 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Traditional, Complimentary and Alternative Medicines, Vol. 6, No. 1, 2009, pp. 9-16 Research PaperApoptosis induction of Centella asiatica on human breast cancer cells Suboj Babykutty1, Jose Padikkala2, Priya Prasanna Sathiadevan, Vinod Vijayakurup, Thasni Karedath Abdul Azis3, Priya Srinivas3, Srinivas Gopala1* 1Department
of Biochemistry, Sree Chitra Tirunal Institute for Medial Sciences and
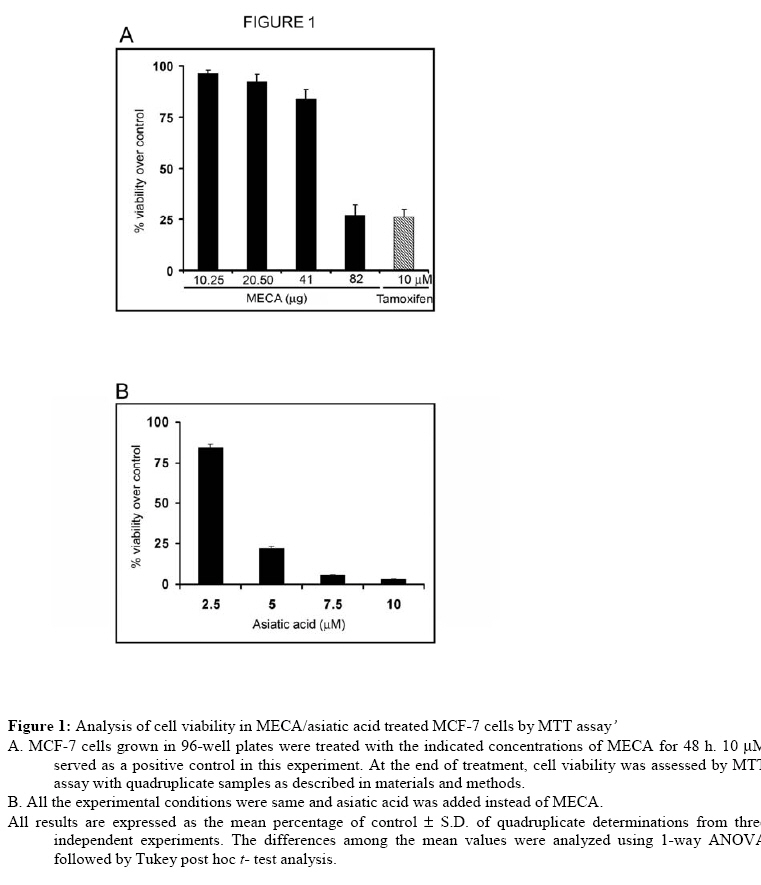
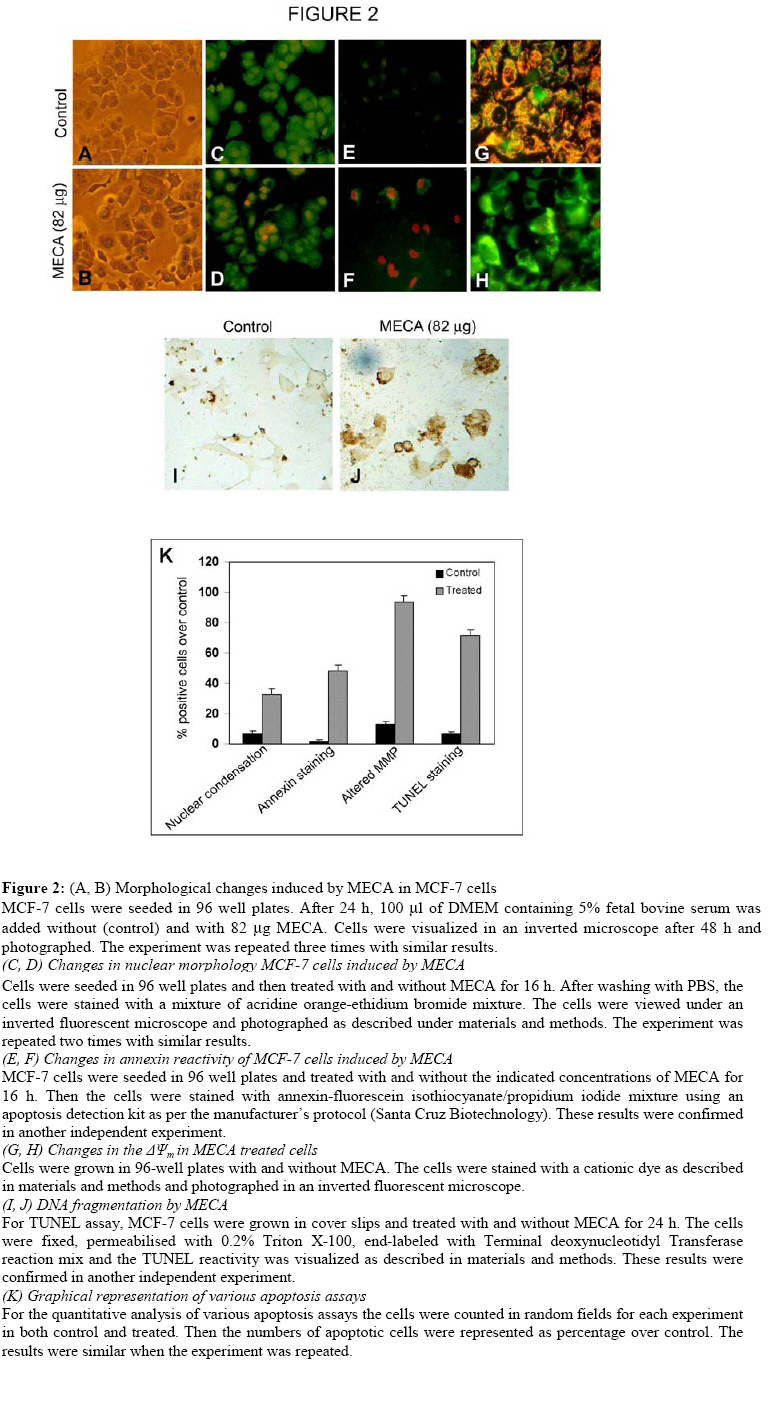
Technology, Thiruvananthapuram 695011, Kerala, India Code Number: tc09002 Abstract The present study evaluated the ability of methanolic extract of Centella asiatica (Linn) Urban (Umbelliferae) to induce apoptosis in different cancer cell lines. MCF-7 cells emerged as the most sensitive cell line for in vitro growth inhibitory activity. C. asiatica extract induced apoptosis in MCF-7 cells as indicated by nuclear condensation, increased annexin staining, loss of mitochondrial membrane potential and induction of DNA breaks identified by TUNEL reactivity. It is possible that the use of C. asiatica extract as a component in herbal medicines could be justifiable. Key words: Apoptosis, Cancer, Centella asiatica Introduction Plant based remedies have always been used as an integral part of traditional medicines throughout the world. There are many examples of potential anticancer effects of various herb/plant extracts in in vitro cell line models (Wing et al., 2007; Sebastian and Thampan, 2007; Suffredini et al., 2007; King and Murphy, 2007; Chandra et al., 2007).Centella asiatica (Linn)Urban (Umbelliferae) is one such plant used in traditional medicine for the treatment of various ailments. C.asiatica is a creeper found in wetlands of most of the Asian countries. The whole plant or the fresh leaves of the plant are widely used in ayurvedic preparations [(C.S. I. R) Council of Scientific and Industrial Research., 1950. The extract [particularly, titrated extract of C. asiatica (TECA)] has been proven as a promising candidate for improving memory, and used for cognitive disorders and for various skin ailments (Dash et al., 1996; Sampson et al., 2001; Satyavati et al., 1976; Sharma, 1992; Sivarajan, 1994). C. asiatica is used in other folklore medicines for the treatment of leprosy, ulcer, extensive wounds, eczema etc (Duke and Ayensu, 1985; Satyavati et al., 1976; Sharma, 1992).Immunomodulatory effects of C. asiatica extracts suggest its chemopreventive and antiproliferative effect (Punturee et al., 2005). Methanolic extract of the aerial parts of the plant showed in vitro antiproliferative effect in mouse fibrosarcoma cells, human liver cancer cells, human gastric adenocarcinoma cells, MK-1, murine melanoma cells, B16F10, keratinocytes, SVK-14 and in vivo tumor model test systems.