 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
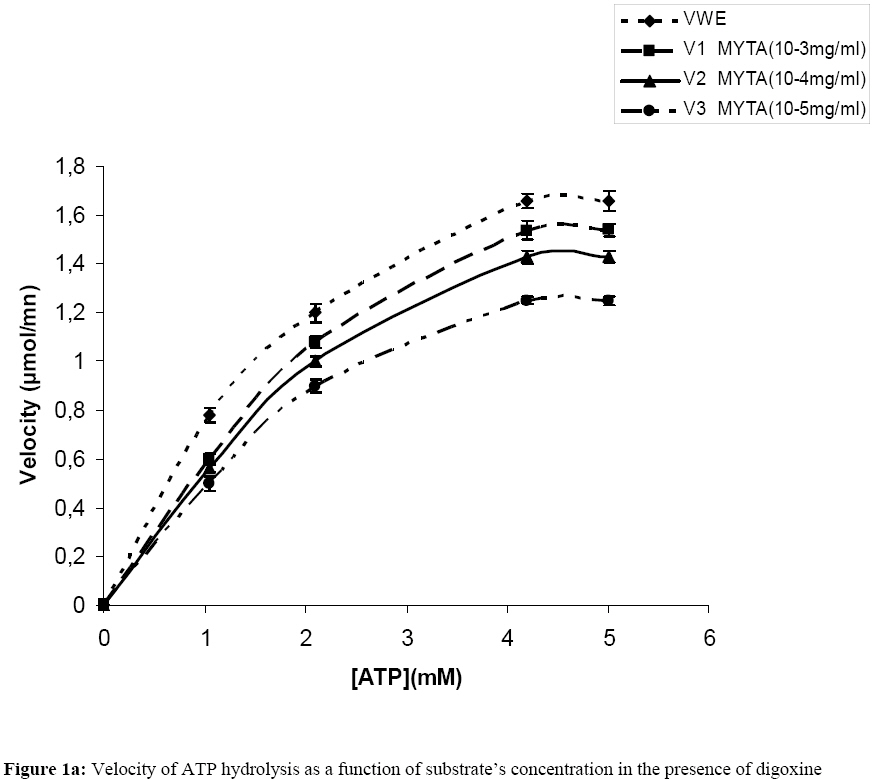
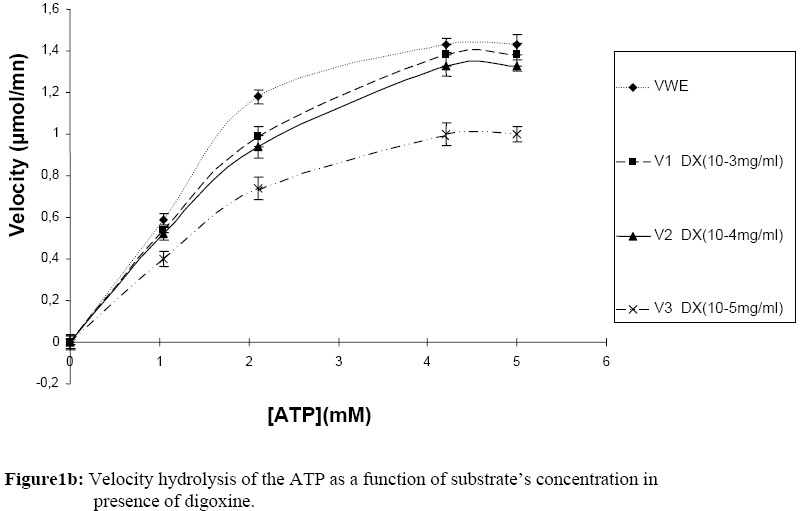
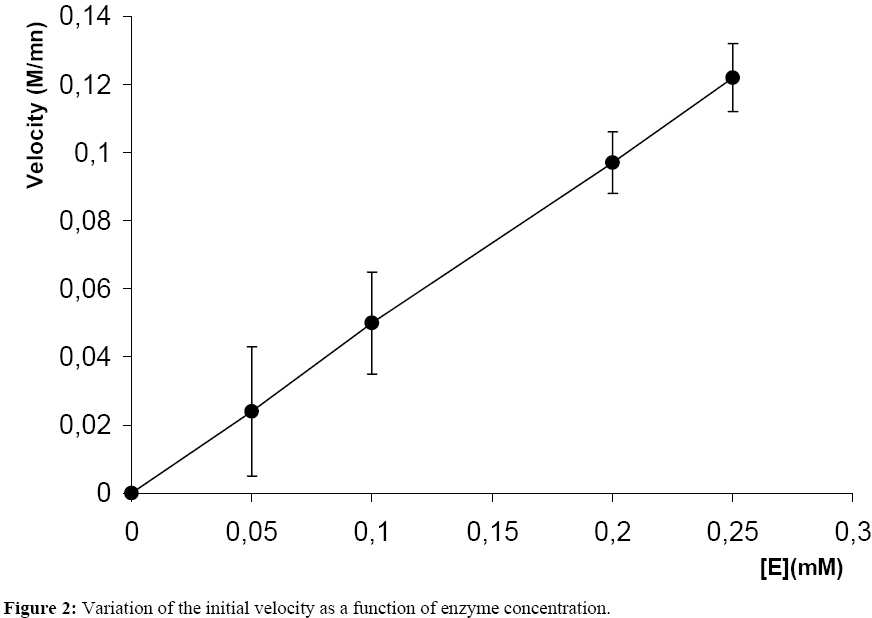
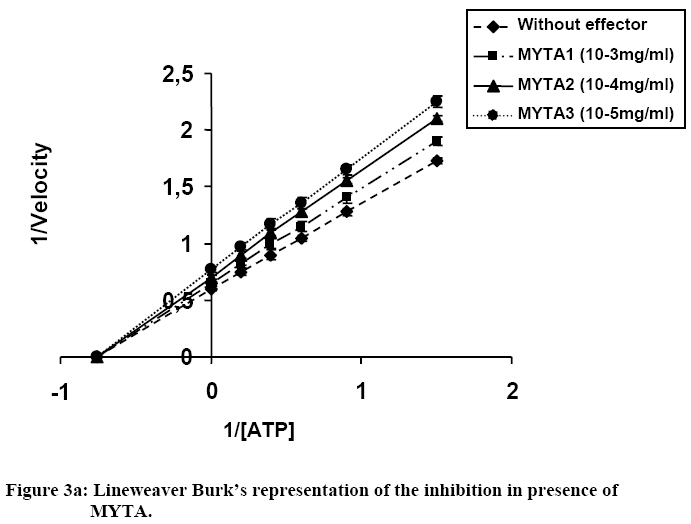
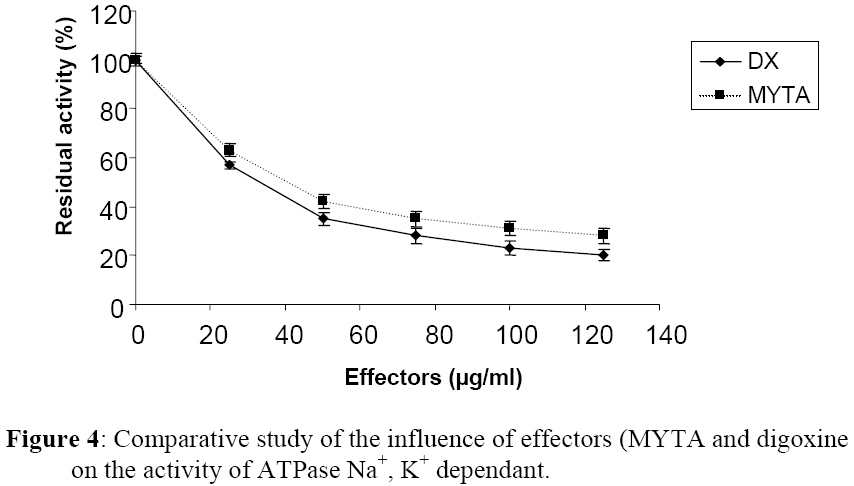
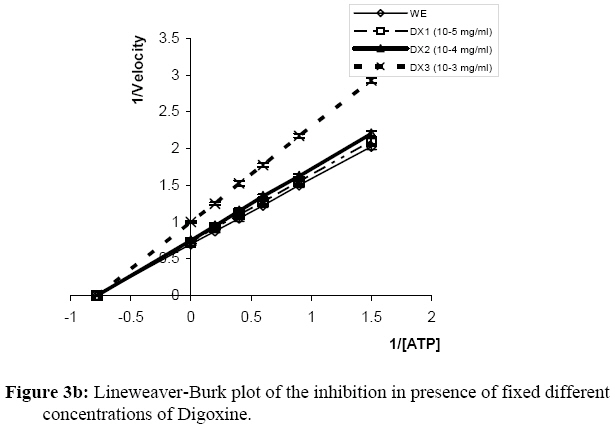
Influence Of Mitragyna ciliata (MYTA) On The Microsomal Activity Of Atpase Na+/K+ Dependent Extract On A Rabbit Heart. Bidie Alain dit Philippe (*)1., Koffi Ernest2., N’Guessan Jean-David1., Djaman Allico Joseph1., Guede-Guina Frédérique1 . 1Laboratoire de pharmacodynamie Biochimique ; UFR Biosciences Université de Cocody, 22 BP 582 Abidjan 22 Côte d’Ivoire. 2Laboratoire de Biochimie et de Sciences des Aliments ; UFR Biosciences Université de Cocody, 22 BP 582 Abidjan 22 Côte d’Ivoire. Email : alphbid@yahoo.fr Code Number: tc08042 Abstract Mitragyna ciliata (MYTA) (Rubiaceae) inhibits plasmodia activity. MYTA induces a cardiotonicity of the digitalic type on rat’s isolated heart. In this work we studied the effect of MYTA on microsomal Na+/K+ -dependant ATPase (Na+, K+ ATPase) extracted from the heart of a rabbit since digitalics inhibit Na+, K+ ATPase. Our results revealed that the Na+/K+ ATPase has an optimum pH of 7.4 and temperature of 37oC respectively. There is a linear relationship between the organic phosphate formed and the incubation time over 25 mins incubation period. The ATP hydrolysis rate in the presence of MYTA was 0.775 µM/min. LINEWEAVER and BURK plots showed that MYTA did not alter KM (1.31 mM) but decreased VMAX. This study shows that MYTA exerts a non-competitive inhibition on the microsomal Na+/K+ ATPase extracted from rabbit heart with a Ci50 of 48 µg / ml. We conclude that the mechanism of action of MYTA is linked to the inhibition of the Na+/K+ ATPase like cardiotonics of the digitalic type. Keys words: Mitragyna ciliata; ATPase Na+/K+; inhibitors of ATPase Na+/K+. Introduction Mitragyna ciliata (MYTA) (Rubiaceae) is used to treat malaria in rural areas of the Republic of Côte d’Ivoire (West Africa). The potency of the methanolic extract (from MYTA) against Plasmodium falciparum chloroquinoresistant strain FCM 29 has been documented (Djè et al., 1997). Further research proved that MYTA induced both a positive inotropic and negative chromotropic effect on isolated rat heart (Bidié, 1997, 2005). Our earlier studies have shown that Digoxine (DX), a well known digitalic, possessed similar effect in vitro (Bidié, 2005). Since digitalics exert an inhibitory effect on the Na+/K+ ATPase pump activity (Guédé-Guina, 1975,1989), we determined the effects of M. ciliata on the heart’s ATPase. The purpose was to clarify the mechanism of action of the extract of M. ciliata on Na+/K+ATPase activity. Materials and Methods Plant