 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal. Traditional, Complementary and Alternative Medicines Vol. 2, Num. 3, 2005, pp. 222-232 Research Paper ACUTE TOXICITTY AND SLEEP-WAKE EEG ANALYSIS OF STACHTARPHETA CAYENNENSIS (VERBENACEAE) IN RODENTS *M. A. Akanmu1,2, G. Olayiwola2, O. E. Ukponmwan2, K. Honda1 1Department
of Biosystem Regulation, Tokyo Medical and Dental University, Tokyo, Japan, 2Department
of Pharmacology, Faculty of Pharmacy, Obafemi Awolowo University, Ile-Ife,
Osun-state, Nigeria, 3Department of Physiological Sciences, Obafemi
Awolowo University, Ile-Ife, Osun-state, Nigeria Code Number: tc05023 AbstractThis study was carried out in order to establish the scientific basis of the reported ethno-medicinal use of Stachytarpheta cayennensis (Verbanaceae) as sedative drug. The acute toxicity and barbituric-hypnotic test of the total aqueous (TASC), 50% ethanol (SCCR) and butanol (SCBT), ethylacetate (SCEA) fractions of dried leaves of S. cayennensis were studied in male rats and mice. The effect of systemic administration of TASC on sleep architecture in rats was also evaluated in Sprague-Dawley rats that were chronically implanted with electrodes for electroencephalogram (EEG) and electromyogram (EMG) recording. The acute toxicity test revealed no lethal effect with doses of SCCR (up to 2000 mg/kg) per oral in rats and doses of TASC (up to 1000 mg/kg and 95 mg/kg) systemically in rats and mice respectively. The extracts significantly (p<0.01) potentiated phenobarbital-induced hypnosis by reducing sleep latency and increasing sleeping time in a dose-dependent manner. Systemic nocturnal administration of single doses (250 and 500 mg/kg) 30 minutes before the onset of dark period of TASC significantly (p<0.05) increased total sleep time, time spent in non-rapid eye movement (NREM) sleep during the 6-h recording period while its effect on REM sleep was biphasic with the higher dose administered. The aqueous extract of Stachytarpheta cayennensis showed profound effects on sleep-wake stages suggesting the potential sedative effects. Key words: Stachytarpheta cayennensis; acute toxicity; barbituric-hypnotic test; Wake; NREM sleep; REM sleep Introduction Plants of the family Verbenaceae have been used traditionally as tonic, anticonvulsant and sedative. It has also been reported in the treatment of diabetes, cold and dysentery, among other uses (Duke, 1994; Schmeda-Hirschmann and Rojas, 1992). Stachytarpheta cayennensis also known, as bastard/false vervain is a member of this family used as a stomachic and febrifuge, and to treat chronic liver diseases (Burkill, 1966; Alice et al., 1991; Comerford 1996; Schwontkowski, 1993). Previous studies showed the chemical constituents of the total extracts contain alkaloids (Alice et al., 1991), ipolamide, 6ß-hydroxyipolamide and Verbascocide (Kooiman, 1975; Schapoval et al., 1998). The iridoid, ipolamide and phenylethanoid glycoside aceteoside were isolated from the active fraction and showed to have inhibitory effect on histamine and bradykinin-induced contractions of guinea pig ileum (Schapoval et al., 1998). The aqueous extract of the plant is reported to possess antiulcer activity (Vela et al., 1997). Most of the ethnomedicinal claims of the plants have not been investigated. Therefore, in this study, we investigated the acute toxicity, sleep-wakefulness EEG analysis, barbituric-hypnotic effects of Stachytarpheta cayennensis. Materials and MethodsPlant materials The leaves of Stachytarpheta cayennensis were collected from the campus of the Obafemi Awolowo University, Ile-Ife. The plant was identified by Adesakin and authenticated by Prof. O. Olorode and Dr. H.C. Illoh of the Department of Botany of the Obafemi Awolowo University. A voucher specimen of the plant samplewas deposited in the herbarium of the Forestry Research Institute of Nigeria, Ibadan. The voucher specimen number is (FHI-106491). Preparation of extracts and fractions The total aqueous extract (TASC) of S. cayennensis was prepared by the method of infusion. The infusion was prepared by steeping the air-dried, moderately coarse powder of the leaves of the plant in boiling