 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
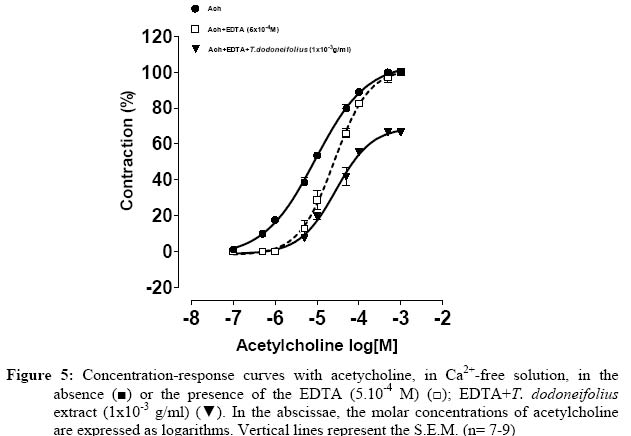
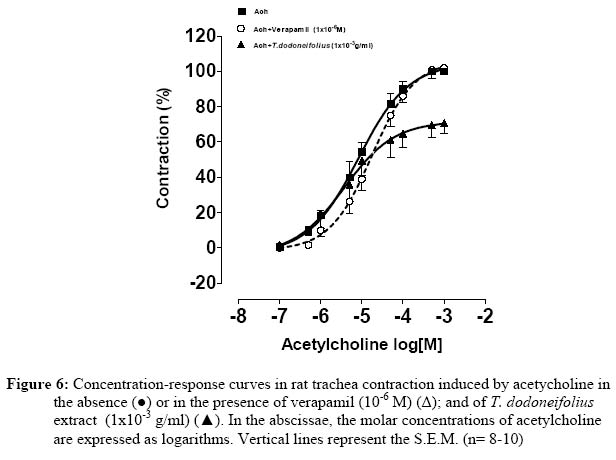
Research Paper PHARMACOLOGICAL EVALUATIONS FOR THE RELAXANT EFFECT OF THE HYDROALCOHOLIC EXTRACT OF TAPINANTHUS DODONEIFOLIUS ON RAT TRACHEA OUEDRAOGO1* Maurice, OUEDRAOGO2 Sylvin, OUEDRAOGO1 Lamini, TRAORE2 Aristide, BELEMTOUGRI1 G. Raymond, SAWADOGO1 L. Laya et, GUISSOU2,3 I. Pierre.
1UFR/SVT,Université de
Ouagadougou, 03 BP 7021 Burkina Faso. 2Institut
de Recherche en Sciences de la Santé (IRSS/CNRST), BP 7192, Ouagadougou,
Burkina Faso. 3UFR/SDS
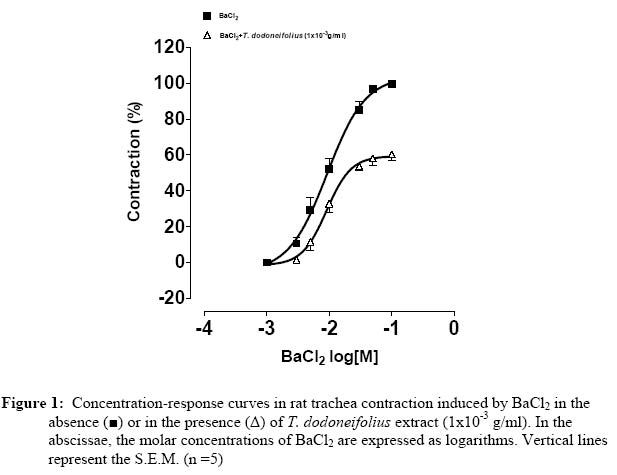
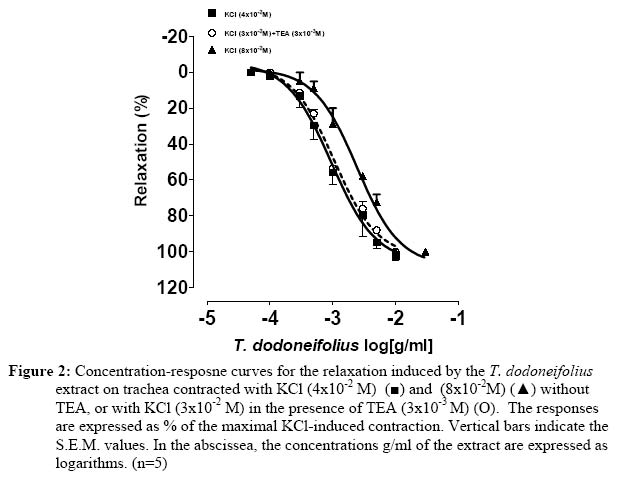
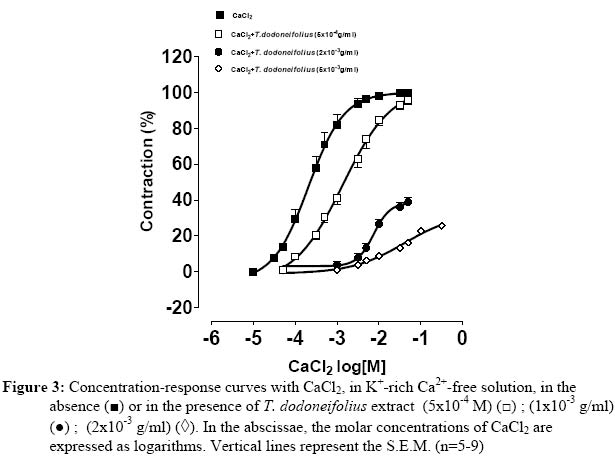
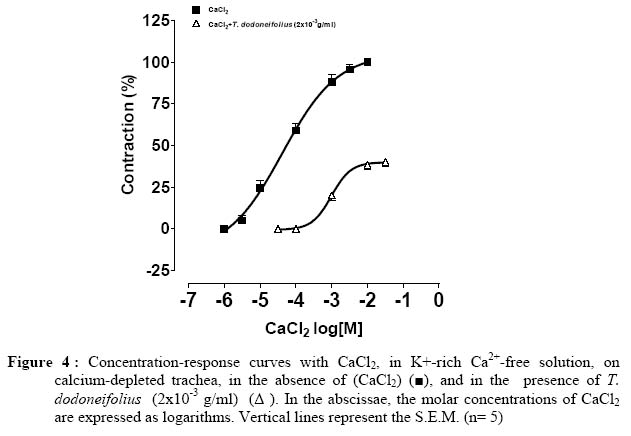
Université de Ouagadougou, 03 BP 7021, Burkina Faso. Code Number: tc05018 Abstract The present study was designed to investigate the blocking of calcium by the hydroalcoholic extract of Tapinanthus dodoneifolius (Tapidod), "in vitro", on rat trachea. To evaluate this effect, the contractile activity of tracheal chains from Wistar Kyoto rats was isometrically recorded. On the isolated tracheal rings the extract produced the following effects: (a) a reduction of the contraction obtained by BaCl2, (b) a bronchorelaxing action, on strips precontracted by KCl, which was not influenced by TEA (3x10-3 M), (c) a concentration-dependent decrease of the spasm evoked by calcium chloride (CaCl2) in K+-rich Ca2+-free physiological salt solution, before and after intracellular calcium depletion (d), an inhibitory effect on contraction induced by acetylcholine in Ca2+-free Krebs-Heinseleit solution supplemented with EDTA (5x10-4M). It is concluded that: 1. The activation of the potassium channels does not play a significant role in the relaxant effect of Tapidod. 2. The antispasmodic property of Tapidod seems to be mediated by the blockade of intracellular Ca2+ release. 3. Most likely an inhibition of the intracellular Ca2+-regulating proteins is involved. Keys words: Extract of Tapinanthus dodoneifolius (Tapidod); Trachea; Calcium. Introduction Asthma is a worldwide chronic airway inflammatory disease, the etiology of which is not known (Holgate, 1997). The mortality and morbidity associated with this disease have not appreciably declined (Sly and O'Donnell, 1997), and its treatment has not changed over the last two decades (Wenzel, 1998). Therefore, research has not reached its goal, and the treatment of asthma is still seeking for targeted, anti-inflammatory therapy (Vaali, 1999). Phytotherapy research represents a promising approach to the discovery of biologically active molecules that may be beneficial in the treatment of asthma. Tapinanthus dodoneifolius is extensively used as medicinal plant in traditional medicine in Burkina Faso. According to Boussim (2002) Tapinanthus dodoneifolius is used in the treatment of cholera, asthma and diabetes. The aqueous extract is taken against gynecologic disturbances, digestives disorders and nervous confusions (Nacoulma/Ouédraogo 1996). Cardiovascular properties of T. dodoneifolius have been recently described by Ouédraogo et al. (2005a). Cepeleanu et