 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
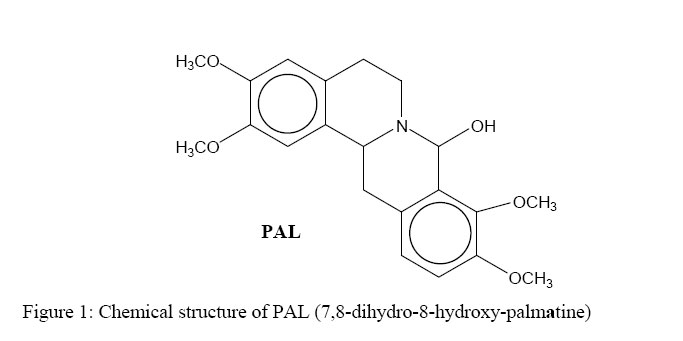
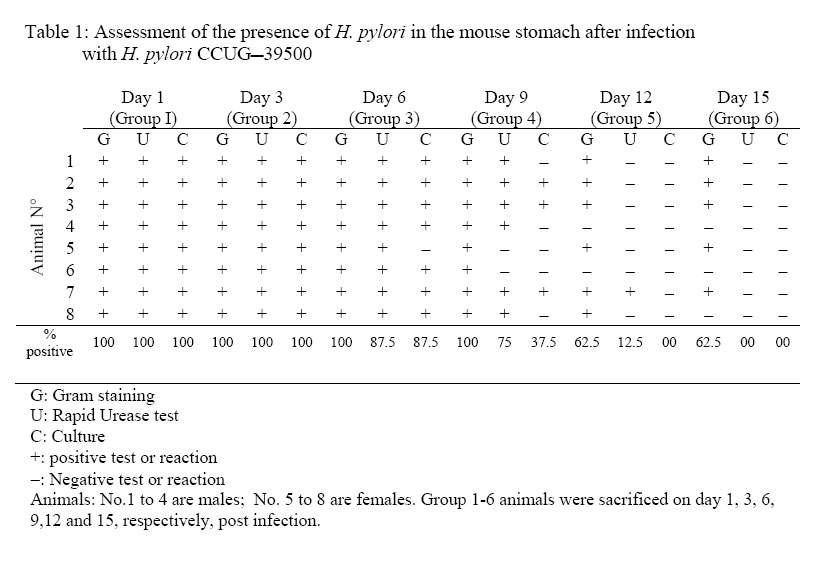
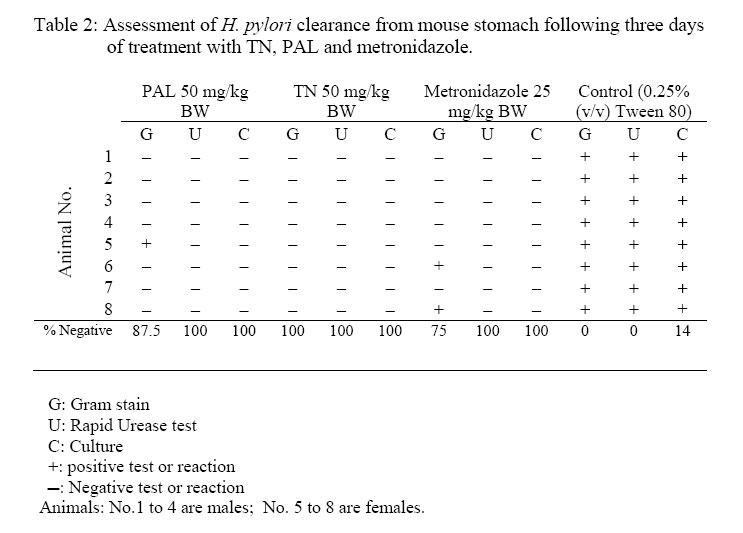
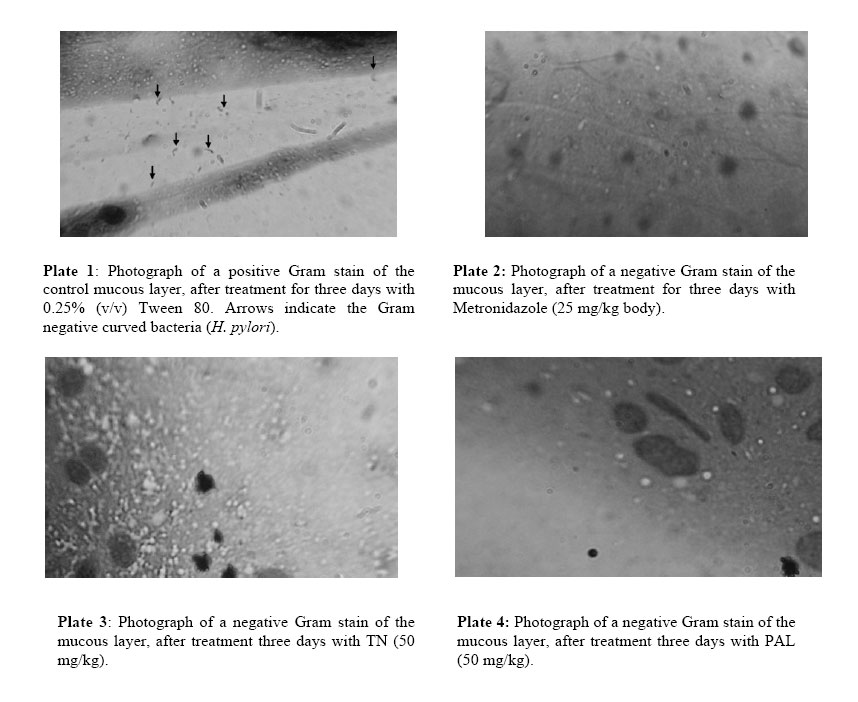
African Journal of Traditional, Complementary and Alternative Medicines, Vol. 3, No. 4, 2006, pp. 102-114 Research Paper RAPID IN VIVO SCREENING METHOD FOR THE EVALUATION OF NEW ANTI HELICOBACTER MEDICINAL PREPARATIONSMaurice Boda*, Paul V. Tan**, Barthelemy Nyasse$. ** Department of Animal Biology & Physiology, Faculty of Science, Box 812, University of Yaounde I, Cameroon, *Department of Biochemistry, Faculty of Science, Box 812, University of Yaounde I, Cameroon, $Department of Organic Chemistry, Faculty of Science, Box 812, University of Yaounde I, Cameroon E-mail: **pvernyuy@yahoo.com, Tel: 237 775 65 84. Code Number: tc06057 AbstractThe objective of this study was to devise a rapid mouse model for the in vivo screening of new anti Helicobacter pylori products. Six to eight week-old mice pre-treated (7 days) with Amoxicillin/Metronidazole (25 mg/kg) to eliminate all Helicobacter-like organisms were inoculated (4x per week) with 200 µl of a bacterial culture (108 CFU/ml of H. pylori CCUG-39500). Colonisation of mouse stomach was assessed from day 1 to 15 post inoculation by the Gram stain, rapid urease test and antral mucus culture. These findings were used as a mouse model of Helicobacter pylori infection to assess the in vivo anti Helicobacter activities of two plant-derived molecules with proven cytoprotective, ulcer healing and in vitro anti Helicobacter properties (TN, a tabersonine chloride alkaloid from the fruits of Voacanga Africana and PAL, a protoberberine alkaloid (7,8-dihydro-8-hydroxy-palmatine) from the bark of Enantia chlorantha). For up to 6 days post inoculation, the test organisms were recoverable by culture from 87.5% of the antral mucus samples while 100% of Gram stains were positive. The infection tended to clear naturally from day 9. Fifteen days post infection, only Gram stains were still positive (37.5%). Three-day oral administration of TN, PAL (50 mg/kg) and Metronidazole (25 mg/kg) completely eliminated the bacterial strain from the gastric antral mucus compared with the controls. This 4-week model is proposed as a rapid screening tool for in vivo evaluation of new anti H. pylori products in order to select candidates for detailed investigations. Keywords: Helicobacter pylori, in vivo, mouse model. Introduction Helicobacter pylori (H. pylori), has probably been part of the human gastric biota since time immemorial (Blaser, 1997), but its implication in chronic gastritis, peptic ulcer disease, gastric cancer as well as mucosa-associated lymphoid tissue (MALT) lymphoma has made it a major public health concern (DeCross and Marshall, 1993; Graham, 1997; Gibson et al., 1999) since these diseases are major causes of death worldwide (Radcliff et al., 1996). Genetic diversity between clinical isolates from different populations is one of the most distinctive characteristics of this pathogen (Niyaz and Sechi, 2005). These findings have considerably revolutionized the management of peptic ulcer disease and gastroduodenal malignancies. H. pylori eradication is now a major step in the therapeutic management of these diseases (Walsh, 1995; Beales, 2001), and effective treatment of H. pylori-induced peptic ulcer disease requires the use of at least two antimicrobial agents (selected among Amoxicillin, Metronidazole, Clarithromycin and Tetracycline) combined with antacids and/or antisecretory agents (Hentschel et al., 1993; Dunn et al., 1997; Beales, 2001; Kanamaru et al., 2001).The Emergence of H. pylori strains resistant to the recommended antibiotics, the associated side effects and low patient compliance, are cited as some of the major causes of treatment