material Mitragyna ciliata (voucher No. 8888) is a tropical forest plant found in the Republic of Côte d’Ivoire. The active principle is found in every portion of the plant. The methanolic extract from the bark of M. ciliata (MYTA) was prepared by stirring 50 g of finely ground portions in 1,5 L methanol using a magnetic stirrer (IKAMAG RCT) for 48 hrs at 0 oC. The methanolic extract was filtered over cotton and whatman No3 filter paper three times. The filtrate was evaporated at 40 oC under reduced pressure with a rotavapor (Buchi 461 watter Batch). The residue obtained was used to prepare MYTA solutions. Animals Wistar rats weighing about 200 g and rabbits (Orictlagus cumulus) weighing about 1.5 Kg were used to run experiments on mechanical activities of the heart and for blood pressure experiments respectively. Preparation of Na+, K+ -ATPase extracts Enzyme extract was prepared by the method of Schwartz (1962) modified by Guédé-Guina (1975). The rabbit was killed and its heart was stored in a 0.32 M sucrose solution at 4 oC. The pH of the solution was adjusted to 7,4 with potassium hydroxide 1 M. About 6 g of heart muscle was homogenized in 0.32 M sucrose solution, pH 7,4 buffer in a ratio of 1:9 (w/v) in an IKA T 25 Basic homogenizer. Homogenization was conducted for 30 s at 9500 rpm with frequent stopping periods to cover the mixer with ice to prevent heat generation. The homogenate was centrifuged (MEDIFRIGER BI SELECTA). The supernatant was discarded and the microsome-rich residue was stored at – 20 oC and used as a source of Na+, K+-ATPase. The residue was resuspended in bi-distilled water at a concentration of 5 mg/ml according to the method of Nakao (1965). The enzyme extract was split into 100 ml portions and stored at –20 oC. Under these conditions optimal activity of the enzyme extract was preserved. Preparation of the ATP-Tris buffer and standard substrate ATP-tris buffer solution preparation followed a procedure of Schwart et al. (1962) and Nakao et al. (1965). The ATP standard was prepared by the method of Guédé-Guina (1975). We used a Tris (tris-hydroxy-methylaminomethane) solution containing ATP (sodium salt) prepared according to Schwartz et al. (1962) and Nakao et al. (1965). A cation exchange resin was used to prepare the ATP-Tris buffer solution. Two columns containing 2,5 g of resin (Dowex 50x4) each were washed with 50 ml HCl N then distilled water, HCl N and distilled water until a neutral pH was reached. Both columns were stored at refrigeration temperature. About 0,5 g of ATP (sodium salt, Sigma Chemical co lot n° 2383) was dissolved in 2 ml of deionised water and loaded on one of the columns. The latter was shaken for 15 min and the filtrate was removed using a Buchner under vacuum. The column was washed twice