al. (1994) revealed larvicidal and molluscidal effects of T. dodoneifolius. Preliminary pharmacological investigations on rat trachea indicate that Tapinanthus dodoneifolius total aqueous extract inhibited contractile activity induced by physiological and non-physiological agonists (Ouédraogo et al., 2005b). Chemical analyses of different extracts from T. dodoneifolius yielded components as triterpenes, sterols, carotenoïdes, saponosides, anthracenosides, anthocyanosides and tannins (Traoré, 2000). The present study was conducted to analyze comprehensively the relaxant effect and to elucidate the mechanism of action of Tapidod in rat trachea. Material and Method Plant and extracts Tapinanthus dodoneifolius (DC.) Danser (syn: Agelanthus dodoneifolius (DC.) R.M. Polhill& D. Wiens) was collected in Loumbila (Burkina Faso) in April 2002 and taxonomically identified by an expert botanist (Prof Boussim I. J., University of Ouagadougou). A voucher specimen is deposited in the Herbarium of the Department of Vegetal Biology, University of Ouagadougou, Burkina Faso, with the reference no. 002. The drying operation was carried out under room temperature with out exposure to sunlight. Air-dried and powdered plants of T. dodoneifolius (100g) were soaked in chloroform (1000 ml) for 24 hours. After filtration, the remainder was dried at room temperature and extracted in ethanol (1000 ml) for 24 hours. The extract was centrifuged, concentrated by rotary evaporation, frozen and lyophilized. Tissue preparation Tracheal rings were dissected from either male or female Wistar rats (weighing 200-300g) following ethyl carbamate anesthesia and exsanguination by carotid artery transection. Thoracic tracheas were removed rapidly and cleaned of surrounding tissue. The isolated tracheas were cut as spirals according to the method described by Brunelleschi et al. (1987); Hazekamp et al. (2001) and suspended in 20 ml organ bath chambers containing modified Krebs-Henseleit physiological solution at pH of 7.4, continuously oxygenated and maintained at 37° C. The rings were mounted by means of two parallel L-shaped stainless steel holders inserted into the lumen. The upper holder was attached with thread to a transducer while the lower one was tied to a stationary glass rod. The preparations were allowed to equilibrate for at least 90 min while the base-line tension was adjusted progressively to 2g as recommended by Tschirhart et al. (1987). Isometric contraction of the smooth muscle was recorded with chart recorder via a force-displacement transducer (Grass FT 03C). Experimental procedures Acetylcholine and barium chloride are tested by cumulative addition, directly in the tissue bath. The total amount of drugs added did not exceed 5 % of the total volume (1ml) of the bath (Van Rossum, 1963). For the determination of the effects of antagonists on the response to the addition of agonists, tissues were incubated in the presence of antagonists 15 or 30 min prior to the addition of agonists. To investigate the involvement of voltage-sensitive Kchannels activation in Tapidod-induced relaxation, two K+ channel blockers (BaCl2 and TEA) (Emine et al., 1998; Imaizumi and Watanabe, 