 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Traditional, Complimentary and Alternative Medicines, Vol. 6, No. 1, 2009, pp. 30-41 Research PaperProtective effects of Annona muricata linn. (Annonaceae) leaf aqueous extract on serum lipid profiles and oxidative stress in hepatocytes of streptozotocin-treated diabetic rats Stephen O. Adewole+ and John A. O. Ojewole* Department of Pharmacology, School of Pharmacy &
Pharmacology, Faculty of Health Sciences, University of KwaZulu-Natal, Private
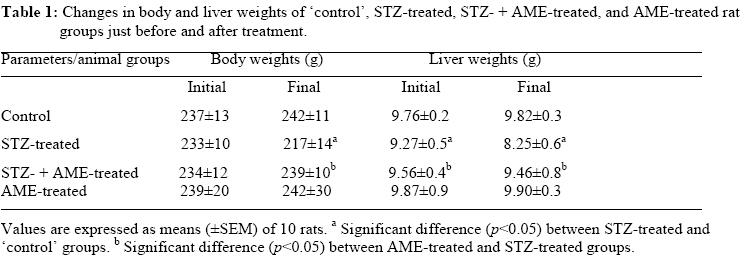
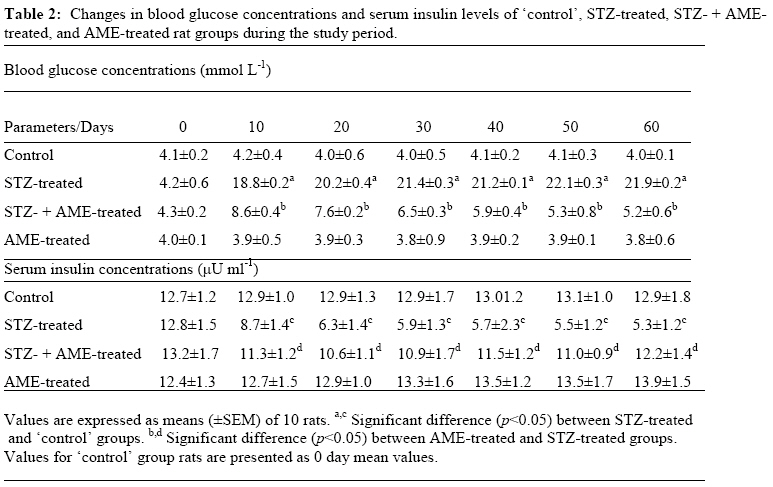
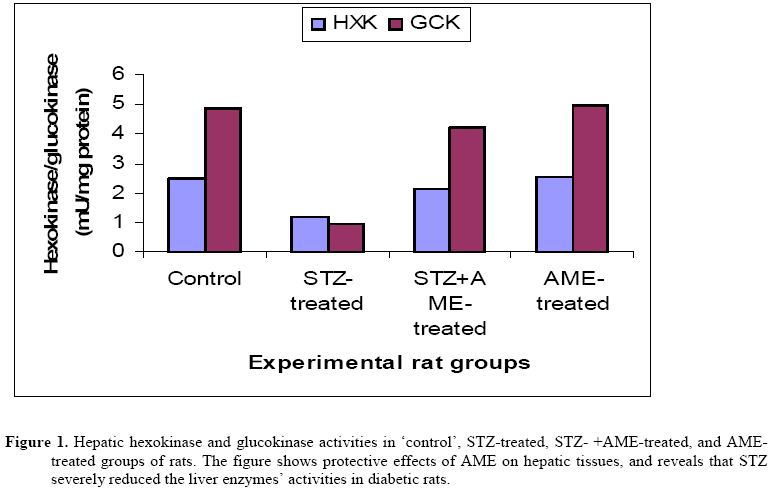
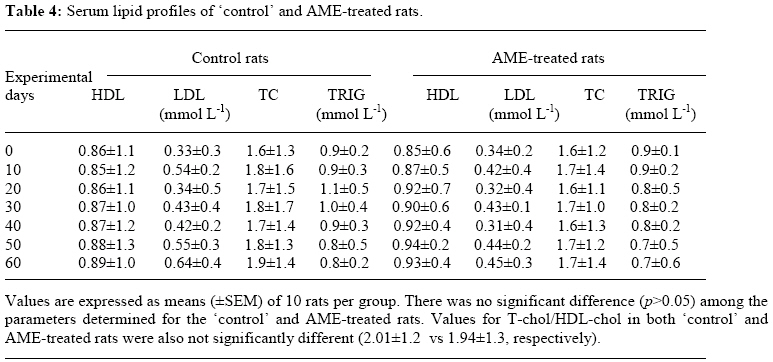
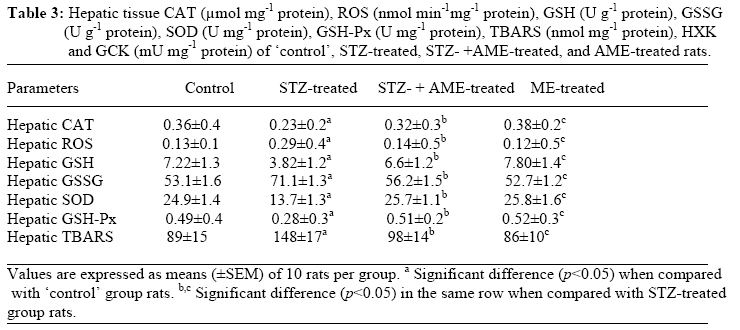
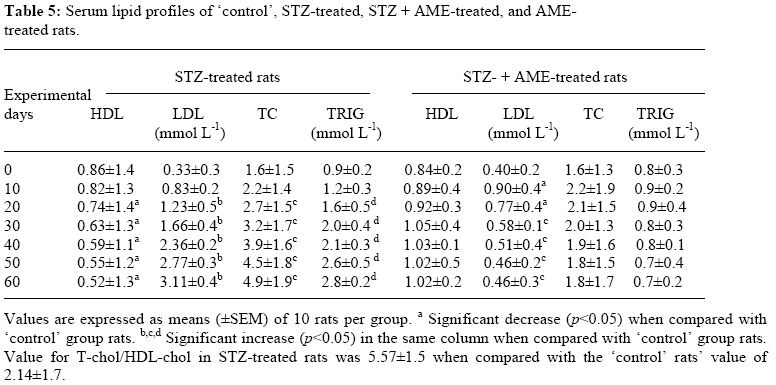
Bag X54001, Durban 4000, South Africa Code Number: tc09005 Abstract Extracts from various morphological parts of Annona muricata Linn. (Annonaceae) are widely used medicinally in many parts of the world for the management, control and/or treatment of a plethora of human ailments, including diabetes mellitus (DM). The present study was undertaken to investigate the possible protective effects of A. muricata leaf aqueous extract (AME) in rat experimental paradigms of DM. The animals used were broadly divided into four (A, B, C and D) experimental groups. Group A rats served as ‘control’ animals and received distilled water in quantities equivalent to the administered volumes of AME and reference drugs’ solutions intraperitoneally. Diabetes mellitus was induced in Groups B and C rats by intraperitoneal injections of streptozotocin (STZ, 70 mg kg-1). Group C rats were additionally treated with AME (100 mg kg-1 day-1, p.o.) as from day 3 post STZ injection, for four consecutive weeks. Group D rats received AME (100 mg kg-1 day-1 p.o.) only for four weeks. Post-euthanization, hepatic tissues were excised and processed biochemically for antioxidant enzymes and lipid profiles, such as catalase (CAT), reactive oxygen species (ROS), glutathione (GSH), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), thiobarbituric acid reactive substances (TBARS), triglycerides (TG), total cholesterol (TC), high density lipoprotein (HDL) and low density lipoprotein (LDL), respectively. Treatment of Groups B and C rats with STZ (70 mg kg-1 i. p.) resulted in hyperglycaemia, hypoinsulinaemia, and increased TBARS, ROS, TC, TG and LDL levels. STZ treatment also significantly decreased (p<0.05) CAT, GSH, SOD, GSH-Px activities, and HDL levels. AME-treated Groups C and D rats showed significant decrease (p<0.05) in elevated blood glucose, ROS, TBARS, TC, TG and LDL. Furthermore, AME treatment significantly increased (p<0.05) antioxidant enzymes’ activities, as well as serum insulin levels. The findings of this laboratory animal study suggest that A. muricata extract has a protective, beneficial effect on hepatic tissues subjected to STZ-induced oxidative stress, possibly by decreasing lipid peroxidation and indirectly enhancing production of insulin and endogenous antioxidants. Key Words: Annona muricata leaf; Aqueous extract; Lipid profiles; Streptozotocin; Oxidative stress; Antioxidants. Introduction Diabetes mellitus (DM) is one of the commonest endocrine and metabolic disorders of the 21st century, and a major threat to healthcare worldwide. Numerous experimental and clinical observations have