water for 10-15 minutes in a conical flask with a close-fitting lid before straining to obtain the filtrate. The aqueous extract was prepared to the required concentrations of 200 mg/ml for pharmacological studies and 50 mg/ml for the sleep EEG analysis. For the 50% ethanol extract (SCCR), 1 kg leaves were air-dried, powdered and extracted cold with EtOH/H2O (1:1) for 48 hours. The marc was re-extracted twice and the combined extracts were concentrated in vacuo to yield 88.52g (8.8%) SCCR. The SCCR was successively partitioned into ethylacetate and n-butanol and similarly concentrated to give EtOAc (SCEA, 19.20g, 21.69%) and BuOH (SCBT, 35.52g, 40.13%). These extracts and fractions were used for the various scientific studies. Animals Male Sprague-Dawley rats (350-544g) obtained from Crea Japan Inc. (Tokyo, Japan) and mice of either sex (32-40g) (Vom strain, Nigeria) were used for the study. The animals were kept on a 12-h light and 12-h dark schedule (lights on 8:00 h and lights off 20:00 h) in a constant air-conditioned room at 25 ± 1OC with relative humidity 60 ± 6%. They had free access to standard laboratory pellet diet (CLEA Rodent diet CE-2, Tokyo, Japan and Standard commercial diet, Ewu, Nigeria) and water. All experimental procedures were in accordance with animal research guidelines. Drugs The drugs used for the experiments in this study are: Diazepam (Roche, Basle, Switzerland), flumazenil (Ro 15-1788, Sigma, St. Louis, USA), penicillin G. potassium (Meiji, Tokyo), phenobarbital sodium, (Abbot Lab; Illinois, USA). Acute Toxicity Test and LD50 Determination The total aqueous extract (TASC), the extract from the 50% ethanol extract (SCCR) and fractions from SCCR (i.e. SCEA and SCBT) were tested on rats and mice for acute toxicity. Graded doses of the extracts and fractions of the leaves of S. cayennensis were administered orally (p.o.) and intraperitoneally (i.p.) into the animals (rats and mice) in the treatment groups. Rats were administered with single doses of 250, 500 and 1000 mg/kg body weight intraperitoneally while mice were given (5-100 mg/kg) intraperitoneally and 50-250 mg/kg body weight by oral administration. The control groups received normal saline only either by oral or intraperitoneal route. Each group of animals has six to eight rats/mice (Dietrich, 1983; Irwin, 1962). The behavioral changes of the animals after the intraperitoneal administration of TASC, SCCR, SCEA and SCBT were observed for 1 hour (rats) and 30 minutes (mice) continuously and intermittently for 6 hours (Irwin, 1962; Gámel et al., 2000) by placing the animals individually into the observation cage. The animals were also observed for toxic symptoms and mortality for 72 hours. The number of animals that died within the period of study was noted for each group and this was used to calculate LD50 (Hayes, 1989). Effect of Extracts on Barbituric-Hypnosis Test The mice in the treatment groups received 10-50 mg/kg, i.p. of extracts or standard drug (Diazepam, Roche, 3 mg/kg) intraperitoneally. The animals in the control groups received equal volume of normal saline. After 30 minutes, all animals received i.p. injection of phenobarbital sodium (40 mg/kg). The onset of the loss of righting reflex and the duration of the loss of the righting reflex were noted. In another experiment, mice were treated with flumazenil (Ro 15-1788, Sigma, 3 mg/kg i.p.) 15 minutes before SCCR (50 mg/kg i.p.). After 30 minutes, all animals received phenobarbital sodium (40 mg/kg i.p.). The control animals received the SCCR without flumazenil, and phenobarbital sodium 30 minutes later. The time elapsed between the loss and voluntary recovery of the righting reflex was recorded as sleeping time and the sleep latency time as the time before the loss of postural reflexes (de Paris et al., 2000). Effect of Extract on Sleep-Wakefulness Male rats were implanted with electroencephalogram (EEG) and electromyogram (EMG) electrodes under general anaesthesia with phenobarbital sodium (50 mg/kg, i.p.) in accordance with standard procedure (Akanmu et al., 2004, Honda and Inoué, 1981, Rhee Y.H. et al., 1990). The rats aged 60-70 days were placed