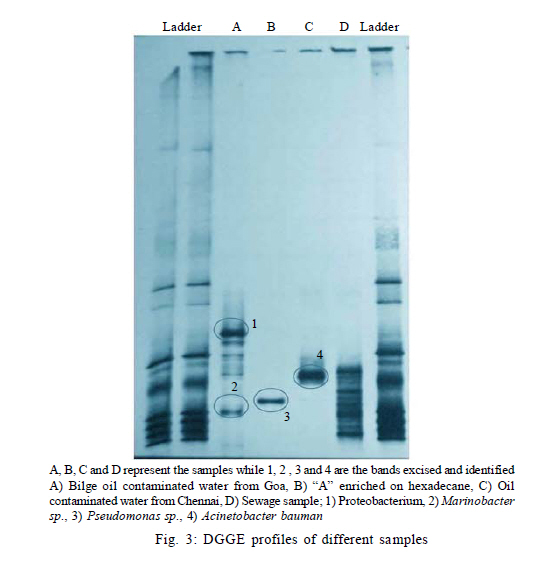
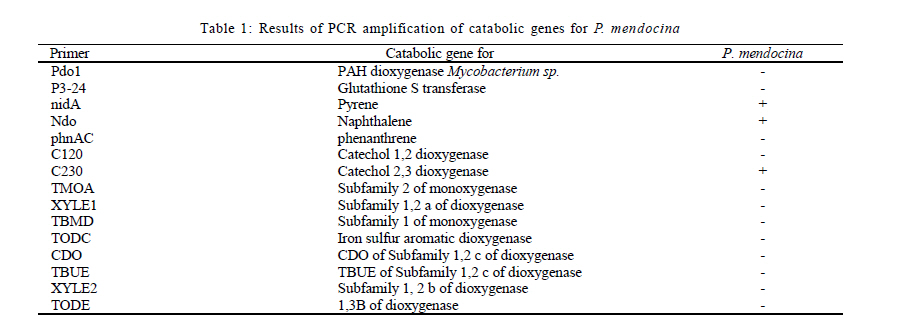
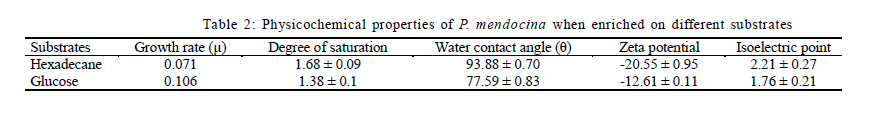
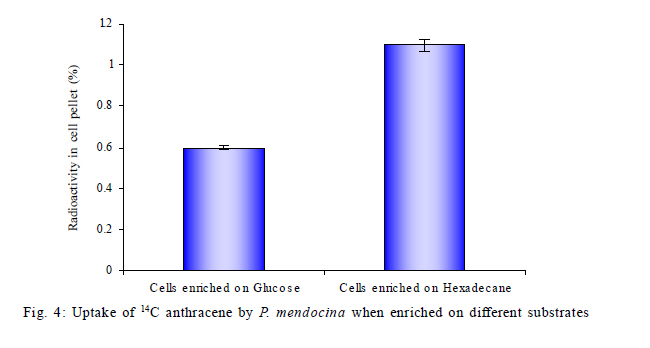
Nagar, India *Corresponding Author Email: srikanth.mutnuri@gmail.com Tel: +91832 2580 125, Fax: +91832 2557 033 Received 12 July 2010; revised 3 December 2010; accepted 18 January 2011 Code Number: st11042 ABSTRACT: Two bacterial strains, i.e. Pseudomonas mendocina and Ochrobactrum sp. were isolated from bilge oil contaminated water of Mormugao harbour, Goa, India and grown in a culture medium with hexadecane as the sole carbon source. Pseudomonas mendocina was used in further studies as it was the dominant strain. This strain effectively degraded tetradecane, hexadecane and octadecane leaving a residual concentration of about 73 %, 54 % and 40 % respectively in 120 h. Sequence analysis of the dominant bands from the denaturing gradient gel electrophoresis profiles revealed the differences between the genera of bilge oil contaminated sea water and its enrichment culture on hexadecane indicating a shift in community structure based on the type of substrate available. Pseudomonas mendocina amplified for the following catabolic genes namely C23O, nid and ndo. Based on the catabolic gene study the potential of the bacterial strain isolated, i.e. Pseudomonas mendocina seems to be interesting as it will be able to degrade polyaromatic hydrocarbons as well. Physicochemical properties of Pseudomonas mendocina indicates production of exopolysaccharides based on the value of its isoelectric point. Keywords: Biodegradation; Catobolic gene; Denaturing gradient gel electrophoresis; Hexadecane INTRODUCTION Petroleum hydrocarbons are major pollutants of marine environments as a result of terrestrial and freshwater runoff, refuse from coastal oil refineries, off shore oil production, shipping activities and accidental spills (Arulazhagan et al., 2010). Bilge oil is the waste oil that accumulates in small quantities inside the lower spaces of a ship. Illegal discharge of bilge oil into the sea accounts for about 10 % of the total oils that enter the sea and oil pollution is a serious threat to the ecosystem. Bilge wastewater generally includes lubricating oil, cleaning diesel oil, oily sludge, spills from engine room, water leaks from internal pipes and sea water filtrations (Nievas et al., 2006). Hydrocarbons, the main constituent of bilge oil waters are toxic in nature and they are of concern due to their long persistence in the environment. Bioremediation is an attractive and a better alternative process for treating contaminated sites with the above mentioned compounds than the conventional methods like incineration, thickening, stabilization, dewatering, drying and disposal. Biological treatment can be defined as the natural ability of living microorganisms to reduce, remove or transform organic and inorganic pollutants. The primary objective of biological treatment is immobilization or stabilization or degradation of contaminants by microorganisms that is safe, effective, economic and alternative to traditional methods of remediation of the contaminants. The success of bioremediation is dependent on the inherent biodegradability of the pollutant, the accessibility of the pollutant to degrading microorganisms and on the optimization of biological activity (Yousefi Kebria et al., 2009). The Exxon Valdez and other similar incidents demonstrated the usefulness of bioremediation as a complete solution to such contaminations as it can be self sustaining and inexpensive (Margesin et al., 2003). In nearly all contaminated environments, there is a diverse group of microorganisms present with potential ability to degrade hydrocarbons. Enumeration of such hydrocarbon degrading bacterial populations by conventional cultivation dependent methods is time consuming and underestimates the numbers due to the inability to cultivate the majority of organisms. Molecular