indicated that hyperglycaemia may directly or indirectly contribute to excessive formation of free radicals (Ceriello, 2003). Diabetes is also known to involve oxidative stress and changes in lipid metabolism (Scoppola et al; 2001). The liver is the main effector organ for maintaining plasma glucose levels within narrow limits. Herrman et al; (1999) reported that streptozotocin (STZ) progressively decreased the volume of hepatocytes and their nuclei, as a result of cytoplasmic changes, and that a basal insulin level is also necessary to maintain the state of aggregation of the endoplasmic reticulum-bound polysomes for secretory protein synthesis. In insulin-deficient animals, loss of rough endoplasmic reticulum reduces amino acid incorporation into protein, and a decrease in rough endoplasmic reticulum-bound ribosomes (Lenk et al; 1992). At the same time, hyperglycaemia can generate a redox imbalance inside the cells, especially in the liver (Gallou et al; 1993). An ideal antidiabetic drug should, therefore, possess both hypoglycaemic and antioxidant properties, without any adverse effect. Increase in free radical-mediated toxicity is well documented in STZ-treated diabetic rats. Increased formation of free radicals in diabetes mellitus can be a risk factor for the disease, and it occurs as a result of two processes: (i) decreased activity of the body antioxidant systems, and (ii) auto-oxidation of reducing saccharides and formation of adducts with proteins. Antioxidant levels in the blood and tissues are important factors for sensitivity of individual tissues to oxidative stress (Durackova, 1999). Antioxidants have been classified according to their mode of action, and Bonnefont-Rousselot et al., (2000) differentiated them into three main groups, namely: (i) antioxidants that prevent the formation of new reactive oxygen species (ROS) such as caeruloplasmin, metallothioniene, albumin, myoglobin, ferritin and transferrin, (ii) scavenging antioxidants which remove ROS once formed, thus preventing radical chain reactions – these include reduced glutathione (GSH), vitamin E, vitamin C, α-carotene, uric acid and bilirubin, and (iii) enzyme antioxidants that function by catalyzing the oxidation of other molecules. This group includes superoxide dismutase (SOD) that produces hydrogen peroxide from superoxide radicals, glutathione reductase (GSH-R), glutathione peroxidase (GSH-Px) and catalase (CAT) which decompose hydrogen peroxide. Type 2 diabetes mellitus (T2DM) has been associated with an increased risk for developing premature atherosclerosis due to increase in triglycerides and low-density lipoprotein levels, and decrease in high-density lipoprotein levels. Annona muricata Linn. (Annonaceae) is commonly known as ‘Soursop’ or ‘Graviola’. Because of the ‘custard-like texture’ of its edible fruit, Annona muricata has been grouped with the ‘Custard-Apple’ plants of the Annonaceae family. It is a deciduous, terrestrial, erect tree of 5–8 metres in height, with an open, roundish canopy. Although a native of America, Annona muricata has now naturalized and become established in many tropical countries of the world. The plant has been used medicinally in many tropical African countries for an array of human ailments, especially for parasitic infections and cancer. It has also been used in some African herbal medicine