(Babu et al., 1995; Bunpo et al., 2004; Lin et al., 2002; Sampsoron et al., 2001;Yoshida et al., 2005). Since apoptosis is described as the main mode of cell death induced by a variety of stimuli such as drugs, stress, radiation etc., the present study was carried out to evaluate the ability of C. asiatica extract to induce apoptosis in different cancer cell lines. The result of this study indicates that apoptosis induction by C. asiatica extract may provide clues regarding its antitumor activity in at least human breast cancer cells. Materials and Methods Preparation of plant extract The fresh plant collected was authenticated by a taxonomist from Kerala Forest Research Institute (KFRI), Peechi, Thrissur, Kerala. A voucher specimen (ACRC –14) was kept in Amala Cancer Research Centre Herbarium. The extraction procedure was done as described (Babu et al., 1995). Briefly, the whole plant C.asiatica was washed, dried under shade at room temperature and powdered. Five grams of dry powder was extracted with 50 ml of 80% methanol under stirring at room temperature overnight. The extract was filtered and the filtrate was evaporated to dryness in vacuum. The yield of the solvent free extract was 20% (i.e. 1gm). Cell culture and maintenance MCF-7, HeLa, HepG2, SW 480 cells were procured from the National Centre for Cell Science, Pune, India. The cells were grown in monolayer culture in Dulbecco’s Modified Eagle’s Medium (DMEM, Life Technologies, USA) containing 10% fetal bovine serum (FBS)(Sigma, USA) and antibiotics (100 U/ml penicillin and 100 μg/ml streptomycin) in a humidified atmosphere of 5% CO2 at 37oC. For all experiments DMEM containing 2.5% FBS was used. Cell viability assay Cell growth assays were carried out as described (Srinivas et al., 2003). We prepared different concentrations of the extract by serial dilution from the stock (82 µg/100 mL). Cells grown in 96-well microtitre plates (7000 cells/well) were incubated for 48 h with and without different concentrations of methanolic extract of C. asiatica (MECA) (10.5-82 μg). Then the medium was removed and fresh medium was added along with 20 μl of 3-(4-5 dimethylthiozol-2-yl) 2-5 diphenyl-tetrazolium bromide (MTT, 5 mg/ml) to each well. The plates were incubated for another 3 h and the formazan crystals formed were solubilized with MTT lysis buffer (20% sodium dodecyl sulfate in 50% dimethyl formamide). The plates were placed protected from light, for 2 h at 37°C in an incubator. The color developed was measured (Measuring wave length: 570 nm, Reference wave length: 630 nm) using a plate reader (BioRad, USA). Likewise, we analyzed the cyotoxicity of asiatic acid (2.5-10 µM) by the same assay. The cell viability was expressed as percentage over the control. This assay is used for the determination of cell viability and cell proliferation because it can be carried out in a microtitre plate. This miniaturization allows many samples to be analyzed rapidly and simultaneously. Acridine orange / ethidium bromide staining methods MCF-7 cells grown in 96-well plates (7000cells/well) were treated with and with out 82 μg extract for 16 h. After washing once with PBS, the cells were stained with 100 μl of a mixture of acridine orange and ethidium bromide (1:1, 4 mg/ml) solutions. The cells were immediately washed once with PBS and viewed under a Nikon inverted fluorescent microscope (TE-Eclipse 300). Acridine Orange/Ethidium Bromide Staining uses combination of two dyes to visualize cells with aberrant chromatin organization. Acridine Orange was used to visualize the number of cells which has