1981) and the method of depolarization described by Karaki and Weiss (1988) were used. In order to study the inhibition of the availability of the calcium by Tapidod three different experimental protocols were applied. These protocols derived from method described by (Lagaud et al. (1996) In the first experimental condition, concentration-response curves to CaCl2 were constructed by cumulative application in a K+-rich Ca2+-free solution. Trachea was previously incubated in a normal physiological salt solution (equilibration period) and depolarized in the K+-rich Ca2+- free solution. The K+-rich Ca2+- free solution was obtained by KCl (8x10-2 M) substitution for an equimolar amount of NaCl, omission of calcium and addition of EDTA (5x10-4 M) (Guan et al. 1987). In the second experimental protocol which intended to study the component of the Tapidod-induced relaxation due to the blocking of the Ca2+ influx, the first experimental condition was repeated after depletion of intracellular Ca2+. Depletion was obtained by repeated exposure of the trachea to acetycholine maximally active concentration (10-3M), in a Ca2+-free solution containing 2mM EDTA , until there was no response to acetylcholine. To study the component of the Tapidod-induced relaxation due to the inhibition of internal Ca2+ release, an intracellular Ca2+ mobilizing agent (acetylcholine) was used on rat trachea in Ca2+-free solution. Acetylcholine was added cumulatively to the bath. A washout period of 45 min was allowed between each experimental protocol. Expression of results The bronchodilatory efficacy of the extract was expressed as Emax=maximal bronchodilatory response, evaluated as % relaxation of the KCl-induced contraction. The potency of the plant extract was expressed as pEC50 = -log EC50 (Jenkinson et al. 1995), calculated as concentrations (g of extract per ml of organ bath) required to produce 50% of Emax (Computer program: GRAPH PAD PRISM® 2.01). Drugs The Krebs-Heinsleit solution had the following composition (millimolar): NaCl, l18; KCl, 4.75; CaCl2, 2.5; KH2PO4, 1.2; MgSO4 7H2O, 1.2; NaHCO3, 25 and Glucose, 5.55. Acetylcholine chloride and verapamil were obtained from Sigma. Tetraetylammonium chloride was provided by Fluka. Ethylene Diamine Tetraacetic Acid (EDTA), BaCl2, CaCl2 and KCl were purchased from Prolabo. All the drugs were prepared as concentrated stock solution and diluted to the final concentration in Krebs-Heinseleit solution. In preparing the stock solution, all the drugs were dissolved in distilled water. Before each experiment the lyophilized extract was dissolved and diluted in distilled water. Statistical analysis Results are expressed as means±SEM of number (n) of experiments. The significance of differences was evaluated by means of ANOVA and student' s t test for unpaired data. P values lower than 0.05 were considered to be significant. Results Tapidod effects on the concentration-response curve of BaCl2 BaCl2 (10-4M to 10-1 M) produced a concentration dependent contraction on rat trachea (pEC50=2.03±0.06). Tapidod reduced significantly the contraction induced by BaCl2 and caused a rightward shift of the concentration-response