 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
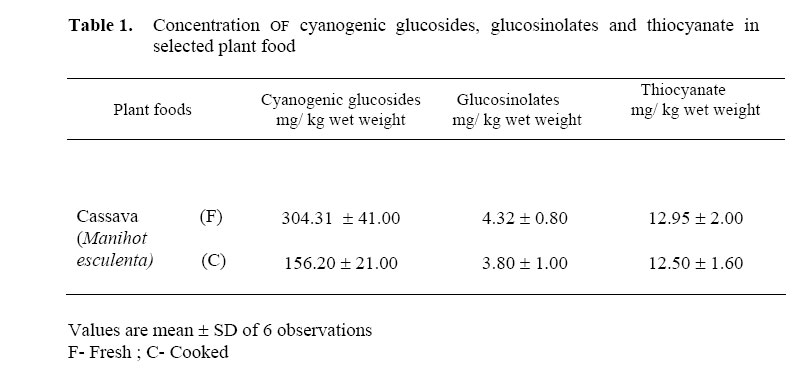
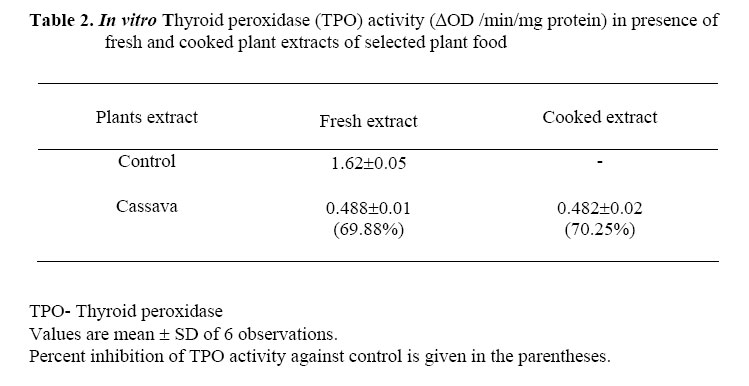
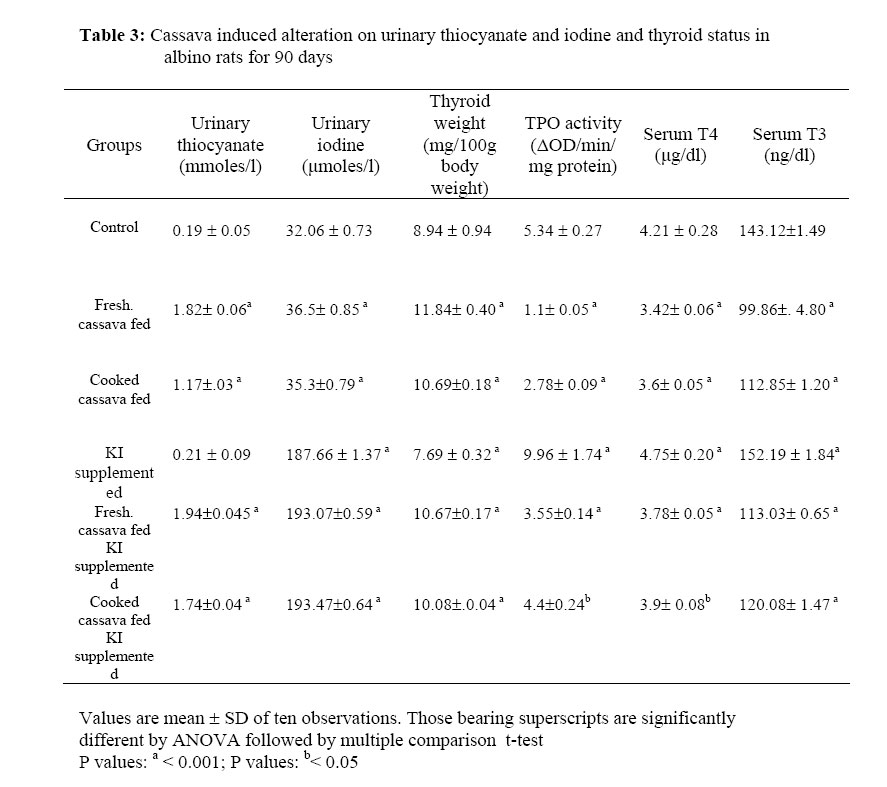
African Journal of Traditional, Complementary and Alternative Medicines, Vol. 3, No. 3, 2006, pp. 87-99 Research Paper EFFECT OF CASSAVA (MANIHOT ESCULENTA CRENTZ) ON THYROID STATUS UNDER CONDITIONS OF VARYING IODINE INTAKE IN RATS Amar K Chandra, Dishari Ghosh, Sanjukta Mukhopadhyay & Smritiratan Tripathy Endocrinology and Reproductive Physiology Laboratory, Department of Physiology, UniversityCollege of Science and Technology, University of Calcutta, 92, Acharya Prafulla Chandra Road, Kolkata 700009, India E-mail address: amark_chandra@yahoo.co.in, Phone : 033-2350 8386 Fax : 91-033-2351-9755 Code Number: tc06041 Abstract Cassava (Manihot esculenta Crantz), a widely used common vegetable of Indian origin contains dietary goitrogens. Goitrogenic content, in vitro anti-thyroidal activity and in vivo antithyroid potential of fresh and cooked cassava of Indian origin in albino rats with and without iodine supplementation were evaluated in this study. In vitro inhibition of thyroid peroxidase (TPO )activity was found with fresh and cooked cassava extract but the presence of extra iodide in the incubation media had shown reduction in such inhibition to an extent. Inhibition constant (IC50) and PTU equivalence of fresh cassava were 42.5 ± 1.35µg and 2.21 respectively.After consumption, cyanogenic constituents in cassava are metabolized to thiocyanate and iodine-retaining capacity seems to be dependent on thiocyanate exposure. In cassava fed rats, the urinary iodine concentration resembled the state of adequate iodine nutriture but thyroid gland fails to utilize available iodine. Increased thyroid weight (P < 0.001), inhibited thyroid peroxidase (TPO) activity and reduced thyroid hormone profiles (P < 0.001) were noted in fresh and cooked cassava fed rats, resembling a relative state of morphological as well as biochemical hypothyroidism even in presence of adequate iodine. Key words: Goitrogens, Iodide, Thyroid hormones, Thyroid peroxidase, Cassava Introduction Cassava (Manihot esculenta Crantz) is a major food source for a large part of the world population, particularly in tropics including India and it is the fourth most planted staple root crop after rice, wheat and cassava (Okezie and Kosikowski, 1982; Padmaja, 1995; Boby and Indira 2004). Cassava contains the cyanogenic glycoside linamarin (93%), 2-(β-D-glucopyronosyloxy) isobutyronitrile and a small amount of lotaustralin (7%) (Nartey, 1968; Barret et al. 1978) and the presence of these two cyanogenic glucosides in cassava is a major factor limiting its use as food or feed (Padmaja 1995). Hydrolysis of linamarin by endogenous β- glucosidase enzyme linamarase, present in cassava produces D-glucose, acetone and hydrocyanic acid (Conn, 1969). It is also hydrolysed in the intestinal tract of both man and animals by the microbial flora (Winkler, 1958). There are evidences suggesting large variation in goitrogen content in the same plant having same taxonomy but from different geographical regions (Michajlovskij and Langer, 1959). Except cyanogenic glucosides, cassava of Indian origin also contains glucosinolates and thiocyanate in its edible part (Chandra et al. 2004). Regular consumption of cyanogenic glucosides, glucosinolates and thiocyanate, the goitrogenic / antithyroid constituents of cyanogenic foods, disrupt the biosynthesis of thyroid hormones in several ways and may develop endemic goitre in long run (Delange et al. 1982). As there are only few experimental studies on cassava