 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
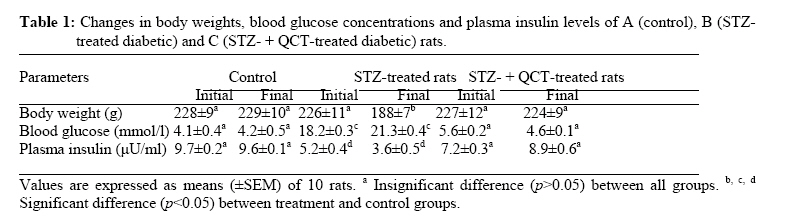
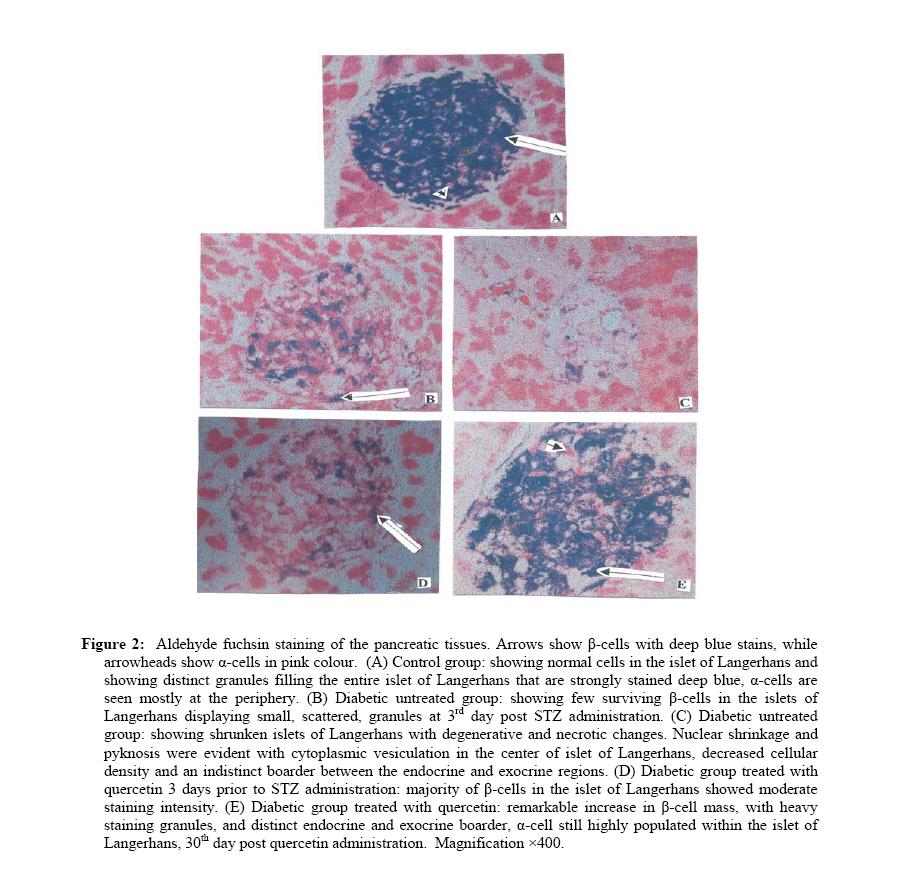
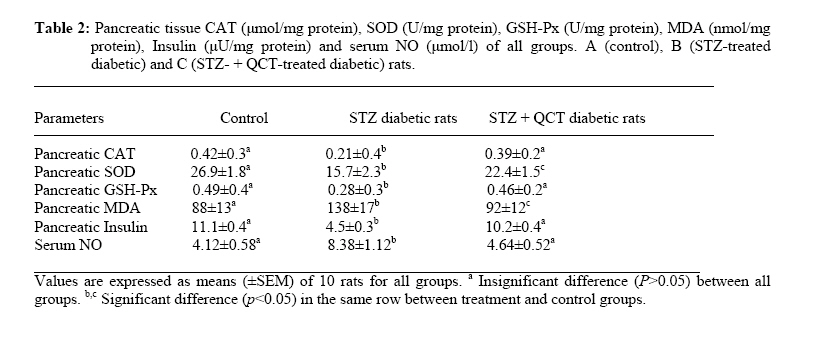
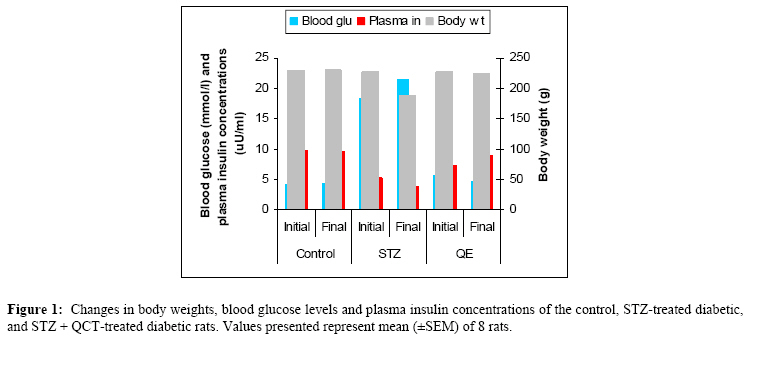
African Journal of Traditional, Complimentary and Alternative Medicines,, Vol.4, No. 1, 2007, pg. 64-74 RESEARCH PAPER PROTECTIVE EFFECT OF QUERCETIN ON THE MORPHOLOGY OF PANCREATIC β-CELLS OF STREPTOZOTOCIN-TREATED DIABETIC RATS Stephen O. Adewole1*, Ezekiel A. Caxton-Martins1 and John A. O. Ojewole2 1*,Department of Anatomy and Cell Biology, Faculty of Basic Medical Sciences, College of Health Sciences, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria; Code Number: tc07010 Abstract This study was undertaken to investigate the protective effects of quercetin (QCT) on the morphology of pancreatic β-cells against diabetes mellitus and oxidative stress experimentally-induced by streptozotocin (STZ) treatment in Wistar rats. Fifty male and female Wistar rats (200–250 g) were randomly divided into three experimental groups (i. e., control, STZ-treated, and STZ + Quercetin-treated groups). Diabetes was induced in the diabetic groups (B and C) of animals, by a single intraperitoneal injection of STZ (75 mg/kg), while each of the rats in the ‘control’ group received equal volume of citrate buffer (pH 6.3) solution intraperitoneally. In group C rats, quercetin (QCT, 25 mg/kg/day i. p.) was injected daily for 3 days prior to STZ treatment, and QCT administration continued until the end of the study period (30 days). Diabetes mellitus was confirmed by using Bayer’s Glucometer Elite® and compatible blood glucose test strips. The rats were sacrificed serially until the end of the study period (after 30 days). The pancreases of the sacrificed rats were excised and randomly processed for histological staining and biochemical assays for antioxidant enzymes [such as glutathione peroxidase (GSHPx), superoxide dismutase (SOD), catalase (CAT), malondialdehyde (MDA) and serum nitric oxide (NO)]. In the diabetic state, pancreatic β-cells of STZ-treated group B rats histologically demonstrated an early chromatin aggregation, cytoplasmic vesiculation in the central β-cells, nuclear shrinkage, and lysis of β-cells with distortion of granules. The morphology of QCT-treated rats’ pancreases showed viable cellularity with distinct β-cell mass. STZ treatment significantly decreased (p<0.05) GSHPx, SOD, CAT and pancreatic insulin content. However, STZ treatment increased blood glucose concentrations, MDA and serum NO. The QCT-treated group of animals showed a significant decrease (p<0.05) in elevated blood glucose, MDA and NO. Furthermore, QCT treatment significantly increased (p<0.05) antioxidant enzymes’ activities, as well as pancreatic insulin contents. Quercetin (QCT) treatment protected and preserved pancreatic β-cell architecture and integrity. In conclusion, the findings of the present experimental animal study indicate that QCT treatment has beneficial effects on pancreatic tissues subjected to STZ-induced oxidative stress by directly quenching lipid peroxides and indirectly enhancing production of endogenous antioxidants.Key words: Quercetin, Streptozotocin, Antioxidant enzymes, Pancreatic β-cell. Introduction Flavonoids occur commonly, and are widespread, in the plant kingdom. They function as plant pigments, and are responsible for the colours in flowers and fruits. Flavonoids are plant polyphenolics found as pigments in leaves, barks, seeds, fruits and flowers. They are often closely associated with vitamin C, to which they offer synergistic effects (Stephen-Cole, 