systems for its sedative and antispasmodic properties. In tropical Africa, including Nigeria, the plant is generally used as antiparasitic, antispasmodic, astringent, anticancer, sedative, hypotensive, insecticide, piscicide, vermifuge, and for coughs, fevers, pain and skin diseases (Watt and Breyer-Brandwijk, 1962). The stem-bark and roots of the plant are commonly used as remedies for diarrhoea, dysentery and intestinal worms (Watt and Breyer-Brandwijk, 1962). The fruit pulp of the plant is also used in treating fevers. The unripe fruit of the plant is astringent, and is used in the treatment of intestinal atony and for scurvy (Watt and Breyer-Brandwijk, 1962). In India, the root-bark and leaf of the plant are used as anthelmintic and antiphlogistic agents, while its flowers and fruit pods are used as remedies for catarrh (Watt and Breyer-Brandwijk, 1962). Several chemical compounds have been isolated from various morphological parts (roots, stem-barks, leaves, fruits, and seeds) of Annona muricata Linn. Some of the reported phytochemicals isolated and characterized from various parts of the plant include: annonaceous acetogenins, lactones and isoquinoline alkaloids; tannins, coumarins, procyanidins, flavonoids, pentacyclic terpenoid saponins; p-coumaric acid, stearic acid, myristic acid, stepharine, reticuline, ellagic acid; phytosterols (β-sitosterol, stigmasterol), sugars, alcohols, aldehydes, organic and inorganic acids, metals, inorganic salts, vitamins B and C; stepharine, reticuline, gamma-amino butyric acid (GABA); annonacin, annocatalin, annomonicin, annomuricin, annomuricatin, corossolone, epomuricenin, gigantetrocin, javoricin, muricine, muricinine, muricapentocin, muricoreacin, montanacin, montecristin, muracin, muricatalin, muricin, murisolin, robustocin, solamin, and so on (Watt and Breyer-Brandwijk, 1962; TDRG, 2002). There are several reasons why medicinal plants should be subjected to scientific scrutiny. First and foremost, many herbal remedies have recognizable therapeutic effects (Bailey and Day 1989); but they may also have toxic side-effect (Keen et al; 1994). Recently, there has been a renewed interest in the use of plant products as antidiabetic agents. The antidiabetic effects of many traditional herbal drugs (phytomedicines) may be ascribed to their flavonoid and other chemical constituents which may also inhibit certain enzymes and possess antioxidant activities. A. muricata has a long history of usage in herbal medicine in the tropical areas of South and North America, as well as in West Africa, especially in Western Nigeria. Although all the morphological parts of the plant have been claimed to be useful in traditional medicine, no scientific studies have been carried out to establish the hypolipidaemic and antioxidant effects of the plant. Therefore, the present study was undertaken to investigate the hypoglycemic, hypolipidemic and antioxidant properties of A. muricata leaf aqueous extract in rat experimental paradigm. Materials and Methods Ethical consideration Experimental protocols and procedures used in this study were approved by the Animal Ethics Committee of the University of KwaZulu-Natal, Durban 4000, South Africa; and conform to the “Guide to the Care and Use of Animals in Research and Teaching” [Published by the Ethics Committee of the University of Durban-Westville, Durban 