ecological studies showed that less than 1% of the environmental microbial diversity could be covered by known cultivation techniques (Torsvik et al., 2002). If the microbial diversity of the environment could be monitored, the direction and rate of processes catalyzed by environmental microbial communities may become better understood and controlled (Kirk et al, 2004). Hydrocarbon degrading microorganisms usually exists in very low abundance in marine environments. Identification of the key organisms that play roles in pollutant biodegradation is important for understanding, evaluating and developing in situ bioremediation strategies.Thus it is highly essential to characterize bacterial communities, to identify responsible degraders and to elucidate the catalytic potential of these degraders. The aim of this study was to isolate a hydrocarbonoclastic bacterial strain from bilge oil contaminated sea water and investigate its properties and its ability to degrade hydrocarbons. It is necessary to study the indigenous microorganisms capable of degrading pollutants because of their varied effects on the environment (Jain et al., 2005). Moreover, the shift in microbial communities from bilge oil contaminated water with that of its enrichment culture was investigated. This Research work was conducted within July 2008 and July 2010 at Bioremediation department, Centre for Environmental Research, Leipzig, Germany and Department of Biological Sciences, Birla Institute of Technology and Science Pilani, Goa, India. MATERIALS AND METHODS All the chemicals and reagents used in this work were of highest purity available. Sample collection Bilge oil contaminated sea water was collected from Marmugao harbour, Goa, India. After collection the samples were transferred immediately to laboratory. Oil was extracted from the sample and column chromatography was run to separate aliphatic and aromatic fractions. The aliphatic and aromatic fractions were quantified by gas chromatography. Microcosm design Microcosms were established in 100 ml conical flasks containing 20 mL of sea water amended with nutrients (20 mg/L NH4NO3 and 10 mg/L KH2PO4). Bilge oil contaminated sea water was taken in a separatory funnel and kept without disturbing for one hour. As the two layers got separated, the sea water from bottom was taken for microcosm studies. Anthracene was added as the carbon source at a concentration of 20 mg/L. Sea water + nutrients + anthracene + mercuric chloride (300 mg/L) served as the killed control and Sea water + nutrients + anthracene served as the biotic microcosm to study the degradation of anthracene by the microbial community present in the sea water. The microcosms were incubated at 30 °C on an orbital shaker (100 rpm). Degradation of anthracene was followed every week for three weeks. Enrichment on hexadecane and anthracene Enrichment cultures were established with either anthracene (20 mg/L) or hexadecane (1 %) as the sole carbon source in erlenmeyer flasks (250 mL) with 50 mL BH medium. After three subcultures, the cultures were plated on BH medium with hexadecane as the substrate. As the flask with anthracene showed no visual turbidity this sample was discarded. Two bacterial strains grew on BH medium with hexadecane as the sole carbon source and one of the dominant strain was used for further studies. The isolated bacterial strain was tested for its ability to degrade tetradecane, hexadecane or octadecane. Genomic DNA extraction Genomic DNA extraction from the bilge oil contaminated sea water as well as from enrichment culture and sea water from a different location (Chennai) was done by Yeates et al. 1998 method. Briefly 1L of bilge oil contaminated water and other samples were filtered through 0.22 µm pore size sterile polycarbonate membranes (Millipore). After filtration, the filter membrane was folded, immersed in High temperature electrolysis (HTE) buffer (50 mM Tris HCl, pH 7.5, 20 mM EDTA) and