failure (Skirrow, 1992; Fauchère, 1999; Kato et al., 2000; Okamoto, 2002). Developing new anti-Helicobacter compounds is therefore of great importance. The literature is rich with information on the in vitro assessment of anti-Helicobacter properties of medicinal plant products (Nataliya et al. 2000; Yee and Koo, 2000; Kanamaru et al., 2001; Marone et al., 2001; Mahady et al., 2003a; Mahady et al., 2003b). However, the therapeutic usefulness of products active in vitro must be confirmed by in vivo tests. Although no animal model of H. pylori infection exactly mimics the human disease, experimental animal models using primates, domestic cats and dogs, gnotobiotic piglets, ferrets, Mongolian gerbils and mice have provided important insights into the in vivo evaluation of new antimicrobials. This has increased the understanding of host-pathogen relationships and virulence factors of the various strains of H. pylori (Danon et al. 1995; Enno et al., 1995; Fox et al., 1995; Sakigami et al., 1996; Lee et al., 1997; Lee, 1998; Krakowka et al., 1998; Kanamaru et al., 2001; Sgouras et al., 2004). The use of large animals is cumbersome and time-consuming but also very expensive and therefore available only to few investigators. Mouse-based H. pylori infection models are very useful tools, and for studies to be comparable between laboratories, standard criteria (the 1995 Lausanne criteria) were agreed upon. The Sydney strain of H. pylori (SS1) has been used successfully for the study of H. pylori pathogenesis, vaccine development and for the screening of anti H. pylori products (Lee et al., 1997; Lee, 1998; Jenks et al., 1999; Sizemore et al., 2002; Loughlin et al., 2003; Sgouras et al., 2004). Short stomach transit time and poor acid stability render many compounds endowed with in vitro anti-Helicobacter properties ineffective in vivo (Haas et al., 1990; Skirrow, 1992, Krakowka et al., 1998). Although, according to the Lausanne criteria, permanent colonisation leading to pathology is a pre-requisite for animal models designed to assess host-pathogen relationships and microbial eradication in well-established H. pylori-induced disease conditions, this may not necessarily be the case when the experimental goal is to screen large numbers of new products for in vivo anti Helicobacter activity. In many developing countries, the lack of experimental facilities usually hamper the effort of scientists working in this domain. For these reasons, we have designed a short-term (two-week) in vivo mouse model that can be used for the screening of new anti H. pylori products prior to the selection of candidates for more detailed expensive experimentation. This study describes a simplified in vivo method for assessing the anti-Helicobacter activity of compounds using the easily-cultured CCUG-39500 H. pylori strain in mice. The paper describes the preparation of H. pylori-free mice, the preparation and maintenance of H. pylori bacterial culture, confirmation of effective infection of mice with the H. pylori strain and the subsequent eradication of the microbes using two test compounds and a standard control drug. Materials and methodsPlant materials The alkaloid (7,8-dihydro-8-hydroxy-palmatine) prepared from Enantia chlorantha Oliver, Annonaceae (voucher specimen N° 25918/SRFCAM) was used. (Figure 1). The bark was collected from Edea in the Littoral Province of Cameroon. The extraction, isolation and purification of the active compound were done as previously reported (Tan et al., 2000). A more detailed report is presented by Wafo et al. (1999). The compound was code named PAL. The purified compound prepared from the fruits of Voacanga africana Stapf (Apocynaceae) is an alkaloid (TN) with Cameroon National Herbarium voucher specimen N° HNC/ 1949: P. Nana. The fruits were harvested in Yaoundé, sun dried and ground to a