 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
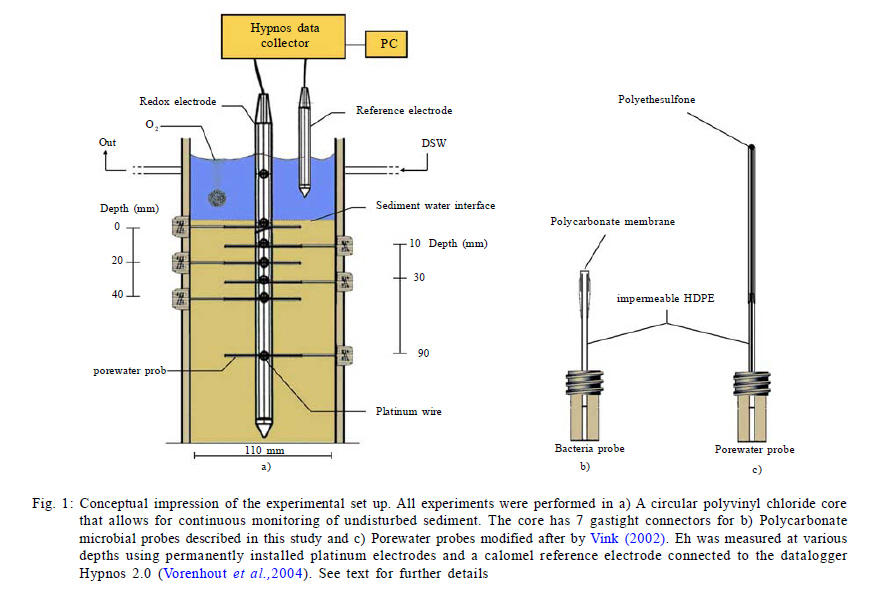
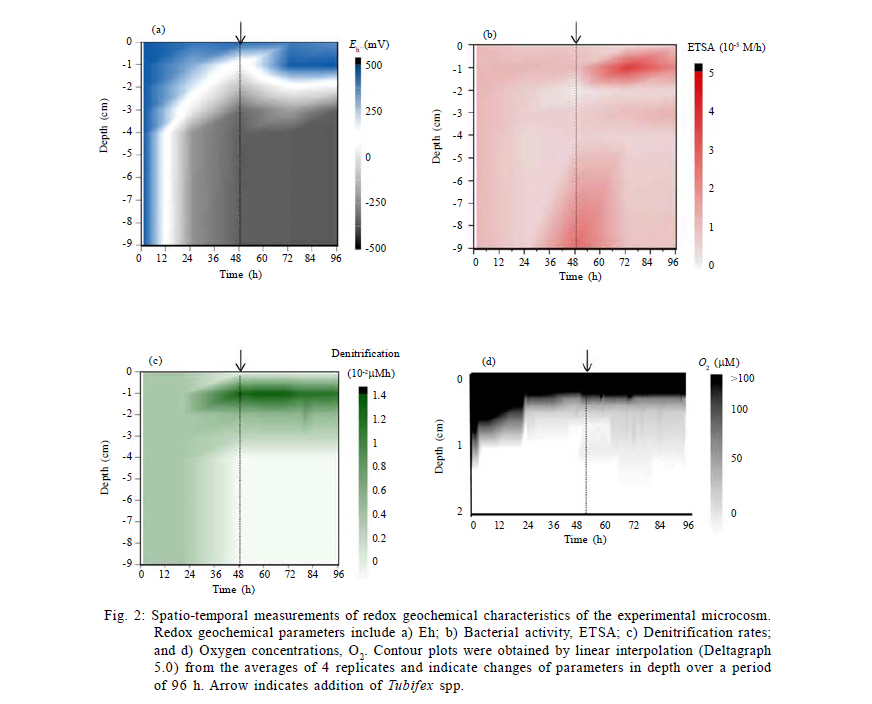
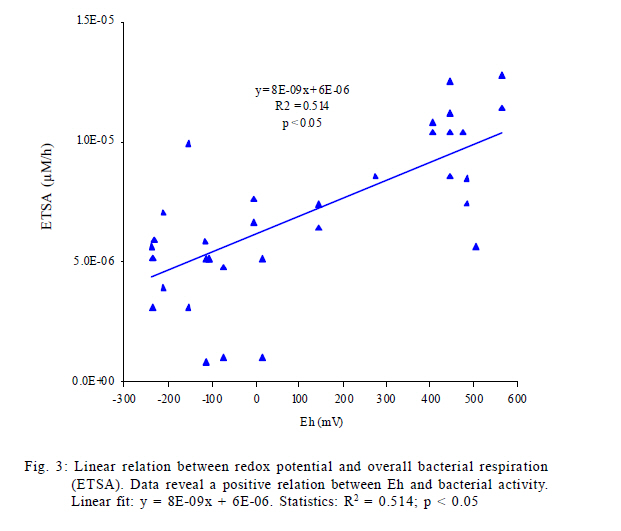
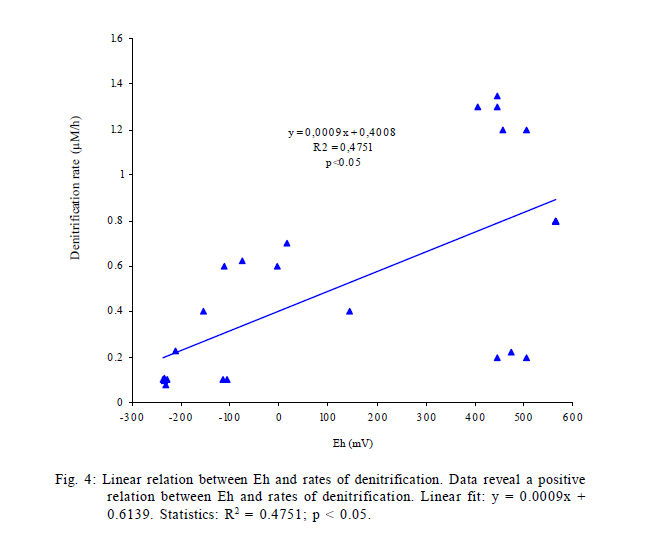
International Journal of Environment Science and Technology, Vol. 8, No. 3, 2011, pp. 553-560 Predictability of bacterial activity and denitrification in aquatic sediments with continuous measurements of redox potential *E. R. Hunting; H. G. van der Geest Institute for Biodiversity and Ecosystem Dynamics, Aquatic Ecology and Ecotoxicology, University of Amsterdam, Amsterdam, the Netherlands *Corresponding Author Email: e.r.hunting@uva.nl Tel./ Fax: +31205257659 Received 25 August 2010; revised 20 October 2010; accepted 27 February 2011 Code Number: st11051 ABSTRACT: Redox potential has been adopted as a qualitative parameter for interpreting solubility changes of nutrients and contaminants and the biological activity within wetland systems for several decades. The majority of studies considering the redox geochemistry in sediments used measurements of bulked material and single point measurement of biogeochemical parameters for interpretation, yet it remains questionable whether this information is reliable for environments that are very dynamic, such as wetlands. In this study it is evaluated whether variations in redox potential reflect dynamics of denitrification and overall bacterial respiration using continuous measurements of redox potential in time-series experiments in laboratory microcosms, in which the biogeochemical variation was enhanced by bioturbation. The results presented here suggest that measurements of redox potential have predictive potential in approximating rates of denitrification and overall bacterial respiration in aquatic sediments. The data clearly suggest that, while sediment bulk measurements and measurements of single profiles of redox potential, denitrification and bacterial activity often fail to provide ecological relevant information in dynamic systems, measurements of spatial and temporal redox potential profiles provide a useful parameter that reflects biogeochemical processes and functioning of sediments. Keywords: Biogeochemistry; Bioturbation; Ecosystem functioning; Tubifex; Wetland dynamics INTRODUCTION The redox potential (Eh) provides an electrochemical measure of a system's electric potential standardized as relative to a hydrogen electrode. Eh electrodes are alluring due to the rapidity and inexpensiveness of measurements and they are adopted as a qualitative parameter for interpreting solubility changes of nutrients and contaminants and the biological activity within wetland systems for already several decades (Jackson, et al., 1993; Teasdale et al., 1998; Biati and Karbassi, 2010; Biati et al, 2010; Mohiuddin et al., 2010; Sekabira et al., 2010). However, Eh is formally defined by the Nernst equation and thereby carries an erroneous assumption that redox reactions in a system are reversible, complete and in equilibrium, while many reactions in nature are neither complete or reversible. The incomplete and irreversible nature of many natural redox reactions in wetland soils and sediments may result in the occurrence of multiple redox `equilibria' and the measurement of a mixed potential. Different reactions are therefore expected to contribute differently to the signal, i.e. irreversible reactions fail to give a potential signal, while reactions that are kinetically slow may provide weak and unstable potentials, whereas other reactions may participate effectively in the measured signals (Grenthe et al., 1992). The interpretability of Eh relies on rigorous theoretical considerations and only a limited number of studies examined actual