resuspended in 2 mL of lysis solution (0.15 M NaCl, 0.1 M Na2EDTA [pH 8.0]) containing 15 mg of lysozyme/mL, and incubated in a 37 °C water bath for 2 h with agitation at 20 to 30 min intervals and then 2 mL of 0.1 M NaCl-0.5 M Tris-HCl (pH 8.0)-10 % sodium dodecyl sulfate was added. Three cycles of freezing in a -70 °C dry ice-ethanol bath and thawing in a 65 °C water bath were conducted to release DNA from the microbial cells. After the freeze-thaw cycles, 2 mL of 0.1 M Tris-HCl (pH 8.0)-saturated phenol was added and the sample was briefly vortexed to obtain an emulsion. The mixture was centrifuged at 6,000 x g for 10 min. A 3 mL sample of the top aqueous layer was collected and then mixed with 1.5 mL of phenol and 1.5 mL of chloroform mixture (chloroform/isoamyl alcohol ratio, 24:1). A 2.5 mL portion of the resulting extract was further extracted with an equal volume of chloroform mixture. Finally, nucleic acids in the extracted aqueous phase (2 mL) were precipitated with 2 mL of cold isopropanol at -20 °C overnight. The pellet of crude nucleic acids was obtained by centrifugation at 10,000 x g for 10 min and then vacuum dried at 23 °C. The nucleic acid pellets, were resuspended in 100 µL of TE buffer (20 mM Tris-HCl, 1 mM EDTA [pH 8.0]). Polymerase chain reaction (PCR) for Denaturing gradient gel electrophoresis (DGGE) PCR amplifications of bilge oil contaminated sea water as well as from enrichment culture and sea water from a different location (Chennai) were conducted in 50 µL volumes containing 1X PCR buffer, 1.5 mM MgCl2, 0.5 µM of each primer, 200 µM of each dNTP, 0.6Uof Taq polymerase and sterile water. 1 µL of template DNA was used in each PCR. Negative controls with 1 µL of sterile water instead of target DNA were included in each PCR amplification. The primers used were 27 F and 1492 R (Naomi et al., 2009). These are universal eubacterial primers binding to phylogenetically highly conserved regions of the 16SrDNA. The PCR programme used was with an initial 95 °C denaturation step for 5 min, followed by 32 amplification cycles of denaturation for 30 s at 94 °C, 30 s at 52 °C and 1 min at 72 °C and also the final elongation step at 72 °C for 10 min. After successful amplification a 5 µL aliquot of the above PCR products were taken for a second PCR utilizing the 968 F primer with a GC clamp and the 1492R primers (Szekely et al., 2009). DGGE DGGE was conducted using the CBS Scientific system. Between 10 15 µL of PCR products were loaded onto 8 % (w/v) polyacrylamide gels (40 % acrylamide stock solution, 2 % bis solution 37.5 :1). 30-60 % gradient of denaturant (7 M urea and 40 % formamide v/v as 100 % denaturant (Nikolausz et al., 2008) was used to prepare the gel and the gel was run for 16 h in 1X Tris-Acetic acid-EDTA (TAE) buffer at 60 volts. The gel was finally stained with SYBR green. The stained gels were inspected under UV transillumination and pictures were taken using a digital camera. The central 1 mm2 portions of strong DGGE bands were excised with a sterile razor blade and soaked in 50 µL of sterile water overnight. A portion (15 µL) was removed and used as the template for PCR as described above to confirm the band ability to amplify. Purified DNA was sequenced with an automatic sequencer and the sequence identification was performed by use of the Basic local alignment search tool nucleotide (BLASTN) facility of National Centre for Biotechnology Information (NCBI). A mixture DGGE PCR products from different bacterial species is applied to the DGGE gel as a marker to check the electrophoresis run. Bacterial cell surface properties The cell surface properties of dominant bacterial strain from the enrichment culture was studied. The bacterial strain was grown on glucose or hexadecane as the sole carbon sources. The cells from both the samples were harvested almost at the same growth rate. Fatty acid differences, cell surface charge by measuring zeta potential was done as by Wick et