in a stereotaxic apparatus, secured with blunt rodent ear bars. The heads were shaved and the skin retracted, revealing the skull. Holes were drilled to allow stereotaxic implantation of the reference, auxiliary and working electrodes. Three cortical gold plated miniature screws (Ø, 0.9mm) served as epidural EEG electrodes for recording EEG while two nuchal stainless-steel electrodes were also implanted for recording EMG. The electrodes were soldered to a plug and chronically fixed over the skull by a dental acrylic resin. Thereafter, each animal received a total of 40,000 U of penicillin G. potassium (Meiji, Tokyo) subcutaneously and locally around the incision. One week was allowed for recovery from surgery while the animals were housed in a special experimental cage, which allowed for continuous monitoring of EEG and EMG along with locomotor and drinking activities (Honda and Inoué, 1981, Rhee et al., 1990). The experimental cages were placed in a soundproof electromagnetically shielded, air-conditioned room. Lead wires of the electrodes were connected to an EEG/EMG amplifier (Nihon-Kohden multichannel Amplifier MEG-6116) via a slip ring (Airflyte Electronics Co. CAY-675) fixed above the cage allowing free movement of the rats. The amplifier was connected to a PC with an AD converter and software (Sleep Sign, Kissei Comtec, Matsumoto, Nagano, Japan) for acquiring and processing data. Data was sampled at 128Hz and subjected to on-line spectral analysis by Fast Fourier Transformation (FFT) over 8-s epochs. Subsequently, data were stored in a magnetic optical (MO) disk that was later visually analyzed offline after auto analysis of the sleep-wake stages. After observing the establishment of circadian rhythms in sleep-wake behavior, the rats were subjected to experiments to obtain baseline data starting at the onset of dark period (Honda et al., 1981). Thirty minutes before the onset of dark period, distilled water (vehicle) or TASC single doses of 250 and 500 mg/kg were administered intraperitoneally and a continuous EEG/EMG recording was done for 6-h. The data were scored on the computer in 8-s intervals and the time spent in wakefulness, NREM and REM sleep were calculated for each recording hour (Honda et al., 1994). Sleep-wake stages were divided into non-REM sleep, REM sleep and wakefulness based on the recordings. Non-REM sleep was determined by a low frequency, high amplitude EEG and a low EMG tone. REM sleep was a high frequency low amplitude EEG and a silent EMG tone but with transient episodes of muscle twitching. Wakefulness was determined by a high frequency, low amplitude EEG and a high EMG tone. The average total sleep time (TST) duration and the frequency of waking, NREM sleep and REM sleep episodes were calculated for each 2-h period, and for the 6-h recording period. The values obtained from the administration of extract on experimental days were compared with values from the baseline recording. Statistical analysis All values are quoted as the mean ± S.E.M. Statistical analysis was done using the student’s t-test for the barbituric-induced hypnotics test and Repeated measures ANOVA followed by Student-Newman-Keuls test for the sleep EEG analysis. *P < 0.05 was considered significant. Results Acute toxicity tests and LD50 determination Oral administration of SCCR (500 - 2000 mg/kg) produced no toxic effect or mortality in rats. 50-250 mg/kg of the extracts administered to mice per oral also did not produce any mortality. Intraperitoneal administration of TASC (250-1000 mg/kg) did not produce any lethal effect; however, SCCR (5-100 mg/kg) produced toxic effect and mortality at doses above 90 mg/kg body weight in mice. Aqueous extract of leaves of S. cayennesis produced effects on gross behavior and decreased spontaneous motor activity and palpebral ptosis was also observed. Barbituric-hypnosis test The intraperitoneal administration of SCCR (10 - 50 mg/kg) decreased the sleep latency and prolonged the sleeping time due to phenobarbital significantly in mice (Table 1). In addition, the n-butanol fraction (SCBT) also increased significantly (p<0.001) the sleeping time induced by