processes underlying Eh. Although the complexity of natural wetland systems generally flocculates predictability and interpretability, several studies were able to relate Eh to concentrations of terminal electron acceptors involved in electron transfer activity, in which Eh correlated well to concentrations of the redox species Mn (II) and Fe (II) and dominant redox processes (Brünger, 1982; Gao et al., 2002; Mansfeldt, 2004; Abdel-Ghani et al., 2009). However, the majority of studies considering the redox geochemistry in soils and sediments used measurements of bulked material and single point measurement without paying attention to temporal and spatial variability. Although this may provide information on underlying mechanisms in stable laboratory microcosms that are in quasi steady state conditions, it remains questionable whether this information is reliable for environments in which components that govern biogeochemical processes are highly variable, such as wetlands. Continuous monitoring of redox conditions with the use of permanently installed redox electrodes has demonstrated that in situ redox conditions may show large spatial and temporal fluctuations in a variety of habitats, where Eh may fluctuate from -400 to 100 mV over several days and may vary up to 200 mV in daily cycles (Vorenhout et al., 2004; 2011). This variability is easily overlooked in measurements of bulked soils and sediments and single profiles and it remains illusive whether parameters related to redox potential reflect similar spatiotemporal variability and whether this variability provides ecological relevant information on geochemical and bacterial processes in natural, dynamic environments. This study sets out to determine whether continuous measurements of Eh provide a key correlate to geochemical and bacterial processes in dynamic aquatic sediments, in which denitrification is under focus, since denitrification is a very active process and also a dominant, very important sink for mineralized nitrogen in Lake ecosystems and constructed wetlands (Saunders and Kalff, 2001). To this purpose, continuous redox potentials, rates of denitrification and bacterial metabolic activity were measured in time-series in microcosms containing sediment collected from Lake Drontermeer, the Netherlands, in 2007, in which spatial and temporal variability was induced by the introduction of the aquatic oligochaete Tubifex spp. MATERIALS AND METHODS Microcosm Organic rich sediment was collected from Lake Drontermeer, the Netherlands (52o58'N, 5o50'E) for natural bacterial communities and organic matter. The collected sediment was frozen in order to kill infauna and thawed to allow for bacterial re-growth. This approach generally does not have major impacts on bacterial community structures (Morly et al., 2005; Männistö et al., 2009). Natural sediments were subsequently mixed with clean river sand (ignited quartz; grain size 0.5 1.0 mm; ratio 1:5), homogenized and oxygenated prior to experimentation. Microcosm consisted of a circular polyvinyl chloride core (110 mm diameter, 250 mm height) with 7 gastight connectors at different depths for pore water probes modified after Vink (2002) and bacterial probes as described below. The pore water probes consisted of a semi permeable polyethersulfone polymer of which its mesh size (~0.1 µm) allows sampling of dissolved compounds in pore water but is impermeable to sediment particles and bacteria (Fig. 1). The probes were flushed with nitrogen gas before use to prevent oxidation processes during sampling. Bacterial probes consisted of a Nuclepore polycarbonate film mounted on polymer HDPE tubes by polymerization of vinyl chloride dissolved in 1,1,2-trichloroethane (Fig. 1). The membrane acts as