of Indian origin specially in the aetiopathogenesis of thyroid disorders, the present study was undertaken to evaluate the thyroid status after chronic consumption of cassava both as fresh and cooked under conditions of varying iodide intake in rats. Materials and MethodsCollection and processing of cassava Fresh cassava was collected from the local markets for the measurement of goitrogenic constituents, assay of in vitro TPO activity and also for feeding the experimental animals. i) Fresh or uncooked or plant food was prepared by coarsely chopping in a cutter and then incubated in an oven at 30-40oC for 4 h following the method of de Groot et al (1991). ii) Cooked plant food was obtained by boiling in an equal weight of water for 15 min. The cooked material was not drained and immediately used for preparing diets following the method of de Groot et al (1991). Animals Sixty Wister rats (80±5 g) were allocated to control and experimental groups of ten each. Animals were caged in unheated well-ventilated stainless steel cages and maintained in the laboratory on standardized normal diet (20% protein) which consisted of 70% wheat, 20% Bengal gram, 5% fish meal powder, 4% dry yeast powder. 0.75% refined till oil and 0.25% shark liver oil and water ad libitum (Chandra et al. 2004). Approval was provided for the study was given by the Ethical Committee of the Department of Physiology, University of Calculta. Animal treatment Control group of rats was fed normal laboratory diet. Fresh cassava fed group received fresh cassava replacing 1/3rd portion of normal laboratory diet. Cooked cassava fed group of rats were fed cooked cassava by replacing 1/3rd portion of normal diet. In potassium iodide (KI) supplemented group, rats were fed normal laboratory diet with extra iodide 12-14 μg/rat/day (Gaitan and Merino, 1976). Fresh cassava fed KI supplemented group of rats were fed both fresh cassava as well as extra iodide and the replacement of the normal diet was as above. Cooked cassava fed KI supplemented group of rats were fed with cooked cassava and also extra iodide as mentioned above. The animals were maintained with above-mentioned dietary regimen for 90 days. Feed consumption, corrected for feed wasted and body weight were measured every seven days. In the last week of the treatment, each group of animals was kept in metabolic cages for 24 h to collect the urine over xylene for the analysis of iodine and thiocyanate. At the end of the experimental period body weights of the rats were recorded and the animals were sacrificed at the end of the 90th day of experiment following ethical procedure. Just before sacrifice, blood samples were collected from each rat from the portal vein under ether anaesthesia and the serum was separated for the assay of thyroxine (T4) and triiodothyronine (T3) and kept at –20oC till analysis. Just after sacrifice, thyroid glands were weighed after removing connective tissues and used to assay thyroid peroxidase activity. Measurement of dietary goitrogens in cassava Cyanogenic glucoside It was measured following the method of Lambert et al(1975). Plant tissues were hydrolysed by the enzyme glucosidase (β-glucosidase) and the hydrocyanic acid thus liberated was trapped in sodium hydroxide. Cyanide content of trapped hydrocyanic acid was then determined quantitatively. Glucosinolates/thioglucosides The enzyme thioglucosidase reacts with thioglucosides present in plant producing thiocyanate. Following this principle thioglucosides was measured as per Gmelin and Virtanen (1960). The thiocyanate thus produced was estimated by the method of Aldridge (1945) as modified by Michajlovskij and