with 1 ml of water. The filtrate was added to the other column which received an identical treatment. The final filtrate was brought to pH 7,4 using Tris-HCl buffer (1 M, pH 7,4). The ATP concentration was adjusted to 15 mM by monitoring the absorbance at 260 nm using a spectrophotometer (Beckman 25). This was made possible by the fact that at 260 nm 0,015 M ATP yielded an absorbance of 0,230. Furthermore, 0,5 g of ATPNa2 yielded 50 to 60ml of ATP-Tris 0,015 M. A total volume of 100 ml of ATP standard substrate solution was prepared according to the method of Guédé-Guina (1975). The solution contained : 25 ml Tris (0,4 M, pH 7,4 ; 10 ml NaCl 1M ; 10 ml KCl 0,3 M ; 1 ml MgCl2 0,3 M and 20 ml ATP-Tris 15mM). Distilled water was added to bring the volume to 100 ml. The final concentration of the chemicals in the solution was as follows: 100 mM Tris-HCl ; 100mM NaCl ; 3 mM MgCl2; 30 mM KCl and 3 mM ATP. Analytical methods ATPase assay: ATP concentration was determined by titrimetric assay of inorganic phosphate released during ATP hydrolysis as described by Sumner (1991). This is a, sensitive and fast method for the determination of organic phosphate. Sumner method is based on the reduction of the phosphomolybdic complex by iron sulfate. In a test tube the following chemicals are added to 1 ml of purified enzyme extract as follows: 0,5 ml ammonium molybdate d’ 6,6% 0,5 ml H2S04 7,5 N 0,5 ml FeSO4 prepared immediately before use and containing (5 g FeSO4, 7 H2O, 50 ml distilled water and 1ml H2SO4 7.5 N) The colour of the reaction mixture is completed after 30 mn and the absorbance is read at 540 nm using a Spectrophotometer (Beckman 25). A standard solution (KH2PO4 5mM) is run in the same conditions. A blank is run at each trial because the method is very sensitive. Protein assay: Protein was estimated by the procedure of Lowry et al. (1951). This method uses Cu2+ ions along with Folin-Ciocalteu reagent, a combination of phosphomolybdic and phosphotungstic acid complexes that react with Cu+. Cu+ is generated from Cu2+ by readily oxidable protein components. The reaction with the Folin reagent gives intensely colored products measurable spectrophotometrically at 700 nm. Bovine serum albumin (BSA) was used as standard. BSA standards containing from 0 to 200 µg/ ml were prepared and the standard curve was conducted each time the assay was performed. Results Characterization of rabbit heart Na+/K+ dependant ATPase Our study showed that rabbit heart ATPase activity was optimum at 37 oC and pH 7,4. The inorganic phosphate released was about 1 mg which is equivalent to 0,032 mmoles using the standard curve. Results of the standard curve for proteins showed that a variation of 0,044 units correspond to 0,1 mg/ml or 100 mg/ml. ATP hydrolysis in the