semi-permeable membrane (mesh size 5 µm) to sample porewater containing bacterial cells while retaining particles and floccules. Sterile 3 mL syringes were connected to the probes for sample extraction. Cores were incubated at 20 ºC under continuous light conditions (40 ìmol photons/m2/s). Overlying water was kept at 30 mm above sediment water interface with continuous inflow of oxygen saturated water (Dutch Standard Water, DSW, a standardized synthetic analogue of common Dutch surface waters, containing 200 mg CaCl2·2H20, 180 mg MgSO4·7H20, 100 mg NaHCO3, and 20 mg KHCO3 per liter demineralized water: pH 8.1; hardness 210 mg/L CaCO3; alkalinity 1.2 meq/L; flow rate 20 mL/h). Overlying water was oxygenated, in which oxygen levels were generally stable at 70 % saturation over the course of the experiment. Experimental procedure Time series experiments were performed to link short term variability in Eh to different biogeochemical processes. The experiment consisted of two parts. First, the replicate (n=4) cores were allowed to stratify for 48 h, after which Tubifex spp.(approximately 6500 individuals/m2) were added to rework the sediment for an additional 48 h. Measurements of parameters in cores commenced immediately after homogenization and continued towards the end of the experimental runs at fixed time-intervals over a period of 4 d. Eh was measured every 15 min and oxygen profiles of the cores were measured twice per day. Pore water was simultaneously sampled alongside the platinum electrodes 1-2 times per day and immediately analyzed for rates of denitrification and overall bacterial respiration. Measurements of biogeochemical processes in this study are restricted to denitrification since denitrification is a very active process, and a dominant, very important sink for mineralized nitrogen in Lake ecosystems (Saunders and Kalff, 2001; Nouri et al., 2010). Analytical techniques Dissolved oxygen concentrations were measured with an oxygen microsensor (Unisense OX25) connected to a picoammeter (Unisense, PA2000). Eh was measured at various depths in the sediment (0 mm; 10 mm; 20 mm; 30 mm; 40 mm; 90 mm) using permanently installed platinum electrodes and a calomel reference electrode connected to Hypnos 2.0 datalogger (Fig. 1)(Accuracy of measurements ± 10 mV). Construction of electrodes and datalogger is described in detail elsewhere (Vorenhout et al., 2004). Eh was corrected for standard hydrogen electrode by adding 245 mV. Concentrations of nitrate in porewater (200 µL porewater sample) were colorimetrically determined (Shimadzu, 1601-UV) using Griess reagents (Griess, 1864). Decreases in concentrations of nitrate (NO3-) over a defined time interval provide rough estimations of rates of denitrification (Anderson, 1977). It should be noted, however, this approach is used to address the predictive potential of Eh and accurate estimations of denitrifications rates were beyond the scope of this study, provided that the approach is very likely to overestimate the actual process (Seitzinger, 1988). Overall respiratory capacity of the bacterial community was approximated by dehydrogenase enzyme activity via the reduction of 2-(p-iodophenyl)- 3-(p-nitrophenyl)-5-phenyl tetrazolium chloride (INT) to formazan (INTF). INT successfully competes with NADH+ and NADPH+ and INT reduction proved an accurate assay of Electron transfer system activities (ETSA) under both anaerobic and aerobic conditions (Smith and Mcfeters, 1997; Bhupathiraju et al., 1999; Rozzi and Remigi, 2004; Hunting et al., 2010). Bacterial cells were simultaneously sampled alongside the platinum electrodes (400 µL porewater sample). The samples were vortexed and incubated at 20 ºC for 30 min. Bacterial cell integrity was subsequently disrupted