Langer (1958). Thiocyanate The plant food was extracted with clean sand and water and refluxed subsequently. The extract containing thiocyanate was then allowed to react with benzidine hydrochloride and the intensity of colour thus formed was measured photometrically following the method of Aldridge (1945) as modified by Michajlovskij and Langer (1958). In vitro inhibitory effect of cassava on thyroid peroxidase activity To evaluate in vitro anti-TPO activity of cassavahuman thyroid tissue was collected from ENT Department, S.S.K.M.Hospital, Kolkata.Edible part of each fresh plant was homogenised in assay buffer (5 mg plant tissue in 5ml, pH 7.2,and 100mM phosphate buffer) and then centrifuged at 700 g for 10 minutes. After centrifugation, 50μl of aliquot of the supernatant of plant was added separately in a 1ml cuvette containing acetate buffer (pH 5.2, 50mM), potassium iodide (1.7 mM) and microsomal fraction of thyroid tissue. Freshly prepared hydrogen peroxide (0.3 mM) was added lastly to start the reaction to assay the TPO activity (∆OD /min/mg protein) under the influence of respective plant extract following the procedure of Gaitan et al (1989) modified by Chandra et al (2004). Anti-TPO activity of the plant extract in the above-mentioned condition was also studied in presence of excess potassium iodide. For this purpose, in the cuvette maintaining the same concentration of assay buffer, plant extract and H2O2, the concentration of potassium iodide was increased and it was found highest after adding 20μl of KI (3.4mM) and ∆OD /min/mg protein was recorded. Assay of IC50 and 6-n-propyl-2-thiouracil (PTU) equivalence The activity of raw plant extract was also evaluated in terms of the concentration necessary to produce 50% inhibition (IC50) of thyroid peroxidase activity. The effect of raw plant extract was studied at different concentrations ranging from 10μg to 150μg original fresh plant to determine the concentration required to produce IC50 of thyroid peroxidase activity. The TPO activity under the influence of the plant at a particular concentration, as a percentage of inhibition of the control value was plotted against the concentration of the original plant extract, and the concentration at which the 50% inhibition occurred (IC50) was determined from the plot. The IC50 value of plant given is the mean ±SD of 6 observations. To compare the relative anti TPO activity of the studied plant against a known antagonist, IC50 of 6-n-propyl-2-thiouracil (PTU obtained from Sigma Chemical Co.) was determined. Analysis of urine Estimation of iodine It was measured by dry ashing following the method of Karmarkar et al(1986). In this method, iodine content in urine sample was estimated by drying urine at 600oC in presence of potassium carbonate and the iodine present in the ash was measured by cerric-arsenite system. Estimation of thiocyanate It was measured following the method of Aldridge (1945) as modified by Michajlovskij and Langer (1958). Trichloroacetic acid was added to urine sample, mixed and centrifuged. To the supernatant saturated bromine water was added and 4% arsenic trioxide (As2O3) was then added to oxidise all bromine present in the sample. After that benzidine hydrochloride and pyridine mixture were added and the colour developed gradually. After 30 min optical density was measured at the wavelength of 525 nm. Measurement of thyroid peroxidase activity A 10% homogenate was prepared using thyroid tissues in phosphate buffer (pH 7.2, 100mM) and sucrose solution (500 mM) at 4oC. Homogenisation was carried out in a Potter- Elvehjem glass homogeniser for 45-60 s at 2000 rpm and about 15 strokes