1998). The pharmacological effects of many traditional herbal drugs (phytomedicines) may be ascribed to their flavonoid constituents (Hertog and Hollman, 1996), which inhibit certain enzymes and possess antioxidant activities. Quercetin (3,3΄,4΄,5-7-pentahydroxyflavone), is a flavonol chemically related to kaemferol. As a member of the flavonoid family, quercetin is widely distributed in plants, and is probably the most abundant of the flavonoid molecules in the plant kingdom. Sources of quercetin include brassica green vegetables, berries, onions, parsley, apple, legumes, green tea, citrus fruits, red grape wines, and so on. Quercetin prevents oxidantive injury and cell death (Larocca et al; 1995) by several mechanisms, including scavenging oxygen radicals (Saija et al; 1995; Miller, 1996; Cox et al., 2000), inhibiting xanthine oxidase (Chang et al; 1993), lipid peroxidation, and chelating metal ions (Chen et al; 1990). Quercetin possesses a catalogue of pharmacological actions, including cardio-protection, cataract prevention, anti-cancer activity, anti-ulcer effects, anti-inflammatory, anti-allergic, antiviral and antibacterial activities, and so forth (Bronner and Landrry, 1985; Stavric, 1994). Quercetin also inhibits oxidation of low density lipoprotein (LDL) cholesterol in vitro, probably by inhibiting LDL oxidation itself, by protecting vitamin E (α-tocopherol) in LDL from being oxidized, or by regenerating oxidized vitamin E (DeWhalley et al; 1990).Streptozotocin (STZ), an antibiotic produced by Streptomyces achromogenes, possesses pancreatic β-cell cytotoxic effect (Weiss, 1982). Streptozotocin has been widely used for inducing diabetes mellitus in a variety of animals. STZ causes degeneration and necrosis of pancreatic β-cells (Uchigata et al; 1982; Merzouk et al; 2000). Although the mechanism of the β-cell cytotoxic action of STZ is not fully understood, experimental evidence has demonstrated that some of its deleterious effects are attributable to induction of metabolic processes, which lead on to an increase in the generation of reactive oxygen species (ROS) (Chen et al; 1990). Apart from production of ROS, STZ also inhibits free radical scavenger-enzymes (Kröncke et al; 1995). The superoxide radical has been implicated in lipid peroxidation, DNA damage, and sulfhydryl oxidation (Tiedge et al; 1997; Matkovics et al; 1998). Nitric oxide synthase (NOS) is present in pancreatic β-cells, and may be involved in the release of insulin under normal physiological conditions (Moncada et al; 1991). However, recent findings suggest that induction of nitric oxide formation may play a role in the destruction of β-cells during the development of type 1 diabetes mellitus (Corbbet et al; 1993). Antioxidant defense mechanisms are important for the protection of cells and tissues against oxidative damage. The major endogenous antioxidant enzyme-systems in mammalian body include superoxide dismutase (SOD), catalase (CAT), selenium-dependent glutathione peroxidase (GSHPx-Se), glutathione peroxidase (GSHPx), and glutathione reductase (GSHR). The major non-enzymatic endogenous antioxidants include glutathione (GSH) and vitamin E (Nadler and Winter, 1996). Reduced antioxidant levels, as a result of increased free radical production in experimental diabetes, have been reported by Grankvist et al., (1981), and Kanter et al., (2003a). Diabetes mellitus (DM) is a group of heterogenous, hormonal and metabolic disorders characterized by hyperglycemia and glucosurea, resulting from defects in insulin secretion, insulin action, or both (WHO, 1999). The chronic hyperglycemia of diabetes is