absence of MYTA This study was based on the release of inorganic phosphate as a by-product of ATP hydrolysis as indicated earlier. Two parameters were studied: -incubation time and enzyme concentration Figure 1a and Figure 1b show the plot of Pi formed versus incubation time for given enzyme concentrations. There was a linear increase in the amount of organic phosphate released over 25 mins of incubation time regardless of the enzyme concentration. Also, the higher the enzyme concentration the higher the initial velocity of ATP hydrolysis (Figure 2). This was expected since Vo = k [ES] = dP/dt = - ds/dt. This linear relationship between [E] and Vo allowed us to determine how much enzyme was present by measuring its activity under standard conditions without purifying the enzyme. In our study we found the standard conditions for Na+, K+ ATPase to be:incubation time 25 minutes, and enzyme concentration : 0,19 mg/ml. Figure 1a and Figure 1b and Figure 2 were used to determine the activity of our microsomal preparation. The international unit for enzyme activity is expressed in micromole of Pi released /minute /ml. The following equation was used: AE = (X / 25) x F.D; F.D : represents the dilution factor of the substrate in the total volume This study allowed us to evaluate ATPase activity in the presence and absence of MYTA. The effect of variable concentrations of MYTA on microsomal ATPase activity was investigated. The inorganic phosphate released in the experiments was monitored as a function of two parameters including the concentration of MYTA and the incubation time (0 to 30 minutes) (see Figure 3a). Our results indicate that MYTA inhibits rabbit microsomal ATPase activity. Residual ATPase activity variation was also determined as a function of MYTA concentration (Figure 4). ATPase activity decreased with increasing MYTA concentration to 130 µg/ml where the activity stabilizes at 22% of the activity in the absence of inhibitor. This residual enzymatic activity is due to Mg2+ dependant ATPase (Schwartzman, 1953; Glynn, 1957; Whittam, 1958; Skou, 1965). The Na+, K+ ATPase activity could be determined by subtracting the ATPase activity devoted to the Mg2+ dependant ATPase. The concentration of MYTA at which Na+, K+ ATPase activity decreases by 50% of its original value is called Ci50 (Ci50 = 48 µg/ ml). This Ci50 could be lower with the purified extract. Effect of substrate concentration on ATPase activity at constant MYTA concentration In this study MYTA concentration was kept constant while the substrate concentration was changed. Therefore, we investigated the influence of MYTA as a function of ATP concentration. MYTA was used at a concentration of 10-5 mg/ml. This research allowed the determination of kinetic parameters KM and