min–1. The homogenate was centrifuged at 1000 g for 10 min and this low speed supernatant was further centrifuged at 10,000 g for 10 min at 4oC to get the mitochondrial fraction. The microsomal fraction containing most of the peroxidase activity was obtained by centrifuging the post mitochondrial supernatant at 1,05,000g for one h. After centrifugation at1,05,000 g, the precipitate was solubilized in phosphate buffer. Thyroid peroxidase activity was measured following I3- formation from iodide using spectrophotometer (UV-1240 Shimadzu, Japan) at 353nm by the method of Alexander (1962). For performing the kinetic assay, in a 1ml cuvette, 0.9ml of sodium acetate buffer (pH 5.2,50mM), 10μl KI (1.7mM) and 20μl microsomal fraction of thyroid tissue containing 0.03-0.04 mg protein were added and the reaction was started by the addition of 20μl freshly prepared H2O2 (0.3 mM) according to laboratory standardization. The thyroid tissue protein level was determined by the method of Lowry et al (1951) using bovine serum albumin as standard. The results are expressed as ∆OD /min/mg protein. Enzyme linked immunosorbant assay of total circulating thyroid hormones Circulating thyroxine and triiodothyronine level was measured using Monobind, Inc. total T4 kit [kit no. MBI 32718/083001]. In this method serum sample (25μl) was taken in a microplate well and enzyme-T4 conjugate was added, then the reactants were mixed. After the completion of the required incubation period, the antibody bound enzyme thyroxine conjugate was separated from the unbound enzyme thyroxine conjugate by decantation. The activity of the enzyme present on the surface of the well was quantified by the reaction with tetra methyl benzidine (TMB) substrate solution with 15 minutes incubation and finally by adding 1N HCl as stop solution and absorbance was read against 450 nm wavelength in ELISA Reader [Merck]. Circulating T3 level was measured with 50 μl serum sample following the above mentioned method. Statistical Analysis All data were statistically analyzed and presented in the table as mean ±SD. Comparison among the groups was performed by ANOVA and the level of significance was expressed at P < 0.05and P < 0.001 after multiple comparison ‘t’ test. ResultsGoitrogen content Goitrogen content in edible portion of fresh and cooked cassava was measured. Cyanogenic glucosides, glucosinolates and thiocyanate content of fresh cassava were measured (Table 1). In the fresh cassava, the concentration of cyanogenic glycosides was found higher than its cooked counterpart. Glucosinolates concentration was reduced moderately in cooked cassava and thiocyanate in fresh and cooked cassava was found almost same. In vitro inhibitory effect of cassava on thyroid peroxidase activity TPO activity of control (10μl KI, 1.7 mM) in absence of plant extract was 1.62±0.054 ΔOD/min/mg protein. In vitro inhibitory effect of edible part of cassava on thyroid peroxidase activity was determined after application of fresh cassava extract with and without extra iodide in the incubation media (Table 2). Relative anti TPO potency The inhibition constant (IC50) and PTU equivalence of fresh cassava were 42.5 ±1.35μg and 2.21 respectively. Urinary thiocyanate, urinary iodine as well as thyroid weight, TPO activity and serum levels of total T4, T3 in control, cassava fed (both fresh and cooked), KI supplemented and cassava fed (both fresh and cooked) KI supplemented groups of rats treated for 90 days are shown in Table 3. Urinary thiocyanate concentrationRats fed with both fresh and cooked cassava showed significant increase in urinary concentration (P < 0.001) as compared to control. Of these