associated with long-term damage, dysfunction, and failure of various organs in the body, especially the eyes, kidneys, nerves, heart, pancreas itself, and blood vessels (WHO, 1985). Pancreatic insulin reserve is an important parameter of islet function, with tight coupling between insulin secretion and production being necessary for adequate functioning of pancreatic β-cells (Kaiser et al; 2003). Insulin store size is determined by the balance between the rates of insulin biosynthesis and secretion, as well as by the number and volume of the β-cells, i.e., the β-cell mass. In adult mammals, β-cell mass varies to adapt insulin secretion to long-term changes in insulin demand. This has been demonstrated repeatedly under physiological as well as pathological conditions of insulin resistance (Bernard-Kargar and Ktorza, 2001). Although mainly demonstrated in rodents, this situation also seems to be the case in obese humans (Kloppel et al; 1985; Butler et al; 2003). The deterioration of metabolic state in type 2 diabetes mellitus results mainly from progressive β-cell failure (UK Prospective Diabetes Study – UKPDS, 1998; Weyer et al; 1999). Therefore, it is critically important to determine whether or not functional β-cell mass is reduced in type 2 diabetics. In the present study, we have used type 1 diabetic animal model to evaluate the variations in pancreatic insulin reserve through β-cell mass, and influence of prompt normoglycaemia induction on pancreatic β-cells, since hyperglycemia is associated with rapid depletion of pancreatic insulin stores, temporal changes in β-cell proliferation, and in death that culminates in disturbed islet morphology (Gadot et al; 1995; Donath et al; 1999; Jorns et al; 2002). Type 1 or insulin-dependent diabetes mellitus (IDDM) results from autoimmune destruction of the pancreatic β-cells (Bach, 1994). Due to severe deficiency or relative lack of insulin, the key pancreatic hormone necessary for glucose homeostasis, patients with type 1 diabetes suffer from elevated blood glucose levels manifested as polyurea, polydypsia, glucosurea, ketoacidosis, and weight loss. Development of long-term diabetic complications, such as nephropathy, retinopathy, neuropathy and micro- and macro-vascular diseases is also very dangerous and lethal. Although oral or inhaled forms of insulin are now being used to achieve better glycemic control, these strategies need to be further improved (Dong et al; 2002). The current increasing prevalence of DM in the world is of great concern. Thus, we have thought it worthwhile to investigate the amelioration of experimentally-induced diabetes mellitus with quercetin. Therefore, the core objectives of the present study were: (a) to evaluate whether the pancreas is subjected to oxidative damage during diabetes, and (b) to examine the accompanying changes in antioxidant status in order to understand the role of antioxidants in the pathogenesis of diabetes mellitus. We have also probed whether quercetin protects pancreatic β-cells against STZ-induced injury. Materials and Methods The experimental protocol used in this study was approved by the Ethics Committee of the University of KwaZulu-Natal, Durban 4000, South Africa; and conforms with the “Guide to the Care and Use of Animals in Research and Teaching” [published by the Ethics Committee of the University of Durban-Westville, Durban 4000, South Africa].Animals Fifty healthy, male, Wistar rats weighing 200–250 g (averaging 12 weeks old) were used in this study. They were housed under standard laboratory conditions of light, temperature and humidity. The animals were given