4000, South Africa]. Animals This study was carried out in healthy, male and female Balb C mice (Mus domesticus) weighing 20-25 g; and healthy, young adult, Wistar rats (Rattus norvegicus) of both sexes weighing 250-300 g. The animals were housed under standard laboratory conditions of light, temperature and humidity. The animals were given free access to food (standard rat pellets) and drinking tap water ad libitum. The rats were randomly divided into four experimental groups of 10 rats each: Group A (distilled water-treated ‘control’), Group B (STZ-treated), Group C (STZ-+ A. muricata leaf extract-treated), and Group D (A. muricata leaf extract-treated) rats. All the animals were fasted for 16 hrs, but still allowed free access to drinking tap water, before the commencement of our experiments. The mice were used for acute toxicity testing of the crude plant’s extract, while the rats were used for hypoglycaemic and hypolipidaemic evaluations of the plant’s extract. Plant material Fresh leaves of Annona muricata (Linn.) (family: Annonaceae) (locally known as “Soursop” or “Graviola” in English, and “Abo” in Yoruba language of Western Nigeria) were collected in Ile-Ife, Western Nigeria, between April and May, 2006. The leaves were identified by the Taxonomist/Curator of the Department of Botany, Obafemi Awolowo University, Ile-Ife, Nigeria, as those of Annona muricata Linn. (family: Annonaceae). A voucher specimen (S/N. SA003) of the plant has been deposited in the Herbarium of the University’s Botany Department. Preparation of Annona muricata leaf aqueous extract A. muricata fresh leaves were air-dried at room temperature. One kilogram (1 kg) of the air-dried leaves of the plant was milled into fine powder in a Waring commercial blender. The powdered leaf was macerated in distilled water and extracted twice, on each occasion with 2.5 1itre of distilled water at room temperature for 48 h (with occasional shaking). The combined aqueous extract solubles were concentrated to dryness under reduced pressure at 60±1oC in a rotary evaporator. The resulting aqueous extract was freeze-dried, finally yielding 36.23 g (i.e., 3.62% yield) of a light green, powdery crude aqueous leaf extract of A. muricata (AME). Without any further purification, the crude aqueous extract thus obtained was refrigerated and subsequently used in this study. Aliquot portions of the crude plant extract residue were weighed and dissolved in distilled water for use on each day of our experiments. Acute toxicity testing The median lethal dose (LD50) of A. muricata leaf aqueous extract (AME) was determined in mice using a modified method of Lorke (Lorke, 1983). Mice fasted for 16 h were randomly divided into groups of eight mice each. Stepwise, graded doses of AME (25, 50, 100, 200, 400, 800, 1600 and 3200 mg kg-1) were separately administered intraperitoneally (i. p.) to the mice in each of the ‘test’ groups. Each of the mice in the ‘control’ group was treated with distilled water (3 ml kg-1 i.p.) only. The mice in both the ‘test’ and ‘control’ groups were then allowed free access to food and drinking tap water, and observed over a period of 48 h for signs of acute toxicity. The number of deaths (caused by the extract in each group) within this period of time was noted and recorded. Log dose-response plots were subsequently constructed