two sets (one fed fresh cassava and another fed cooked cassava) thiocyanate excretion was higher in the fresh cassava fed group. Iodine supplementation increased the thiocyanate excretion in both the cassava fed groups in comparison to cassava fed iodine non-supplemented groups. Urinary iodine concentrationUrinary excretion of iodine was increased but not significantly higher in cassava fed rats than the control. Urinary excretion of iodine was more in iodine-supplemented groups of rats. Thyroid weight Significant increase in thyroid weight was observed in both the cassava fed rats and control groups (P < 0.001). The gain in thyroid weight was more in the group fed fresh cassava. The apparent gain in thyroid weight was more in both the groups fed cassava in different conditions than the cassava fed iodine supplemented groups but the differences were not significant. Thyroid peroxidase activityIn cassava fed rats, thyroid peroxidase activity was reduced significantly to that of their control group (P < 0.001). Between the two cassava fed groups i.e. one fed fresh and another fed cooked cassava TPO inhibition was more in the group fed fresh cassava. Supplementation of iodine in both the cassava fed groups had increased the TPO activity in comparison to that of only cassava fed groups (both as fresh and cooked) but failed to bring back its activity at control level. Serum T4 and T3 levelsTotal serum circulating T4 and T3 levels were determined in both groups fed cassava and compared with control groups. Total serum T4 and T3 levels were reduced significantly (P < 0.05 and P < 0.001) after feeding cassava. Between two cassava fed groups the reduction was more pronounced in the group fed fresh cassava than its cooked counterpart. Supplementation of iodine in both the cassava fed groups had increased the serum T4 and T3 level to that of only cassava fed groups but could not bring back to the control level. Discussion Cassava of Indian origin contains cyanogenic glucosides as well as glucosinolates and thiocyanate at various concentrations. The concentration of cyanogenic glucosides was reduced markedly after cooking while glucosinolates concentration was slightly lower and thiocyanate concentration was found almost same in cooked cassava in comparison to their uncooked counterpart. These goitrogenic substances act in conjunction with iodine deficiency in the development of endemic goitre and associated disorders (Lagasse et al, 1980) that emphasizes the need to investigate the role of goitrogens in influencing iodine utilization in the thyroid under varying state of iodine intake. In India, consumption of cyanogenic plant as evidenced by urinary thiocyanate level is considered as etiological factor for the persistence of residual goitre in post salt iodization phase (Marwaha et al, 2003). It was observed that the edible parts of the fresh and cooked plant have potent in vitro inhibitory activity on thyroid peroxidase enzyme. There are evidences to reverse the goitrogenic/ antithyroid potential of thiocyanate or thiocyanate like compounds present in cyanogenic plants by supplementing extra iodine (Rao and Lakshmy, 1995; Schone et al, 1990). Therefore in vitro anti-TPO potency of cassava had also determined in presence of extra iodide and found that extra iodide supplementation though had reduced the anti TPO activity to an extent but failed to cancel it. After ingestion, cyanogenic glucosides and glucosinolates present in foods are readily converted to active goitrogenic/ anti-thyroid agents, viz. thiocyanate and isothiocyanate etc. by glucosidases, sulphur transferase enzymes and myrosinase present in plant