al. (2003) and Neumann et al. (2006). Briefly, the isoelectric points of bacteria on both the substrates was determined from pH plots obtained by measuring µ in 10mM HNO3 - KNO3 solutions with pHs varying between 2 and 6.5 using an MPT-2 autotitrate (Malvern Instruments Ltd, Malvern, Worcestershire, UK). Bacterial lawns for contact angle measurements were prepared by collecting cell suspensions in 10 mM K NO3 on 0.45 µm pore size Micropore filters, mounting the filters on glass slides and drying them for 2 h at room temperature. Cells exposed to 1-decanol were washed 6 times with 10 mM K NO3. Cell surface hydrophobicities were derived from qw values for water drops on the bacterial lawns, using a DSA100 drop shape analysis system (Kruss GmbH, Hamburg, Germany). Uptake studies of 14C anthracene Hydrophobicity of the bacterial strain grown on hexadecane or glucose was also studied in the presence of 14C anthracene. The washed cell pellets of the bacterial strain grown on hexadecane and glucose at a concentration of 1 absorbance unit at 560 nm were suspended in 5 ml of BH medium with 20 mg/L of anthracene and 0.75 kBq/5 mL of 14C anthracene. Abiotic vials served as controls. Sodium azide was added at a concentration of 30 mM to all the samples to inhibit active transport (Bugg et al., 2000). After 1 h, 1 mL of samples in duplicate were removed from the experimental vials and centrifuged at 13000 rpm for 20 min and the amount of radioactivity partitioned into pellet was measured using Liquid scintillation counter (LSC) spectrophotometer. The pellet was resuspended in 1 mL of sterile distilled water and added to the scintillation cocktail for measurement. Quantification of hydrocarbons The total hydrocarbons were extracted from 100 mL of bilge oil contaminated seawater using three 50 mL aliquots of chloroform by solvent solvent extraction. The three 50 mL aliquots were pooled together and concentrated by rotor evaporator. The pooled sample was evaporated to 1 mL. The separation of aliphatic and aromatic hydrocarbons was done by column chromatography. This was done in a 25 cm chromatographic column filled with n hexane and 8 g of silica gel (Khan et al., 2005; Moreda et al., 1998). The aliphatic hydrocarbons were obtained in the first separation with 20ml of n-hexane. This was separated twice and the fractions were pooled together. The aliphatic hydrocarbon containing n-hexane was concentrated in a rotor evaporator to 1ml. The aromatic hydrocarbons were obtained in the separation with 3 x 20 mL of chloroform. All samples were analyzed by gas chromatography with Flame Ionization Detector (FID) detector. The analysis was performed with Chemito 2100 model equipped with a split splitless injector. The GC conditions are Oven 50 °C with a rise of 5 °C per minute till it reaches 290 °C. The injection temperature was 290 °C and the detection temperature was 300 °C. Detection of catabolic genes by PCR The hydrocarbonoclastic strain was confirmed for the presence of catabolic genes. The catabolic genes checked were C12O and C23O (Sei et al., 1999), pdo (Krivobok et al., 2003), p3-24(Dore et al., 2003), ndo (Teramoto et al., 2009), nidA (Brenza et al., 2003), phn (Jennifer et al., 2007) and all benzene,toluene, ethyl benzene and xylene (BTEX) catabolic genes as mentioned in Hendrickx et al. (2006). The reaction mixture (50 µL) contained 1X taq buffer, 2.5 mM MgCl2, 200 mM concentration of each deoxynucleoside triphosphate, 50 pmol of each primer, 2.5 U of Taq DNA polymerase (Sigma Aldrich, Mumbai, India) and 1 µL of DNA template. PCR ampliûcation was performed using a Gradient thermocycler (Eppendorf, Hamburg, Germany) and programs for each single reaction were followed as per literature. The veracity of the PCR products were checked by running it parallel with a 100 bp and 1 Kb ladder using 2% agarose gel in 1 X TAE buffer at 70 V for 1 h. RESULTS AND DISCUSSION The percentage of aliphatic and aromatic hydrocarbons