phenobarbital sodium compared with the control. The results obtained with n-butanol fraction (SCBT 5 mg/kg: 160.0 ± 5.6 min; SCBT 10 mg/kg: 171.0 ± 4.1 min; 20 mg/kg: 140.0 ± 3.6 min; 40 mg/kg: 115.0 ± 4.6 min, Diazepam 3 mg/kg: 170.0 ± 4.4 min; control: 49.0 ± 2.5 min, n=5). The results also showed that flumazenil (3 mg/kg i.p) significantly reversed the effect of the extract (SCCR) at the highest dose administered, resulting in increased sleep latency (4.2 ± 0.2 min) and a decreased sleeping time (63.7 ± 4.1 min, Table 1). Table 1: Effects of different doses of 50% ethanol extract of Stachytarpheta cayennensis (SCCR) on hypnotic effects in mice and influence of treatment with flumazenil (FMZ: 3mg/kg, i.p.) on the hypnotic effect induced by SCCR (50 mg/kg). Each value represents the means ± SEM (n=5). Student’s t test, *p<0.05 versus control; #p<0.05 versus FMZ. SCCR, 50% Ethanol extract; FMZ, Flumazenil
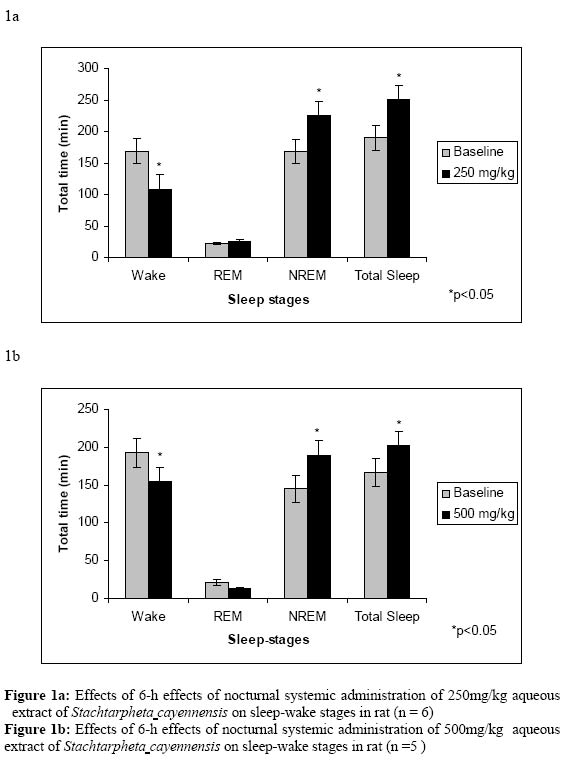
Effects of extract on sleep-wakefulness The effects of total aqueous extract (TASC) on the mean (±S.E.M) of the total amount of wake, NREM sleep and REM sleep in 2-h intervals and corresponding baseline (control) are presented in Table 2. The results show that after intraperitoneal injection of 250 mg/kg TASC, the total duration of NREM sleep significantly (p<0.05) increased in all time intervals scored except during the first 2hr time block, showing the sleep continuity or stabilization effect of the plant (Table 2). Similar results were observed with the dose of 500 mg/kg during the second 2-h interval and significant tendency (p=0.05) during the third 2-h interval (Table 2). Table 2: 2-H time intervals analysis of nocturnal systemic administration effects of total aqueous extract (TASC) of Stachtarphetacayennensis on sleep-wake stages.
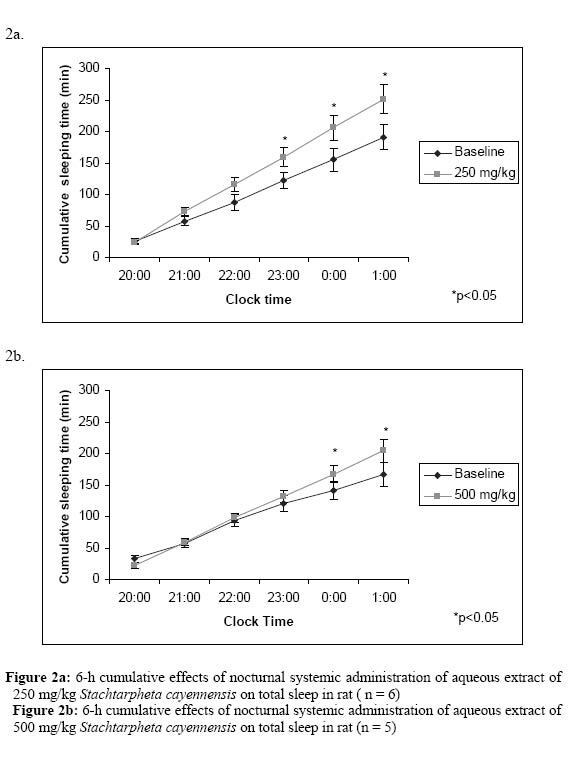
In considering the total recording period (6-h), the total duration of NREM sleep was significantly increased throughout the recording period (Figures 1a & b), however, TASC showed biphasic effect on REM sleep, thus showing dose-dependent effects on REM sleep. REM sleep duration was increased due to administration of 250 mg/kg of TASC while it was decreased by the administration of higher dose of 500 mg/kg (Figures 1a & b). These results reflected the biphasic effect of this plant on sleep-stages. Data analysis of 2-h time intervals also showed similar results on REM sleep. Our results show that during the dark period, which is the active period for rats, the total sleep time was significantly increased during the entire period of EEG recording (Fig. 1a & b). The two doses of TASC (250 mg/kg and 500 mg/kg, i.p) administered significantly increased total sleep period (repeated measures ANOVA, 250 mg/kg: treatment effect +33.0 % (p<0.05); 500 mg/kg: treatment effect +21.0% (p<0.05). These effects were due to an increase in episode duration. Moreover, the cumulative hourly amounts of total sleep were mostly above the baseline levels for both doses especially after the administration of the lower dose of 250 mg/kg (Fig. 2a & b). The results generally show that TASC significantly decreased wakefulness during the experimental period with concomitant increase in sleep. (Figure 2) Discussion and conclusion The results of the acute toxicity tests show that the 50% ethanol extract (SCCR) and total aqueous extracts (TASC) are safe, as they produced no toxic effects or mortality when administered to the test animal via the oral route of administration. Systemic administration of TASC also produced no toxicity, however, the intraperitoneal route for SCCR extracts produced toxic effects and mortality. LD50 for the extract using the intraperitoneal route is 90 mg/kg for mice. The safety of the extracts via oral administration correlates with the prescribed mode of using the plant for the different ailments i.e. aqueous or alcoholic oral infusion. The behavioral profile of rats under the influence of the extract induced a decrease in locomotor activity and increase passivity. The administration of 1000 mg/kg intraperitoneally induced profound sedative effects that the animals did not even