standard rat pellets and tap water ad libitum. The rats were randomly divided into 3 experimental groups: A (control), B (diabetic), and C (diabetic + quercetin). Control group animals were treated with citrate buffer solution intraperitoneally. Diabetes was induced in both groups B and C rats by a single intraperitoneal (i.p.) injection of STZ (75 mg/kg body weight, freshly dissolved in 0.1mol/l citrate buffer, pH 6.3), (Rossini et al; 1978). The experimental animals in both groups B and C became diabetic within 48 h after STZ administration. Quercetin (25 mg/kg/day, i.p.) was injected daily for 3 days prior to administration of STZ in group C rats, and QCT treatment continued until the end of study period (30 days).Histopathological procedures Pancreatic tissues were harvested from the sacrificed animals before, and, 5 and 30 days after, STZ and QCT treatments. Pancreatic tissues were fixed in Bouin’s fixative solution, and were sequentially embedded in paraffin blocks according to the standard procedure, sectioned at 6 µm thickness, and then stained for functional pancreatic β-cells with Gomori’s Aldehyde Fuchsin (GAF), (Mowry et al; 1980).Biochemical assays Blood glucose and plasma insulin analyses Two days after STZ treatment, development of diabetes mellitus was confirmed by measuring blood glucose from tail vein blood samples, using Bayer’s Glucometer Elite® and compatible blood glucose test strips (Henry, 1984). STZ-treated rats with blood glucose concentrations ≥18 mmol/l were considered to be diabetic, and used in this study. Blood glucose levels in control animals remained normal (4.1–4.5 mmol/litre) for the duration of the study. Plasma insulin was determined by a double-antibody radioimmunoassay (RIA) kit (Amersham Radiochemical Centre, Bucks, UK) (Kanter et al; 2003b). All serum insulin samples were assayed by RIA, using crystalline rat insulin as standard. Free and antibody-bound insulin was separated by precipitation with ethanol (Desbusquois and Aurbach, 1971). Preparation of pancreas homogenates The rats were sacrificed by cervical dislocation and pancreata were excised, rinsed in ice-cold physiological saline, and homogenized with Potter Elvehjem homogenizer. 10% homogenates were prepared in 6.7 mM phosphate buffer, pH 7.4 and centrifuged at 10,000 rpm for 10 min at 4oC, and the supernatant was used for antioxidant enzyme assays. For the determination of lipid peroxidation, pancreatic tissues were homogenized in 1.15% KCl solution to obtain a 10% (w/v) homogenate. Protein content of pancreas homogenates was determined by using bicinchoninic acid (BCA) protein assay reagent (Pierce Chemical Company, Rockford, IL).Catalase Activity (CAT) The activity of catalase (CAT) was measured using its perioxidatic function according to the method of Johansson and Borg (1988). 50 µL potassium phosphate buffer (250 mM, pH 7.0) was incubated with 50 µl methanol and 10 µL hydrogen peroxide (0.27%). The reaction was initiated by addition of 100 µL of enzyme sample with continuous shaking at room temperature (20oC). After 20 minutes, reaction was terminated by addition of 50 µL of 7.8 M potassium hydroxide. 