in the sample collected was 43.16 % and 2.79 % respectively. Nievas et al. (2006) quantified total hydrocarbons in Bilge waste oily phase (BWOP) by gas chromatographic analysis. The total hydrocarbons present were 542 g/kg BWOP which includes resolved total hydrocarbons as well as Unresolved complex mixture (UCM). Aliphatics was found to be 89 % and aromatics was found to be 11%. The obtained results are in agreement with the analysis of Nievas et al. (2006, 2008) which higher concentration of aliphatics than aromatics in bilge oil waste. According to the observed kinetics of Nievas et al. (2008), three main types of hydrocarbons i.e. n-alkanes, biodegradable UCM and recalcitrant UCM are present in BWOP. Due to the presence of alkanes and biodegradable UCM, microcosm studies were conducted to study the potentiality of the native microorganisms in degrading a model hydrocarbon like anthracene. Microcosm studies Gas chromatographic analysis of microcosm studies revealed degradation of anthracene. The analysis showed degradation was significant after first week. There was abiotic loss of about 35 % of anthracene in three weeks and biotic loss of 55 % (Fig. 1) indicating presence of anthracene degrading bacteria in the bilge oil contaminated sea water. Active bacteria with ability to degrade three ring PAH anthracene is present in the bilge oil contaminated waters which was shown by the microcosm studies. Although the aromatic fraction of hydrocarbons is not high in the bilge oil contaminated waters, microcosm studies showed inherent ability of indigenous microcosm to degrade aromatic hydrocarbon like anthracene. This is of interest as hydrocarbon's complex molecular structure and low solubility in water limit the application of conventional remediation techniques (Santos et al., 2008). Enrichment studies Hydrocarbons are persistent compounds and thus a significant interest exists in studying and isolating microorganisms present in contaminated environments as a means for bioremediation (Haritash and Kaushik, 2009). The fate of hydrocarbons in the environment is associated with both abiotic and biotic processes which includes volatilization, photooxidation, chemical oxidation, bioaccumulation and microbial transformation (Cerniglia, 1992). The bilge oil contaminated water was enriched on hexadecane and anthracene so as to isolate bacteria that could utilize aliphatic and aromatic hydrocarbons. The enrichment culture grew on hexadecane. The enrichment culture when plated on Bushnell Haas medium agar plates with hexadecane gave two isolates which was found to be Pseudomonas mendocina and Ochrobactrum sp. by 16S rRNA sequence analysis. P. mendocina was used for further studies as it was the dominant strain. Growth was monitored by measuring absorbance at 560 nm. There was no increase in turbidity when anthracene was used as the substrate. According to Margesin et al. (2003) there is no correlation between prevalence of hydrocarbon degrading genotypes and the number of culturable hydrocarbon degrading microorganisms.P. mendocina could grow on different aliphatic hydrocarbons namely tetradecane, hexadecane and octadecane (Fig. 2a). Chromatographic analysis of residual hydrocarbons after inoculation with P. mendocina at 120 h showed about 73 % tetradecane, 54 % of hexadecane and 40 % of octadecane (Fig. 2b). Enrichment culture has long been the method of choice for isolating bacteria expressing specific phenotypes (Golovlev, 2001) and has been used successfully to isolate bacteria capable of degrading hydrocarbons. Harayama et al. (2004) emphasize there is little or no understanding of the vast majority of marine bacteria that remain uncultured and more efforts should be made to improve current methods for isolating oil degrading or oil emulsifying bacteria not only for assessing the fate and effects of the spilled oil but also for isolating novel bacteria that would be useful for the petroleum industry. DGGE