powder. Detailed preparation procedure for TN is reported by Tan and Nyasse (2000). Preliminary chemical and physical data suggested that the compound may correspond to tabersonine hydrochloride (m.p.192-195; [K]D -307; elemental analysis: C20H26O2N2.HCl) and its detailed structural elucidation awaits publication. Animals Six to eight-week old inbred healthy Swiss mice weighing between 17 and 23 g were used for the study. To avoid coprophagy, they were housed in raised wire mesh bottom cages. The animals were maintained on a 12 h light/dark cycle with water ad libitum, and fed with laboratory baked food made of maize (50%), soybeans (25%) and wheat flour (25%), supplemented with table salt, palm oil, fish and bone powder. Prior authorization for the use of laboratory animals was obtained from the Cameroon National Ethics Committee (Reg. No. FWA-IRB00001954). Culture media, and culture media supplements Columbia agar (CM0331), Brain Heart Infusion (CM0225), Lacked Horse Blood (SR0048), Horse Serum (SR0035), Vitox supplement (SR0090), Helicobacter pylori DENT supplement (SR0147) as well as the CampyGen gas pack (CN0024A) were all purchased from Oxoid, Basingstoke, England. The microbial strain Lyophilised Helicobacter pylori CCUG 39500 was obtained from the Culture Collection University of Göteborg (CCUG), Sweden. The strain was revived using Brain Heart Infusion supplemented with 10% horse serum (BHI-serum) and identified using the rapid urease and the catalase/oxydase tests (Fauchere, 1999; Dunn et al, 1997). It was maintained under microaerophilic conditions at 37°C on Columbia agar supplemented with 5% (v/v) lacked horse blood and 1% (v/v) Vitox, and subcultured every 96 hours to a fresh medium. The microaerophilic conditions were generated using CampyGen in a 2.5 L air-tight anaerobic jar (Oxoid). The isolation of H. pylori from mouse stomach biopsies was done on Columbia-DENT agar supplemented with 5% (v/v) lacked horse blood and 1% (v/v) Vitox (CD-Vitox). The infective inoculum was prepared by suspending 72-h colonies in 2 ml of sterile normal saline to make a turbidity standard of McFairland N° 1. The cell suspension of this turbidity standard was evaluated by standard plate count and was found to vary from 1.60 x108 to 3.05 x 108 CFU /ml (Cheesbrough, 1985). Antibiotics Amoxicillin (Amoxyl 500 mg/5 ml) and Metronidazole (Metronidazole USP RTU®;500 mg/100 ml) purchased from a local pharmacy were, respectively, from GlaxoSmithKline Australia Pty Ltd. and Baxter Healthcare Corporation, Deerfield. Stock solutions were prepared The concentrations used in mice were prepared using sterile normal saline. Eradication of Helicobacter-like organisms. In order to ensure that the mice used were free from other Helicobacter-like organisms (HLOs), they were pre-treated with a mixture of Amoxicillin (25 mg/kg) and Metronidazole (25 mg/kg) administered per os once daily for seven consecutive days in a final volume of 200 µl. A preliminary experiment was conducted on 20 animals grouped as follows: a test group (5 females and 5 males) treated for seven days with the HLO eradication regiment and a control group (5 females and 5 males) treated for 7 days with 200 µl of sterile normal saline (NaCl 0.9 %). The animals were fasted on the 8th day after which they were sacrificed. The stomachs were removed and the antral mucus layer was scraped. Part was used to prepare Gram stain slides and part was inoculated in urea broth while the remainder was homogenised in a test tube containing 500 µl of BHI. The homogenate was inoculated by flooding on CA-Vitox -DENT plate. The plate was incubated under microaerophilic conditions for five days before discarding. The urea broth was incubated in a candle jar for 3 hours after which the tubes were observed for any colour change. The Gram stain, urease test and culture technique