100 µL of purpald (4-Amino-3-hydrazino-5-mercapto-1,2,4-triazole, 34.2 mM in 480 mM HCl) was immediately added, and the mixture was again incubated for 10 minutes at 20oC with continuous shaking. Potassium peroxidate (50 µL 65.2 mM) was added to obtain a coloured compound. The absorbance was read at 550 nm in a spectrophotometer. Results are expressed as micromoles of formaldehyde produced/mg protein.Superoxide Dismutase Activity (SOD) Superoxide dismutase (SOD) activity was determined by inhibition of cytochrome C reduction by the method of Flohe and Otting (1984). The reduction of cytochrome C was mediated by superoxide anions generated by xanthine/xanthine oxidase system, and monitored at 550 nm. One unit of SOD was defined as the amount of enzyme required to inhibit the rate of cytochrome C reduction by 50%. Results are expressed as units/mg protein.Glutathione Peroxidase Activity (GSH-Px) Glutathione peroxidase (GSH-Px) activity was measured by NADPH oxidation, using a coupled reaction system consisting of glutathione, glutathione reductase, and cumene hydroperoxide (Tappel, 1978). 100 µL of enzyme sample was incubated for five minutes with 1.55 ml stock solution (prepared in 50 mM Tris buffer, pH 7.6 with 0.1 mM EDTA) containing 0.25 mM GSH, 0.12 mM NADPH and 1 unit glutathione reductase. The reaction was initiated by adding 50 µL of cumene hydroperoxide (1 mg/ml), and the rate of disappearance of NADPH with time was determined by monitoring absorbance at 340 nm. One unit of enzyme activity is defined as the amount of enzyme that transforms 1 µmol of NADPH to NADP per minute. Results are expressed as units/mg protein.Lipid Peroxidation contents (LPO) The product of the reaction between malondialdehyde (MDA) and thiobarbituric acid reactive substances (TBARS) were measured by a modified method of Ohkawa et al., (1979). For each sample to be assayed, four tubes were set up containing 100, 150, 200 and 250 µL of tissue homogenate, 100 µL of 8.1% SDS, 750 µL of 20% acetic acid, and 750 µL of 0.8% aqueous solution of TBA. The volume was made up to 4 ml with distilled water, mixed thoroughly and heated at 95oC for 60 minutes. After cooling, 4 ml of n-butanol was added to each tube, the contents mixed thoroughly, and then centrifuged at 3000 rpm for 10 minutes. The absorption of the clear upper (n-butanol) layer was read at 532 nm. 1, 1, 3, 3 tetraethoxy propane (97%) was used as the external standard. Results are expressed as nmoles TBARS/mg tissue.Nitric Oxide (NO) Serum nitrite/nitrate levels were determined by converting the nitrate to nitrite, using enzyme nitrate reductase followed by addition of Griess reagent to colorimetrically quantify the nitrite concentration (Green et al; 1982). The serum was diluted 1:5 in PSB before a 25-µl aliquot was added to a mixture of 25 µl nitrate reductase (1.5 U/ml) and 25 µl of NADPH (0.134 mg/ml), both prepared in 40 mM Tris, pH 7.6. The samples were thereafter incubated at room temperature for 3 hours. Following this period, 100 µl of Griess reagent (1:1 mixture of 1% sulphanilamide in 5% phosphoric acid and 0.1% naphyly-ethylenediamine) was added and incubated for a further 10 min at room temperature; the absorbency of the samples was measured at 540 nm with a 650 nm reference. The concentration of nitrite/nitrate was determined from a standard curve of sodium nitrate and calculated as µM nitrite/nitrate.Pancreatic insulin contents The splenic regions of the pancreatic tissues from euthanized rats were weighed and homogenized, on various experimental days, in acid-ethanol solution (75% ethanol, 23.5% distilled water1.5% concentrated HCl). After overnight incubation at 40C, the suspensions were centrifuged, and the supernatants were collected and assayed for insulin contents, using a competitive ELISA kit (Kekow et al; 1988). Plates were coated with rabbit anti-guinea-pig Ig secondary Ab (Organon Teknoka, Durhan, NC), followed by incubation with a guinea-pig anti-human insulin Ab (Cortex Biochem, San Leandro, CA). Following two washing steps, various extract dilutions or insulin standards (Linco