itself and animal tissues (Schone et al. 2001) and can give rise to thiocyanate in blood followed by its appearance in the urine. Consistent with this observation, thiocyanate levels in the urine of the cassava fed animals were elevated markedly. Thiocyanate or thiocyanate like compounds primarily inhibits iodide-concentrating mechanism of thyroid gland (Ermans, 1981). These compounds further stimulate the iodide efflux from thyroid gland (Ermans and Bourdoux, 1989;Maloof and Soodak 1959) resulting an increase in iodine excretion through urine (Schone et al. 2001). Present findings are consistent with those observations suggesting that thiocyanate precursors in cassava were metabolized to thiocyanate that had prevented iodide uptake by thyroid gland, stimulated thyroidal iodide efflux and was responsible for excreting more iodine through urine. The weight of the thyroid glands was increased significantly in both the fresh and cooked cassava fed groups mediated through the goitrogenic constituents and the weight gain was more in fresh cassava fed group than that of cooked one.Supplementation of extra iodine had reduced the gain in thyroid weight in both the cassava fed groups to an extent but it failed to normalize the effect. Thiocyanate has the same molecular size to that of iodide and thus inhibits the incorporation of iodide into thyroglobulin by competing with iodide at the thyroid peroxidase level and also inhibits the iodide oxidation (I- leads to I2) (Maloof and Soodak, 1959; Virion et al. 1980). Glucosinolates derivatives also interfere in the thyroid gland with organification of iodine and the formation of active thyroid hormone by interfering TPO activity, and their action usually may not be antagonised by iodine (Maloof and Soodak, 1959; Schone et al. 1990). Adequate iodine supplementation though had reduced the thyroid weight, but failed to bring back it in a normal morphological state and this observation further supports that goitrogenic potentiality of cassava may not be reversed even after the supplementation of excess iodine. Diet deprived of iodide but rich in thiocyanate, glucosinolate lowers the circulating level of thyroxine (T4) and triiodothyronine (T3) in rats (Papas et al. 1979; Lakshmy et al. 1995). Intake of cassava decreased the serum total T4 and T3 level significantly. Reduced TPO activity may be responsible for decreasing thyroid hormone levels because it regulates the synthesis of thyroid hormones (Taurog, 1970; Gaitan, 1990; Van Etten, 1969; Stoewsand 1995;Virion et al. 1980). Inhibition of iodide uptake and thyroidal iodide efflux due to excess thiocyanate (Ermans and Bourdoux, 1989;Maloof and Soodak 1959) might be other reasons for the decreased synthesis of thyroid hormones in cassava fed rats. Supplementation of extra iodide through diet have elevated thyroid hormone levels slightly but failed to bring back circulating normal thyroid hormone synthesis because the inhibition in TPO activity under cyanogenic plant diet not withdrawn even after extra iodine. The overall observations showed that chronic consumption of both fresh and cooked cassava had developed a relative state of biochemical as well as morphological hypothyroidism even in presence of adequate iodine in circulation. The etiological factors underlying this phenomenon are the dietary goitrogens present in cassava and their derivatives. Extra iodide supplementation along with cassava failed to prevent the TPO inhibition. References
© Copyright 2006 - African Journal of Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc06041t3.jpg] [tc06041t1.jpg] [tc06041t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}