did not reveal the presence of HLOs in both the test and control groups. Thus, the antibiotic pre-treatment satisfactorily certified the animals as helicobacter-free. Infection of mice with H. pylori CCUG-39500 and assessment of colonisation Forty-eight mice Helicobacter-free mice (24 males and 24 females) were allowed to rest for a further 7 days in order to clear the system of circulating antibiotics. Following a 12-h fast, the mice were inoculated per os with 200 µl of bacterial culture containing 108 CFU/ml of H. pylori CCUG-39500. The inoculation was done four times a week, with a 24-h interval between inoculations. Each inoculation was preceded by a 12-h fast. In order to confirm effective infection, six groups of 8 mice each (4 males and 4 females) were sacrificed respectively on days 1, 3, 6, 9, 12, and 15 following the last inoculation. H. pylori colonisation was assessed by Gram staining, rapid urease activity, and culturing of samples of the mucus layer of the antrum as described above. Positive cultures were further confirmed with the urease, catalase and oxydase tests. Assessment of H. pylori eradication. Mice (8 per group) infected as described above, were allowed to rest for a day after the last inoculation. They were then given the test products orally, once daily for three consecutive days. The plant preparations were administered in a final volume of 500 µl of 0.25% Tween 80 at the dose of 50 mg/kg. A positive control group was treated with Metronidazole (25 mg/kg) and a negative control group was given 500 µl of a sterile solution of 0.25% Tween 80. Following the last day of antibiotic treatment, the mice were fasted for 12 h after which they were sacrificed. The stomachs were excised and H. pylori clearance was assessed by Gram staining, rapid urease test and culture as described above. ResultsAssessment of H pylori CCUG-39500 infection of the mice In order to make sure that the animals used for the experiment were Helicobacter-free, all were pre-treated with an eradication regimen (Amoxicillin and Metronidazole) before infection with the experimental strain of H. pylori. The assessment of evidence of colonisation with H. pylori CCUG-39500 was simultaneously done by Gram staining, and the rapid urease test and culture for isolation of viable H. pylori. A slight change in the colour of the urease test tube within 3 hours was considered as positive. Some of the tubes showed a clear pink colour after 1 to 2 h but in some tubes a pale pink colour could be observed after three hours. There was therefore variation in the speed and the intensity of the urease reaction. From the 1st to the 6th day, evidence of colonisation was demonstrated by the Gram stain, rapid urease test and the culture technique (Table 1). The infection began to clear from the stomachs of 5 out of the 8 animals 9 days after inoculation with the bacterial culture. This was shown by negative cultures and urease tests. However, the Gram staining technique still revealed the presence of curved Gram negative bacteria in the smears prepared from the stomachs of all the group 4 animals sacrificed on day 9. Although the Gram stain continued to reveal the presence of infection in 5 to 6 animals (62.5 – 75%) up to day 15, the urease test and culture plates did not reveal signs of infection as of day 12 post inoculation. No apparent differences were noticed between male and female mice in infectivity with the microbe or onset of clearance from the stomachs. In vivo eradication of H. pylori CCUG-39500 in mice This assay was done in order to assess the ability of compounds with proven in vitro H. pylori activities to rid the stomach antrum of H. pylori following in vivo treatment. All the animals in the negative control group were successfully infected with H. pylori throughout the test period and the treatment with 