undergone apoptosis, but it cannot distinguish viable from non-viable cells. To achieve this, a mixture of Acridine Orange and Ethidium Bromide was used. The differential uptake of these two dyes allows the identification of viable and non-viable cells. Annexin/propidium iodide staining For annexin/propidium iodide staining, the cells (7000 cells/well) were seeded in 96-well plates and treated with and without 82 μg extract for 16 h. Then they were washed with PBS and treated with 1x assay buffer, annexin-fluorescein isothiocyanate and propidium iodide as per the protocol described in the annexin V apoptosis detection kit (sc-4252 AK) from Santa Cruz Biotechnology. After 10-20 min, they were washed with phosphate-buffered saline and the greenish apoptotic cells were viewed using a Nikon fluorescent microscope and photographed.In the early stages of apoptosis, there occurs translocation of phosphatidyl serine (PS) from the inner side of the plasma membrane to the outer layer, exposing PS at the surface of the cell. Annexin binds to PS with high affinity. Similarly, Annexin V-Biotin binds in a calcium dependent manner to negatively charged phospholipid surfaces, and shows affinity for PS. Simultaneous staining of DNA (with propidium iodide) will allow the discrimination of necrotic cells from apoptotic cells. Mitochondrial membrane potential assay Mitochondrial membrane potential was measured by using a MitochondrialMembrane Sensor Kit as described by the manufacturer (ClontechLaboratories, Inc., Palo Alto, CA). After 16 h treatment with 82 μg of MECA, the cells were washed with serum-free medium. 1μl mitosensor reagent was dissolved in 1ml incubation buffer; 100 μl of it is added to the cells. Cells were then incubated at 37°Cin a humidified, 5% CO2 incubator for 15 to 20 min. Cells were washed with incubation buffer and examined with a Zeiss Axioskope 2 Plus microscope using blue filter and documented. MitoSensor aggregates in the mitochondria of healthy cells and fluoresces red. In apoptotic cells the mitochondrial potential is altered and MitoSensor cannot accumulate in mitochondria and remain in the cytoplasm as monomer and fluoresces green. Terminal deoxynucleotidyl transferase mediated dUTP nick end labeling assay The assay was carried out using in situ cell death detection kit, POD (Roche Diagnostics GmbH, Germany). Cells were cultured with cover slips and treated with and without 82 µg for 24 h. The cells were then washed with PBS and fixed in 4% paraformaldehyde. The cells were again washed with PBS and blocked with 3% hydrogen peroxide in methanol and permeabilised using 0.1% triton X 100 in 0.1% sodium citrate for 2 min on ice. The staining was performed according to the manufacturer’s protocol. TUNEL assay is a non-radioactive system designed to provide simple, accurate and rapid detection of apoptotic cells in situ at the single cell level. Statistical analysis All statistical calculations were carried out using the statistical package for social sciences (SPSS) software program (version 9.0) for Windows. All values were expressed as mean ± SD. The data were statistically analyzed using one way-ANOVA followed by Tukey’s post Hoc-t-test analysis and significant