Research, St. Louis, MO) were mixed with constant concentration of HRP-conjugated rat insulin (Organon Teknika) for 4 h at room temperature, or at 40C overnight, before competitive capturing was allowed for 3 h. After washing five times, Sigma FAST OPD tablets (Sigma, St. Louis, MO) were used as substrate. Results were analyzed using ceres 900 C ELISA-reader. Statistical Analysis The data obtained were expressed as means (±SEM), and analyzed using repeated measures of variance. The differences between the means were analyzed statistically with one-way analysis of variance (ANOVA; 95% confidence interval). Values of p<0.05 were taken to imply statistical significance. Results Characteristics of diabetic state Forty-eight hours after STZ administration, all animals that had been treated with STZ displayed glucosurea, ketonurea, hyperglycemia, hypoinsulinemia and a moderate loss of body weight. Blood glucose concentrations and plasma insulin levels of the STZ-treated experimental animals are shown in Table 1. The baseline weight of the rats at the beginning of the study was similar in all groups. At the end of the study period (30 days), diabetic animals in group B presented with weight loss. The initial and final body weights were not significantly different in control and quercetin-treated groups. Histopathological findings The histology of pancreatic islet cells of control rats was normal. In diabetic rats without QCT treatment, the most consistent findings in the sections of pancreatic tissues were breakdown of micro-anatomical features such as extensive β-cell degranulation, decreased cellular density, and an indistinct boarder between the endocrine and exocrine regions. There were also diffused degenerative and necrotic changes, and shrunken in the islets of Langerhans (Figure. 2B).The nucleus of necrotic cells showed either pyknosis or marginal hyperchromasia, and the cytoplasm were filled with hydropic changes (Figure. 2C). The exocrine pancreatic acinar epithelium, ductal and connective tissues appeared normal. In diabetic rats treated with quercetin, identifiable pancreatic islets 48 h after STZ-treatment were of low cellularity (Figure. 2D). There was no evidence of inflammatory cell infiltration or fibrosis. At the end of the study period (30 days), quercetin proved to be protective to the pancreatic β-cells, and the islets of Langerhans were distinct with increase in granulation intensity. Biochemical findings There was a clear evidence that STZ-induced pancreatic injury was associated with free radical injury and oxidative stress. Oxidative stress was characterized by increased lipid peroxidation and/or altered non-enzymatic and enzymatic antioxidant systems (Zima et al; 2001). Effects of STZ and STZ + QCT treatments on pancreatic tissue CAT, SOD, GSH-Px, MDA, insulin and serum NO are presented in Table 2. The antioxidant levels and pancreatic insulin contents significantly decreased (p<0.05), while serum NO concentrations and pancreatic MDA significantly increased in the STZ-treated, diabetic rat group. Quercetin (QCT) treatment significantly (p<0.05) decreased the elevated NO and MDA, but also significantly increased (p<0.05) the reduced antioxidant enzyme activities. Discussion The findings of the present study indicate that quercetin exerts protective effects on STZ-induced oxidative stress and β-cell damage in diabetic rats. Free radical mechanisms and the possible sources of oxidative stress in the pathogenesis of diabetes and diabetic complications have been extensively studied in animal models and in diabetic patients (Baynes, 1991). Diabetic individuals