0.25% (v/v). Tween 80, 500µl daily did not eradicate the pathogen (Table 2; Plate 1). In contrast, the cultures of the gastric tissue samples from the infected mice treated with PAL and TN as well as Metronidazole did not yield any growth after three days of treatment (Table 2; Plates 2-4). In the group treated with TN, the Gram stain, the rapid urease test, as well as the tissue cultures were negative for all the 8 animals. Positive Gram stains were obtained in one case for PAL and in two cases for Metronidazole. DiscussionStudies on the ecology of Helicobacter have proposed that various animal species harbour specific Helicobacter species. Thus, H. pylori, H. felis, H. mustelae, and H. heilmannii are species adapted to man, cats/dogs, ferrets and primates, respectively. (Stephen Danon et al., 1995). This notwithstanding, primates have been known to become infected with H. pylori, mouse models of H. felis which mimic the human pathology have been developed, a colony of SPF cats has been infected with a H. pylori isolate, and H. heilmannii is also known to be a natural feline Helicobacter ( Lee, 1998; Enno et al., 1995; Lee et al., 1990). Although early studies showed that mice could not be infected by clinical isolates of H. pylori, Lee et al. (1997) quickly developed a mouse model using a human clinical isolate, the “Sydney strain” of H. pylori. This is a cagA, vacA positive strain which has been widely used satisfactorily in Helicobacter-related peptic ulcer research. We have recently carried out an in vitro screening study of the antimicrobial activity against Helicobacter pylori (CCUG 39500) and Campylobacter jejuni-coli using 69 crude extracts obtained from Cameroonian medicinal plants (Tan et al., unpublished). The in vitro antimicrobial actions of the pure compounds used in the present study have also been demonstrated (Tan et al., unpublished). Unfortunately, these results can not be relied on as evidence of in vivo activity. Because of the high cost of equipment and instrumentation necessary to work in in vivo conditions that conform strictly to the Lausanne criteria of H. pylori experimentation, only few laboratories are privileged to work in this domain. For example, 1 – 2 months are needed after inoculation before colonisation with the Sydney strain of H. pylori is assessed, and it takes up to 8 months before a histologically-detectable chronic active gastritis develops in mice (Lee et al., 1997). For this reason, it would be of advantage during massive screening exercises to use a low-cost short-term model that can demonstrate the antimicrobial potencies of new products in vivo. A select number of highly active products can then be used as candidates for detailed work involving pathology in collaboration with advanced laboratories. The H. pylori strain used (the CCUG 39500 strain) is a human species that corresponds to the ATCC 43504 and NCTC 11637 strains (CCUG Web site a & b) which are routinely used for in vitro work largely due to their ease of cultivation (Best et al., 2003; Hassan et al., 1999; Svenson et al., 2002; Takeshi et al., 2002). Following CCUG instructions, the microbe was successfully revived and maintained in culture with regular sub culturing every 72 hours. All standard microbiological techniques for culture and assessment of colonisation were observed. However, the results obtained showed that the microbe was able to colonise the stomach mucous layer for up to 15 days maximum in contrast to the long-term colonisation usually obtained with the SS1 strain. Thus, the CCUG strain, although a human pathogen, is unable to withstand the mouse defence mechanism beyond two weeks and therefore cannot survive long enough in the mouse to induce pathological gastritis. However, the length of time the microbe survived in the mouse offers a window of opportunity to test the