respond to noxious mechanical stimuli in contrast to the response given by control animals. SCCR and SCBT decreased the sleep latency and increase the sleeping time due to pentobarbital. The effect of the extracts on the sleeping time induced by pentobarbital was significantly different from the control group. The butanol fraction (SCBT) produced effects that are similar to the crude extract (SCCR) but SCBT is effective at low concentration of 5 mg/kg while SCCR is ineffective at this dose. The potentiation of the barbiturate-induced sleep further suggests that S. cayennesis possesses some sleep inducing properties. The observed palpebral ptosis during the acute toxicity study lends further support for the sedative properties of the aqueous extract. Sleep, is a basic necessity of life and it is as important to our health and well-being as air, food and water. It is related to brain and mind functions through reducing cerebral metabolism (Maquet, 1995). We need to have continuous sleep (≈ 6-h sleep) that becomes restorative and results in feeling refreshed and alert. During sleep, we pass through different states and these stages of sleep are more likely to be experienced with continuous and uninterrupted sleep. This “sleep architecture” follows a predictable pattern of rapid-eye movement (REM) and Non-REM (NREM) sleep. Both states are important to experiencing quality sleep and getting the right mix and enough of both helps to maintain the natural sleep architecture that can lead to restful and restorative sleep. In Particular, NREM sleep seems to be associated with energy conservation through down regulation of body temperature and metabolic rate (Benington et al 1995). In the EEG sleep–wakefulness analysis, the two doses employed (250 and 500 mg/kg i.p) enhance NREM sleep stage especially during the third 2-h block interval after the administration of the aqueous extract. However, the extract showed a biphasic effect on REM sleep with administration of higher dose of the extract. REM sleep was significantly decreased during the second 2-h time interval with significant decrease in total duration. This shows similarity to the sleep effect of systemically administered modulator of Gamma-aminobutryic acid (GABA) receptors such as barbiturates, benzodiazepine, cytokines, zopiclone and neuroactive steroids, and the selective GABAAagonist (muscimol) that have been found to increase sleep continuity, promote NREM sleep and sometimes suppress REM sleep at higher dose (Lancel, 1999; Mckernan, 2000; Kubota et al., 2000, 2001). High dose of TASC inhibit REM sleep while simultaneously enhancing NREM sleep. Thus, our results also show that the leaves of S. cayennensis might be exerting its hypnotic effects via the GABAA receptors sites in a similar way as other GABAAagonists. Flumezanil reversed the effect of the extract on both sleep latency and duration of sleep. This observation lends credence to the assertion that the extract may be acting via the GABAA receptor for its sedative activity (Mckernan, 2000; File, 1986; Batatinha M, 1995). The enhancement of NREM sleep was due to an increase in the number of episodes (frequency) and prolongation of the episodes (duration). The cumulative hourly amount of NREM sleep and total sleep were mostly above the level of baseline recordings with the lower dose of 250 mg/kg. S. cayennensis induced profound effects on the sleep-wake stages and in this study it is clearly shown that this plant modulate both NREM and REM sleep stages. Interestingly, only the lower dose (250 mg/kg i.p.) enhanced both REM sleep (non-significantly; p=0.07)) and NREM sleep stages (significantly; p<0.05), whereas the higher dose (500 mg/kg i.p.) enhanced NREM sleep and influenced biphasic effect on REM sleep (Table 2). The aqueous extract and the butanolic fraction of Stachytarpheta cayennensis showed profound effects on sleep-wake stages suggesting the potential sedative effects of the plant. Thus, the results obtained from this study scientifically showed that the plant has sedative-hypnotic properties. References
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

{kind=link}
{kind=link}