curve for BaCl2. In tracheal rings the maximal contractile response to BaCl2 in the presence of Tapidod (1x10-3g/ml) was 59.326±2.54 % of the maximal response observed in its absence (Figure 1). Effects of Tapidod on the KCl-induced contraction TEA (3x10-3 M) potentiated KCl-induced contraction. Tracheal rings, pre-treated by TEA, maximal response to 3x10-2 M KCl did not differ significantly from 4x10-2 M KCl-induced contraction without TEA. This equivalence in developed tension has been determined by comparing KCl-induced contraction concentration-response curves in the absence or in the presence of TEA (3x10 M). Tapidod elicited a bronchodilatory effect on rat trachea pre-contracted by KCl (pEC=3.02-350±0.13, E=106max.2±15.22%, in absence of TEA; pEC=2.9750±0.075, E=102.9max±5.33% in presence of TEA)(Figure 2). In tracheal rings precontracted by KCl 8x10-2M, the extract produced a concentration-dependant bronchorelaxant response (pEC=2.6150±0.07 and E=107.8max±7.02%). Tapidod effects on the concentration-response curve of CaCl2 in k+ rich Ca2+-free solution In trachea bathed by K+-rich Ca2+-free physiological salt solution, CaCl2 (10-5M to 10-2 M) evoked concentration-dependent spasm (pEC50=3.68±0.06). When the effect of Tapidod (5x10-4, 2x10-3, 5x10-3 g/ml) was evaluated on CaCl2 cumulative concentration-effect curve a dose-dependant depression of the curve was detected in the high K+ Ca2+-free solution (pEC50=2.86±0.07 and Emax=103.3±5.09%; pEC50=2.13±0.06 and Emax=38.67±2.11%; pEC50=1.38±0.16 and Emax=33.11±5.14%, respectively) (Fig. 3). Calcium depletion did not influenced Tapidod (2x10-3g/ml) antispasmodic effect (pEC50=3.01±0.06, Emax=39.96±2.34%) (Figure 4). Tapidod effects on the concentration-response curve of acetylcholine in Ca2+ free solution. Calcium removal from the bath shifted to the right the concentration-response curve for acetylcholine but did not influenced Emax. Tapidod (1x10-3g/ml) inhibited the effect of acetylcholine in Ca2+-free solution. (pEC50=5.041±0.04 in the absence of Tapidod; pEC50=4.55±0.05 and Emax= 69.40± 2.12% in the presence of Tapidod; Figure 5). Effect of Verapamil and Tapidod on acetylcholine dose-response curve The contraction induced by acetylcholine was markedly inhibited by the pre-treatment with Tapidod (pEC50=5.041±0.04 in the absence of Tapidod; pEC50=5.43±0.12 and Emax=71.68±2.76% in presence of Tapidod (1x10-3 g/ml)) (Figure 6). However, in the presence of verapamil (1x10-6M), acetylcholine concentration-effect curve was not significantly influenced (pEC50=4.75±0.06 and Emax=106.02±2.33%). Discussion The presence of Voltage-dependant K+ Channels, Ca2+-dependant K+ channels and ATP-dependant K+ channels in tracheal smooth muscle has been reported by Kotlikoff et al. (1990); Marthan et al. (1989) and Black et al. (1990). Barium, a K+ channels blocker (Thirstrup et al., 1997), induced concentration dependant contractile activity inhibited by Tapidod. The blocking of potassium conductance increases the transmembrane Ca2+ influx as a result of the cell membrane depolarization (Corrompt et al., 1998), eliciting a contractile response. It has been suggested that the activation of K+ channels