difference of means was determined at the level of p<0.05. ResultsThe study was initially done on HeLa, HepG2, SW480 (data not shown) and MCF-7 cells. Preliminary data and analysis showed that MECA predominantly showed a concentration dependent cytotoxicity to MCF-7 cells only. Therefore, further experiments were conducted on MCF-7 cells. Growth inhibitory effects of MECA/asiatic acid on MCF-7 cellsMECA and asiatic acid inhibited the proliferation of human breast cancer cell line MCF-7, in a concentration dependent manner as shown in Figure 1. LD 50 value of MECA for MCF-7 was also calculated and was found to be 66 mg. The highest concentration of the extract (82 mg) inhibited MCF-7 cell growth almost equivalent to growth inhibition obtained by 10 mM tamoxifen; a known antiestrogen drug currently used in breast cancer patients. On the contrary asiatic acid (10 µM) induced ~95 % cell death in 48 h. This shows that MECA possess only moderate cytotoxicity compared to the higher cytotoxicity of asiatic acid, one of its active components. Apoptosis induction by MECA in MCF-7 cells The phenotypic characteristics of cells treated with MECA were evaluated by microscopic inspection of overall morphology. Treatment of MECA below 41 mg did not show a significant evidence of cell death even after 24 h. Treatment with higher concentrations of MECA extract for 48 h resulted in the formation of apoptotic bodies. In contrast, cells with control medium were well spread with flattened morphology (Figure 2 A, B). Nuclear condensation –Determination by acridine orange/ ethidium bromide staining The ability of the MECA to induce apoptosis was initially screened by using acridine orange/ ethidium bromidestaining. The MECA treated cells (32.3%) showed obvious nuclear condensation after 16 h treatment. Control cells (7%) showed bright green nucleus with uniform intensity and had not taken up ethidium bromide, where the apoptotic cells appeared orange in color (Figure 2 C, D, K). Based on the above cytomorphological changes and cell death the effect of MECA in these cells were indicative of apoptosis. MECA induces annexin V binding We further confirmed apoptosis induction due to the extract with annexin V binding. It is one of the early indicators of apoptosis. Bright green annexin FITC staining was imparted to membrane of the apoptotic cells (47.8%), indicating the early stagesof apoptosis. The nuclei of cells with later stages of apoptosis exhibited red color of propidium iodide, signifying its condensed status. Control cells (1.5%) were negative for annexin FITC staining (Figure 2 E, F, K). Loss of mitochondrial membrane potential (DY) of MCF-7 cells Cells after 16 h treatment with 82 mg MECA induced a decrease in mitochondrial membrane potential and those cells fluoresce as green (93.7%). But the control cells (13.3%) turned up as red due to the higher membrane potential (Figure 2 G, H, K). This indicates that apoptosis induction by MECA involves mitochondrial pathway. MECA induces DNA strand breaks dUTP labeled 3’ OH groups of cleaved DNA indicated a reliable substantiation of MECA induced apoptosis in MCF -7 cells (71%) after 24 h of treatment. The cells without MECA treatment (6.6%) showed minimal staining by TUNEL assay (Figure 2 I, J, K). Discussion There have been so many reports showing the medicinal properties of C. asiatica extract in a wide range of disease conditions like diabetic microangiopathy, edema, venous hypertension, venous insufficiency (Incandela et al., 2001a; Incandela et al., 2001b; Incandela et al., 2001c). The role of C. asiatica extract in the treatment of memory enhancement and other neurodegenerative disorders is also well documented (Mohandas Rao et al., 2006). The first report concerning the antitumor property of C. asiatica extract was on its growth inhibitory effects on the development of solid and ascites tumor and that lead to increased life span of the tumor bearing mice (Babu et al., 1995). The authors also suggested that the extract directly impeded the DNA synthesis. In our study, C. asiatica extract showed an obvious dose dependent inhibition of cell proliferation in breast cancer cells, MCF-7. In MCF-7 cells, we could show a concentration dependent decrease in cell viability (by MTT assay) on treatment with different concentrations of C. asiatica extract. However, in other cell lines such as HeLa, HepG2 and SW 480 we did not observe a concentration dependent decrease in cell viability. We observed a higher LD50 for MECA that may be due to the synergistic action of both cytotoxic and cytoprotective components present in the extract. Our study showed nuclear condensation, a characteristic apoptotic feature visualized by Ethidium Bromide/Acridine Orange staining upon treatment with MECA. The binding of Annexin V to the phosphatidyl serine of the cell membrane emphasize the ability of the extract to initiate apoptosis. The observed loss of mitochondrial membrane potential suggests the involvement of an intrinsic pathway of apoptotic induction by MECA. DNA strand breaks induced by MECA, a characteristic feature in programmed cell death was also observed. Even though we have observed a higher LD50 value with MECA, asiatic acid (10 μM), one of the active components of MECA killed ~95% cells. The individual components of the extract may show opposing roles and it may be important in making the crude drug less effective than the isolated component. In this connection the increased cell death by means of asiatic acid may due to ROS generation (Park et al., 2005; Yoshida et al., 2005). In contrast, methanolic extract of the same plant is known to have antioxidant properties (Jayashree et al., 2003). We are unable to comment on the individual components present in C. asiatica extract responsible for the documented anticancer effects. However, we conclude that C. asiatica extract induces apoptosis in MCF-7 cells by induction of nuclear condensation, flip-flop movement of the membrane, loss of mitochondrial membrane potential and by inducing DNA strand breaks. Further investigation is essential for deciphering the molecular mechanism of action of MECA in MCF-7 and also to look whether the cytotoxicity is specific to other breast cancer cell lines as well. References
© Copyright 2009 - African. Journal. Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc09002f1.jpg] [tc09002f2.jpg] |
| |||||||||

{kind=link}
{kind=link}