by 5 min sonication at room temperature (Branson, 1510). An aqueous INT solution (200 ìL; 6.2·10-2 M) was added and the samples were mixed and incubated for 1 hour at 20 ºC under dark room conditions. Enzymatic activity was stopped by adding 500 ìL mL of acetone and measured spectrophotometrically at 490 nm (Shimadzu, 1601-UV). An INT formazan standard was used for calibration, in which two protons equal the transfer of a single electron and samples were corrected for abiotic reduction of INT by subtracting formalin treated replicates. RESULTS AND DISCUSSION Homogenized sediments generally reached quasi steady state redox condition after 36 to 48 h (Fig. 2). Redox geochemical characteristics are provided in figure 2a-d. Contourplots represent the average of four replicates and show the changes in the parameters in depth over a period of 96 h. Eh ranged -300 to 500 mV at all depth measured in the core (Fig. 2a). Oxygen generally penetrated to 10 mm depth. Oxygen concentrations were highest at the sediment surface (237 ìM) and rapidly decreased after a few millimeters (Fig. 2d). Rates of denitrification ranged 0 1.35 ·10-2 ìM/h and were highest at 1 cm depth at steady state conditions (Fig. 2c). Overall bacterial activity (ETSA) arranged from 0 to 5·10-5 M/h. Bacterial activity was generally most pronounced at 1 cm depth and often decreased with increasing depth in deeper layers of the sediment, yet was occasionally enhanced at 4 cm and 9 cm depth in some of the replicates (data of replicates not shown), which is visible as slightly increased colour intensities at these depths in the contourplots of the averaged bacterial activity (Fig. 2b). Bacterial cells were simultaneously sampled alongside the platinum electrodes (400 µL porewater sample). As a consequence, the activity of the bacterial fraction sorbed to sediment particles is overlooked. However, it is consistently reported that the sorbed fraction fails to contribute significantly to measurements of bacterial activity (Bright and Fletcher, 1983; Bonin et al., 2001; Yousefi Kebria et al., 2009; Zhang et al., 2011). Addition of Tubifex spp. at t = 48 h resulted in a delayed increased penetration of dissolved oxygen (Fig. 2d) and increased rates of denitrification and bacterial activity in deeper areas of the sediment (Fig. 2b and c). It is well established that Tubifex spp. increase heterogeneity of the sediment through selective feeding and burrowing activity (Wavre and Brinkhurst, 1971; Van de Bund et al., 1994; Rodriquez et al., 2001; Kaonga et al., 2010). Mechanical reworking of sediments principally influences biogeochemical processes, as burrowing activity enhances irrigation and allocates terminal electron acceptors to deeper layers of the sediments (Robbins et al., 1979; Chakrabarty and Das, 2006; Nwuche and Ugoji, 2010). This enhances the redox potential, which, in turn, create redox conditions at which nitrate is more stable and dissimilatory nitrate reduction is allowed. An increased heterogeneity after addition of the aquatic oligocheate Tubifex spp. was visible in the spatio-temporal measurements of Eh, denitrification and bacterial respiration (Fig. 2b and c), coinciding with an increased oxygen penetration depth (Fig. 2d), suggesting that biogeochemical characteristics in studied microcosms evolved in a comparable fashion and were tightly coupled. Continuous measurements of Eh thus reflected the spatial and temporal heterogeneity of biogeochemical processes in studied microcosms. Performing linear regression on measured variables reveals a positive relation between redox potential and the overall bacterial respiration (Fig. 3) and rates of denitrification and redox potential (Fig. 4). Although some studies were able to correlate