for the plant’s extract, from which the LD50 of the plant’s leaf aqueous extract was determined. Induction of experimental diabetes Diabetes mellitus was induced (in Groups B and C ‘test’ rats) by intraperitoneal injections of STZ (70 mg kg-1), freshly dissolved in 0.1 mol L-1 citrate buffer (pH 6.3) (Rossini et al., 1978). Diabetic state was confirmed by measuring basal blood glucose concentrations 72 h after STZ injection. Diabetes was allowed to develop and stabilize in these STZ-treated rats over a period of 4-7 days. The ‘test’ compound [i.e., Annona muricata leaf aqueous extract (AME, 100 mg kg-1 day-1 p.o.)] was administered orally by intragastric intubation to fasted Groups C and D rats. In Group C rats, administration of AME (100 mg kg-1) commenced as from the 3rd day post STZ injection, and continued for the next 4 consecutive weeks. Biochemical assays Blood Glucose and serum insulin estimations Blood samples were obtained by repeated needle puncture of the tail tip veins. Blood samples were obtained 1 day before STZ treatment, and subsequently on each other day after induction of diabetes mellitus. Blood glucose concentrations were determined by means of Bayer Glucometer Elite® and compatible blood glucose test strips. Fasted STZ–treated rats with blood glucose concentrations ≥18 mmol L-1 were considered to be diabetic, and used in this study. Serum insulin concentrations were determined by an enzyme-linked immunosorbent assay (ELISA), using a commercial kit (Crystal Chem, Chicago, IL; USA). Hexokinase (HXK) and glucokinase (GCK) activities Frozen liver tissue (1 g) was homogenized at 4oC in a 9-ml cold buffer solution (pH 7.4) containing Na-HEPES, 50 mM; KCl, 100 mM; EDTA, 1 mM; MgCl2, 5 mM and dithiothreitol (DTE), 2.5 mM; using a glass-Teflon Potter Homogenizer. The suspension formed was centrifuged at 12000 x g for 1 h at 4oC. The clear supernatant formed was used for the measurement of HXK and GCK activities by the coupled enzyme assay procedure of Davidson and Arion (1987). The incubation mixture contained the following ingredients in a final volume of 1 ml: HEPES, 50 µmol; KCl, 100 µmol; MgCl2, 7.5 µmol; and DTE, 2.5 µmol; fatty acid free bovine serum albumin, 10 mg; NAD+, 0.5 µmol; G-6-PD, 4 units; liver supernatant, 100 µl for HXK assay or 10 µL for total HXK and GCK assays; and D-glucose, 0.5 µmol for HXK and 10 µmol for total enzyme activities. Both the ‘control’ and ‘test’ tubes were pre-incubated at 25±1oC for 5 min. To the ‘control’ tubes, 0.2 ml of H2O was added, and to start the reactions in the ‘test’ tubes, 0.2 ml of a solution containing 0.5 µmol of ATP was added. ‘Control’ tubes were adjusted to zero absorbance in DU-7 spectrophotometer at 340 nm, and the increase in absorbance in the ‘test’ tubes at this wavelength was plotted against time period of 15 min. The reaction was found to be linear with time. Total enzyme activities (GCK + HXK) and HXK activities were calculated in terms of mU ml-1 of the liver supernatant. One milliunit of the enzyme corresponds to the amount of the enzyme producing 1 nmol of NADH per min under assay conditions at 25±1oC. Hexokinase activities were subtracted from the total HXK + GCK activities to obtain glucokinase activities. Protein content of the liver homogenate was determined by using bicinchoninic acid (BCA) protein assay reagent (Pierce Chemical Company, Rockford, IL, USA). Catalase activity (CAT) The activity of catalase (CAT) was measured by using its perioxidatic function according to the method of Johansson and Borg (1988). 