analysis PCR-DGGE analysis was carried out on bilge oil contaminated sea water from Goa (Sample 1), enriched culture on hexadecane (Sample 2) as well as oil contaminated sea water from Chennai (Sample 3). Sewage sample was taken as Sample 4 just to compare the complexity of the diversity. Prominent bands from all the samples 1, 2 and 3 were excised and after PCR amplification were sequenced. Sequence analysis of the dominant 16S rRNA bands from the DGGE profiles revealed the differences between the genera between the bilge oil contaminated sea water and its enrichment culture on hexadecane (Fig. 3). Sample 1 showed two prominent bands (marked as 1 and 2 in the picture below), identified to be alpha proteobacterium and Marinobacter sp. Sample 2 showed one prominent band (marked as 3 in the picture below) which was identified to be Pseudomonas sp. which matched with the 16S rRNA sequencing of the dominant strain isolated from the enrichment culture. Sample 3 had one prominent band (marked as 4 in the picture below) which was identified to be Bacillus sp. These results indicate the shift in dominant bacterial strain depending on the type of substrate it is enriched upon. Enrichment on single hydrocarbons might lead to selection of different bacterial groups. Enrichment of bilge oil contaminated water on the type of hydrocarbon had a significant impact on the type of microbe enriched or on the microbial community structure. The cumulative result of the changes that occurred with the addition of specific hydrocarbons was selection of hydrocarbon degrading bacteria. According to Abed et al. (2002), almost all DGGE bands corresponding to field bacteria were replaced by other bands during the treatments demonstrating that there were dramatic shifts in the community structure. The total number of bands was higher for the controls than for the treated preparations, indicating that the model compounds inhibited the growth of some bacteria (Abed et al., 2002). McKew et al. (2007) showed that polycyclic aromatic hydrocarbons (PAH) degrading communities were distinct from those in microcosms containing alkanes.The bacterial communities within the hydrocarbon enriched microcosms were distinct from that of the initial seawater community (<10 % similarity), but community profiles from microcosms enriched with different classes of hydrocarbon were also distinct from each other. Catabolic genes Molecular approaches aiming to unravel the organisms and genes involved in in situ degradation are still needed (Jennifer et al., 2007). The isolated bacterial strain was tested for different catabolic genes. PCR products of expected size were obtained for strain carrying the corresponding target gene as shown in Table 1. The bacterial strain amplified for C23O, nid and ndo.The multicomponent enzyme system called naphthahlene dioxygenase initiates the metabolism of low molecular weight PAHs and is involved in the aerobic degradation of PAHs in the environment (Jennifer et al., 2007). Meta cleavage of catechol catalyzed by catechol 2,3-dioxygenase seems to be the most common pathway in the subsequent steps of PAH degradation. On account of the key position of the initial dioxygenase and the C23O during complete polyaromatic hydrocarbon mineralization, these genes seem to be suitable targets to detect the presence of PAH degradation potential at the DNA level (Haritash and Kaushik, 2009). Therefore the potential of the bacterial strain isolated i.e P. mendocina seems to be interesting as it will be able to degrade polyaromatic hydrocarbons as well. Physical properties The degree of saturation, water contact angle and zeta potential values (Table 2) indicate that the Pseudomonas mendocina is more hydrophobic when grown on water insoluble substrate. The isoelectric point for the bacterial strain was below 2.28 (Neuman et al., 2006) indicating the production of exopolysaccharides by the bacterial strain. Cells exposed to hexadecane