Eh with concentrations of nitrate in water bodies and soil (Brünger, 1982; Gao et al., 2002; Mansfeldt, 2004), the majority of studies have difficulties obtaining relations of Eh with bacteria-mediated geochemical processes. In the set up, only ~50 % of the variations in the measured rates in bacterial respiration and rates of denitrification are explained by redox potential. The lack of accuracy is most probably the result of the spatial resolution of measurements (10 mm increments), which unlikely captures fluctuations that may occur at millimeter and micrometer scales relevant to the microenvironment of the platinum electrodes, and this proved a critical issue in many experimental approaches in studying underlying mechanisms of Eh (Kappler et al., 2005). It is anticipated that correlations will improve if analytical sensitivity is enhanced. Since environmental redox reactions fail to behave in a pure Nernstian fashion, it is impossible to derive absolute quantitative information on redox processes from Eh measurements. For this reason it has been proposed that Eh measurements may at best be used qualitatively to describe sediments and soils as either oxidizing or reducing (Gao et al., 2002), or semi-quantitatively by considering critical Eh ranges sensu Mansfeldt (2004), in which highly oxidizing conditions provide little information due to the lack of electro-active redox systems, i.e. the low electron exchange density of oxygen at the platinum surface, while the lower levels of Eh can be used as an indication as to whether specific species are present or absent and which reactions dominate. Despite both theoretical and practical concerns regarding the interpretability of Eh, the redox potentials measured with permanently installed electrodes are positively related to rates of denitrification and bacterial respiration in the microcosms. Concordantly, continuous measurements of Eh were recently positively related to Cu-availability in artificial streams (Van der Geest and Leon Paumen, 2008). Although these results are correlative and the parameters were not exhaustive, the emerging patterns clearly reveal a predictive potential of Eh and encourage a closer look into the correlative nature of Eh with other biogeochemical processes. Evidently, since Eh subsumes a variety of biogeochemical reactions, which may be typical to specific systems (e.g. sulphate reduction is more common in marine sediments compared to freshwater sediments), prior knowledge on dominant processes in the system of interest is required to obtain predictability in Eh. If this information is available, Eh may provide a low cost and rapid measure that reflects biogeochemical processes and functioning of sediments. CONCLUSION This study aimed to evaluate the quantitative interpretability of Eh in relation to underlying processes within spatially and temporally dynamic aquatic sediments. The results presented here suggest that measurements of Eh have predictive potential in approximating rates of denitrification and overall bacterial respiration in aquatic sediments. The presented data clearly suggests that, while sediment bulk measurements and measurements of single profiles of Eh and bacterial activity likely fail to provide ecological relevant information, the use of continuous measurements of Eh provide a useful parameter that reflects biogeochemical processes and dynamics of sediments. ACKNOWLEDGEMENTS The authors express their gratitude to Michel Vorenhout and Girly Kelly for assistance and useful discussions. REFERENCES
© IRSEN, CEERS, IAU The following images related to this document are available:Photo images[st11051f2.jpg] [st11051f1.jpg] [st11051f4.jpg] [st11051f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}