reduces the voltage-dependant Ca2+ influx and [Ca2+]i through tonic hyperpolarisation of smooth muscle cells (Kannan and Johnshon, 1995). As Tapidod inhibits the tracheal smooth muscle response to the blocking of K+ conductance , we suggest that this extract may act by activating the K+ influx or by modulating the Ca2+ influx. Moreover, one way to study a possible K+ channel opener is to increase the Ke to 40 mM level (Vaali, 1999). Tapidod relaxed 80 mM K+-induced contraction indicating that its bronchorelaxing action is not mediated by a K+ channels activation. Tapidod prevented the contraction induced by Ca2+on rat tracheal muscle bathed by K+-rich Ca2+-free physiological salt solution. Intracellular calcium depletion did not influence Tapidod effect. These inhibitory effects indicate a blocking of the Ca2+-influx. L-type Ca2+ channel blocker (Verapamil, 1x10-6 M) did not reverse acetylcholine-induced contraction. This result is similar to those mentioned by Foster et al. (1984) and Advenier et al. (1984). However, T. dodoneifolius antagonized acetylcholine indicating that the pharmacodynamic profile of Tapidod differs from that of the L-type Ca2+ channels blockers. In Ca2+-free solution Tapidod produced a depression of the contractile response to acetylcholine . Eglen et al. (1994) and Marthan et al. (1987) reported that acetylcholine elicits contractile activity by using calcium from internal stores. One characteristic in smooth muscle is that pharmacomechanical coupling can be seen even when extracellular Ca2+ concentration is zero and must therefore depend upon intracellular stores. The depression of the contractile response to acetylcholine in Ca2+-free Krebs solution suggests an action on the intracellular mobilization of Ca2+ ions. The two most important factors affecting smooth muscle contraction are [Ca2+]i and the contractile machinery's sensitivity to calcium (Vaali, 1999). As the precise mechanism of action of Tapidod remains to be elucidated, the present results suggest that its inhibitory action may involve its binding to intracellular Ca2+-regulating proteins (i.e. calmodulin) thereby preventing Ca2+ from binding to these proteins. Similar mechanisms have been reported by Hazekamp et al. (2001) to explain the bronchodilatory effect of Clerodendrum petasites (Verbenaceae) on guinea-pig trachea. Conclusion It is beyond the scope of this paper to speculate further on the structure-activity relationship. Tapidod is a crude extract that would contain several constituents able to present additional effects. The purpose of this paper was to demonstrate the Ca2+-blocking activity of Tapidod, which relaxes KCl-induced contraction on rat trachea and inhibits agonists-induced contraction. It was concluded that Tapidod effect may involves an intracellular calcium sequestration or/and a calcium-regulating proteins inhibition. Acknowledgements This work was supported by grants from IFS. References
© Copyright 2005 -African. Journal. Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc05018f1.jpg] [tc05018f3.jpg] [tc05018f2.jpg] [tc05018f6.jpg] [tc05018f5.jpg] [tc05018f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}