showed significantly increased water contact angles qw = 93.88 ± 0.70 in comparison with qw = 77.59 ± 0.83 of cells growing in presence of glucose. According to the results, it is evident that there is negative correlation between cell hydrophobicity and surface charges. According to Kordel and Rinke (2001) the pollutants in the environment are commonly divided into a chemically available fraction, fraction available to the organisms (bioavailable fraction) and the fraction that is taken up by the organism (effective concentration). The success of bioremediation depends on the availability of pollutants to the microorganisms. Surfactants from microorganisms are of high interest as they increase bioavailability by providing components in readily accessible surfactant micelles or to disperse the organic phase and its interfacial area (Akhavan Sepahy et al., 2005). As there is indication of production of exopolysaccharides, care has to be taken as repeated growth in liquid batch cultures is highly selective for the planktonic mode of living and as a result the strains may lose the ability to excrete exopolysaccharides (Johnsen et al., 2007). Uptake studies Since the degree of saturation of the bacterial membrane was different in the presence of hexadecane and glucose, it was interesting to study the uptake mechanism of hydrocarbons using 14C labeled hydrocarbon. Anthracene was selected as the model hydrocarbon as the selected strain P. mendocina does not degrade anthracene. As hydrocarbons have to travel across bacterial membranes to reach cytoplasmic metabolic enzymes for mineralization, some authors suggested active uptake mechanisms of polyaromatic hydrocarbons (Kumar et al., 2007). As Sodium azide at a concentration of 30mM inhibits active transport but do not affect short term enzyme activity, it was used to inhibit mineralization of anthracene if at all. P. mendocina does not utilize anthracene but due to the presence of C23O gene, sodium azide was added as a preventive measure and studied the amount of anthracene uptake by hexadecane enriched and glucose enriched bacterial cells. The uptake of 14C anthracene is low with respect to glucose enriched cells than hexadecane enriched cells confirming the changes in hydrophobicity of the membrane when enriched on different compounds (Fig. 4). CONCLUSION In conclusion, the undertaken microcosm studies indicated the presence of anthracene degrading bacteria, however the isolation of anthracene degrading bacteria was not successful. The site seems to harbor phylogenetically diverse microbial populations that are able to tolerate and degrade complex hydrocarbon like anthracene. Furthermore, an efficient hydro-carbonoclastic bacterial strain was isolated from bilge oil contaminated sea water. The isoelectric point indicates production of exoploysaccharides by the isolated strain. The future parts of tht study focused on characterization of exopolysaccharides produced by the strain as well as its usage in treatment of bilge oil contaminated waters. ACKNOWLEDGEMENTS This study was done as a part of Department of Science and Technology, Government of India funded project (SR/FT/L-73/2005). The authors would like to thank Dr. Lukas Wick, Department of Environmental Microbiology, Helmholtz Centre for Environmental Research, Leipzig for helping us in physicochemical properties of the bacterial strain. The authors also thank German Academic Exchange Service (DAAD) for awarding two month fellowship under reinvitation programme to Dr. Mutnuri which helped a lot in this work. Authors also wish to thank "OCEAN SPARKLES, Goa" for providing Bilge oil contaminated water samples and Mormugao Port Trust for giving us the permission. REFERENCES
© IRSEN, CEERS, IAU
The following images related to this document are available:Photo images[st11042f1.jpg] [st11042t1.jpg] [st11042f3.jpg] [st11042f2.jpg] [st11042t2.jpg] [st11042f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}