VMax. Figure 3a shows the amount of Pi released as a function of ATP concentration. The curves in absence and presence of MYTA are presented and both followed MICHAELIS-MENTEN kinetics. The phenomenon of saturation is depicted when the entire enzyme is saturated with substrate. Under these circumstances the enzyme is operating at its maximum capacity and further increase in substrate concentration cannot increase the velocity any farther. From the reciprocal transform (1/V vs. 1/[S]) called LINEWEAVER and BURK plot, the two hyperbolas were transformed to two straight lines both of which intercept the x axis at the same point (-1/KM). The graphical determination of this value allowed us to calculate KM. We found KM = 1,31 mM for MYTA. We noticed that KM did not change for different concentration of MYTA (inhibitor) or in the absence of MYTA. However, VMAX changed. These results are typical of a non-competitive inhibition. Therefore, MYTA exert a non-competitive inhibition on the Na+/ K+ ATPase. Effect of Digoxine (DX) on rabbit’s heart ATPase activity This study was conducted under the same conditions as that carried out with MYTA. This comparative study reveals a similarity of effects between these two substances. Indeed, the rate of hydrolysis of ATPase Na+/K+ in the absence and presence of MYTA was determined (Figure 1b) as well as the Michaelis Menten constant KM from the Lineweaver-Burk plot (Figure 3b). On this last figure the plots had the same x-intercept (KM unaffected) but different slopes and y-intercepts (Vmax variable). Results showed that DX exerts a non competitive inhibition like MYTA on rabbit’s heart ATPase Na+/K+. In addition, residual activities (Figure 4) for both substances were not significantly different (40,3 g/ml for MYTA vs. 37,8 g/ ml for DX). Discussion This work allowed the study of the effect of MYTA on the Rabbit heart microsomal ATPase activity. Similar work was conducted on rabbit brain Na+/K+ ATPase using an extract from Mai-Bao (Yapo, 2003).Characteristics of our enzyme including optimal pH 7,4 and temperature 37 oC were determined. These parameters are similar to those found by Guédé-Guina (1975) for rabbit brain microsomal ATPase. There is a linear relationship between Pi released over 25 minutes of incubation period. The effect of MYTA on ATP hydrolysis by our Na+/K+ ATPase proceeds at a rate of 0,775 mM/min. This result shows that MYTA is an inhibitor for our ATPase. Further, reciprocal plots of LINEWEAVER and BURK were used to determine Michaelis constant (KM ). The graphical representations of our data showed that MYTA was a non competitive inhibitor for the enzyme under investigation. According to some authors (Patzelt et al., 1975), the Na+, K+ ATPase has an identical number of binding sites for