and experimental animals exhibit high oxidative stress due to persistent and chronic hyperglycemia, thereby depleting the activity of oxidative defense system, and thus promoting de novo free radical generation (Hammers et al; 1991). Oxidative stress has recently been shown to be responsible, at least in part, for pancreatic β-cell dysfunction caused by glucose toxicity. Under hyperglycemia, production of various reducing sugars, such as glucose-6-phosphate and fructose, increases through glycolysis and polyol pathways. During this process, reactive oxygen species (ROS) are produced and cause tissue damage (Sakurai and Tsuchiya, 1988; Hunt et al; 1990; Matsuoka et al; 1998).Oxidative stress, implicated in the pathogenesis of a wide range of clinical disorders, refers to the cytological consequences of a mismatch between the production of free radicals and the ability of the cells to defend against them. Oxidative stress can thus occur when the generation of free radicals increases, or the capacity to scavenge free radicals and repair of oxidatively modified macro-molecules decreases, or both (Sies, 1997). This imbalance leads to accumulation of oxidatively modified molecules, predominantly end- products of superoxide (O2-) and hydroxyl (OH-) actions. Hydrogen peroxide (H2O2) and peroxynitrate (ONOO-), although not free radicals themselves, contribute to the cellular redox state. Collectively, these molecules, commonly referred to as reactive oxygen species (ROS), produce significant functional alterations in lipid, protein and DNA molecules (Jacob and Burri, 1996). Oxidative lipid damage, referred to as lipid peroxidation, produces a gradual loss of cell membrane fluidity, reduces membrane potential, and increases permeability to ions like Ca2+. Oxidative stress has been proposed to be involved in the pathophysiology of many chronic diseases, including atherosclerosis (Gutteridge and Halliwell, 1990) and diabetes (Richelle et al; 1999), and is known to accelerate ageing processes (De la Fuente and Victor, 2000). Antioxidants form an important part of a cell’s defense mechanism against free radical damage. Antioxidant enzymes, in particular, constitute a major part of this defense mechanism. Superoxide dismutase (SOD) detoxifies peroxide radicals, giving rise to hydrogen peroxide (H2O2), and is the only known enzyme that uses free radicals as a substrate. However, H2O2 is itself a potent free radical generator, and can generate toxic hydroxyl radicals by reacting with ferrous ions, which can induce lipid peroxidation of cell membranes; but cellular GSH-Px and CAT easily detoxify H2O2. It is important that an enhanced SOD activity be followed up to the activities of CAT and GSH-Px, in order to prevent accumulation of toxic H2O2 (Slater, 1984). In addition to these enzymes, glutathione transferases are a group of isoenzymes capable of detoxifying exogenous and endogenous substances in conjugation with glutathione, including organic hydroperoxides. Lipid peroxidation (MDA) is widely used as an indicator to reflect oxidative stress and cell membrane damage (Halliwell and Gutteridge, 1989). Free radicals, like superoxide anion and hydroxyl radical, exert their toxic effects by acting on DNA, membrane proteins and lipids. Cytotoxic drugs like streptozotocin are known to exert their cytotoxic effects by a free radical mediated mechanism (Dumontet et al; 2001). In our study, we induced experimental type 1 diabetes, and we have shown that quercetin is effective not only in insulin generation and sensitivity, but also in β-cell protection. Quercetin appears to have many beneficial