antimicrobial effect of anti-Helicobacter products under in vivo conditions. Thus, the decision to dose the animals with the antimicrobial products from days 2 to 4 post inoculation was based on the certainty of effective presence of the pathogen in the mucous layer within this period as shown by the Gram stain, urease and culture tests (Table 1). The SS1 strain can be left for up to one week after inoculation after which a two-week antimicrobial therapy is conducted (Lee et al., 1997). Thus, H. pylori CCUG-39500 is a human gastric mucosa-adapted strain of H. pylori, that is not adapted to mice for use in the study of the mechanism of chronic gastritis and gastric malignancy associated with H. pylori infection and to understand the pathogenesis of the infection. It may be necessary therefore to conduct pilot experiments order to ascertain the exact window of the short infection period that can be exploited for antimicrobial assessment using the CCUG-39500 strain. Since complete clearance of a standard H. pylori strain from the stomach was obtained during the treatment period, further quantitative analysis was not employed in order to detect its presence. However, it is suggested that in using the present model, quantitative evaluation, especially for new products showing partial clearance, should be carried out. Important insights into bacterial factors governing the colonisation as well as the host factors associated with the development of pathology have been provided by several animal models (Danon et al. 1995, Enno et al., 1995; Fox et al., 1995; Sakigami et al., 1996; Lee et al., 1997; Krakowka et al., 1998; Lee, 1998). For the in vivo assessment of anti Helicobacter compounds, mice and Mongolian gerbils are most often used (Kanamaru et al., 2001; Sgouras et al., 2004). However, there is no animal model of H. pylori infection that exactly mimics the human disease (Lee, 1998). The in vivo antimicrobial tests carried out using this short-term mouse model confirm the activity of metronidazole as an antibiotic of choice in H. pylori eradication therapy. The results show that in addition to the in vitro activity of PAL and TN, these new compounds also have in vivo antibacterial effects. The gastric anti-secretory activity of TN similar to that of H2 receptor blockers of histamine has been shown, and the ulcer healing properties of these compounds are associated with an enhancement of gastric mucus production (Tan et al., 2000; Tan and Nyasse, 2000). In combination with ranitidine, PAL and TN have also exhibited potentiating and synergistic antisecretory effects, respectively (Tan et al., 2002). The combined ulcer healing and antimicrobial potencies of these compounds enlist them as possible future candidates for the formulation of monotherapy antiulcer regimens with improved patient compliance. Positive results obtained from in vitro screening of extracts for anti Helicobacter activity do not constitute absolute proof of eventual in vivo efficacy. Thus, in spite of the obvious limitation of the present model which consist in the inability of the CCUG 39500 strain to colonize the mouse gastric mucous layer beyond two weeks, it nevertheless offers the opportunity for the short-term rapid screening of new plant products under in vivo conditions. Possible candidates for more detailed investigations can thus be selected at low cost. AcknowledgementsThis research was supported by the International Foundation for Science, Stockholm, Sweden, and the UnitedNationsUniversity (UNU), Tokyo, Japan, through grant F/2882-2 awarded to Dr Paul V. Tan and IFS/OPCN grant F2626-3 awarded to Prof. B. Nyasse. References
© Copyright 2006 - African Journal of Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc06057t2.jpg] [tc06057f1.jpg] [tc06057f2.jpg] [tc06057t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}