Ouabain(an inhibitor for the Na+, K+ ATPase) and ATP. The location of these distinct receptor binding sites is in favour of a non competitive inhibition mechanism. Non competitive inhibitors bind to the enzyme either to a different site from the substrate, to the enzyme-substrate complex (E-S) to yield a ternary complex ESI or on both enzyme and substrate. However, the inhibition is not suppressed by increasing substrate concentration. Non competitive inhibition depends on inhibitor concentration and the affinity of the enzyme for the inhibitor. Our results show that 1/KM did not change, whereas 1/Vmax increased. Thus Vmax decreased compared to the control. MYTA’s characteristics are similar to those of digitalics such as mansonine (Guédé-Guina, 1975) and Digoxine as shown in this work. This work with MYTA reveals that this plant has interesting pharmacological potential. The plant is not only an antiplasmodic of the digitalic type but also a hypotensive agent, a myorelaxor of rabbit duodenum and a cardiotonic of the digitalic type (Bidié, 1997). MYTA’s mechanism of action is likely due to the inhibition of the cardiac sodium pump which increases the intracellular Na+ concentration, leading to stimulation of the Na+-Ca2+ exchanger which extrudes sodium in exchange for inward movement of calcium. Large amounts of Ca2+ stimulates muscles to contract. Consequently, MYTA displayed positive inotropic effects and negative chronotropic effects during bioactivity study (Bidié, 1997). This mechanism of action was also observed by Vaughan-Williams (1958), Katzung (1975), Ducouret (1976). This hypothesis is more likely because the inhibition of Na+/K+ dependant ATPase by DX follow a mechanism similar to that of many cardiac glycosides such as Sanguinin, Ouabain (Seifen et al., 1979; Biedert et al., 1979) or Strophantidin (Langer et al., 1970). Thus, the positive inotropic effect of glycosides is probably due to the lack of equilibrium between Na+-Ca2+ in favour of the Ca2+ ions following the inhibition of the Na+/K+ ATPase. Conclusion MYTA exerts a non-competitive inhibition action on Na+/K+ dependant ATPase extracted fromrabbit heart. Furthermore, MYTA's inhibitory effect on the Na+/K+ dependant ATPase pump enhance the exchange of Na+/Ca2+(1 Ca2+ for 3 Na+) in favour of inward Ca2+ stream. Consequently, contractile proteins are stimulated and cell membrane activated. The mechanism of action of MYTA is likely due to the mobilisation of the intracellular calcium, which is an important intracellular signal. References
© Copyright 2008 - African. Journal. Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc08042f2.jpg] [tc08042f4.jpg] [tc08042f3a.jpg] [tc08042f1b.jpg] [tc08042f1a.jpg] [tc08042f3b.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}