effects on human health, including cardio-protection, anti-cancer activity, anti-ulcer effects, anti-allergy activity, cataract prevention, antiviral and anti-inflammatory effects (Miller, 1996). The main mechanism of action of flavonoids, as a rule, is antioxidant activity, and a number of quercetin’s effects appear to be due to its antioxidant activity. Quercetin scavenges oxygen radicals (Saija et al; 1995), and inhibits xanthine oxidase and lipid peroxidation in vitro (Chang et al; 1993; Chen et al; 1990). The anti-inflammatory activity of quercetin protects pancreatic β-cells from type 2 diabetes, by inhibiting inflammation-producing enzymes (cyclooxygenase, lipoxygenase) and subsequent inhibition of inflammatory mediators, including leukotrienes and prostaglandins (Della Logia et al; 1988; Kim et al; 1998). Inhibition of histamine release from mast cells and basophils (Fox et al; 1988; Bronner and Landry, 1985), also contributes to quercetin’s anti-inflammatory activity. Quercetin also blocks aldose reductase, an enzyme which catalyzes the conversion of glucose to sorbitol, which has been linked to nerve, eye and kidney damage in diabetes mellitus (Chaudry et al; 1983). In the present study, the effects of quercetin in rats with STZ-induced oxidative stress and β-cell damage were evaluated through histological staining and biochemical assays. Quercetin caused significant decrease in elevated blood glucose and an increase in plasma insulin concentrations in STZ-induced diabetic rats. Histologically, quercetin partly protected pancreatic β-cell integrity. It saved the tissue from the usual shrunken islets, degeneration, degranulation, hydropic and necrotic changes peculiar to STZ-induced pancreatic injuries. In this study, we also examined changes in free radical levels, and we found that STZ treatment caused significant increases in lipid peoxidation (MDA) and nitric oxide (NO) generation, and decreased CAT, SOD, GSH-Px activities as well as pancreatic insulin contents when compared with the control animals. Decreases in SOD, CAT and GSH-Px, and simultaneous increases in MDA and NO activities, reflect susceptibility of pancreas to STZ’s significant oxidative stress. Thus, this study has demonstrated that STZ alters the activity of antioxidant defenses in the pancreas. This observation is in consonance with the studies reported by Henriksen (2002), Gul et al., (2002), Goodyear and Kahn (1998), and Derouich and Boutayeb (2002). Based on our findings, we conclude that STZ administration is associated with oxidative stress in pancreatic tissues. Nevertheless, quercetin exhibited antioxidant activity, and is able to diminish and/or prevent, pancreatic oxidative damage produced by STZ. Flavonoids are frequently consumed in human diet, especially if the diet is rich in fruits and vegetables. An increased consumption of antioxidants in the diet of individuals is strongly recommended, so that when an individual is subjected to greater oxidative stress, s/he would have better antioxidant defense characteristics, thus counteracting the effects of any pro-oxidant. However, further studies are needed before antioxidants can be used safely as food additives and supplements. Acknowledgements The authors are grateful to Prof. C. Rogers for the gift of quercetin used in this study, and to Messrs Adeogun Oludele, Joseph Ibeh, and Izobo Stanley for their technical assistance in tissue processing.References
The following images related to this document are available:Photo images[tc07010t2.jpg] [tc07010f2.jpg] [tc07010t1.jpg] [tc07010f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}