50 µL potassium phosphate buffer (250 mM, pH 7.0) was incubated with 50 µL methanol and 10 µL hydrogen peroxide (0.27%). The reaction was initiated by addition of 100 µL of enzyme sample with continuous shaking at room temperature (25±1oC). After 20 minutes, the reaction was terminated by addition of 50 µL of 7.8 M potassium hydroxide. 100 µL of purpald (4-Amino-3-hydrazino-5-mercapto-1,2,4-triazole, 34.2 mM in 480 mM HCl) was immediately added, and the mixture was again incubated for 10 minutes at 25±1oC with continuous shaking. Potassium peroxidate (50 µL of a 65.2 mM solution) was added to obtain a coloured compound. The absorbance was read at 550 nm in a spectrophotometer. Results are expressed as micromoles of formaldehyde produced mg-1 protein. Reactive oxygen species (ROS) The amount of ROS activity in the liver was measured by using 2´,7´-dichlorofluorescein diacetate (DCF-DA), which gets converted into highly fluorescent DCF by cellular peroxides (including hydrogen peroxide). The assay was performed as described earlier by Socci et al., (1999). Briefly, the liver tissue (10 mg) was homogenized in 1 ml of ice-cold 40 mM Tris-HCl buffer (pH 7.4), and further diluted to 0.25% with the same buffer and placed on ice. The sample was divided into two equal fractions. In one fraction, 40 µL of 1.25 mM DCF-DA in methanol was added for ROS estimation. The other fraction to which 40 µL of methanol was added, served as a ‘control’ for tissue auto-fluorescence. All samples were incubated for 15 min in a 37oC water-bath. Fluorescence was determined at 488 nm excitation and 525 nm emission, using a fluorescence plate reader (Tecan Spectra Fluor Plus, Germany). Result are expressed as nmol min-1 mg-1 protein. Reduced GSH and oxidized glutathione GSSG levels Liver GSH and GSSG contents were measured as described by Hissin and Hilf (1973). To measure GSH contents, 4 ml of the liver homogenate was precipitated by adding 1 ml of a 25% metaphosphoric acid and centrifuged at 10,000 x g (Ultracentrifuge, Hitachi, Japan) for 30 min. Supernatant was diluted 20 times with the same buffer, and 100 μL of orthopthaldehyde (OPT) was added. In addition, for GSSG assay, 0.5 ml supernatant was incubated at room temperature with 200 μL of 0.04 mol L-1 N-ethylmaleimide solution for 30 min, and to this mixture, 4.3 ml of 0.1 mol L-1 NaOH was added. A 100 μL sample of this mixture was taken for the measurement of GSSG, using the procedure described above for GSH assay, except that 0.1 mol L-1 NaOH was used as the diluent instead of phosphate buffer. Samples were incubated at room temperature for 15 min and fluorescence was measured using spectrofluorometer (Tecan Spectra Fluor Plus, Germany) at 350 nm (Ex)/420 nm (Em). The values obtained were ascribed to the amount of glutathione in the liver. Superoxide dismutase activity (SOD) Liver SOD activity was assayed by the method of Kakkar et al., (1984). The reaction mixture contained 1.2 ml of sodium pyrophosphate buffer (0.052 mM, pH 7.0), 0.1 ml of phenazine methosulphate (PMS) (186 μM), 0.3 ml of nitro blue tetrazolium (NBT) (300 μM). 0.2 ml of the supernatant obtained after centrifugation (1500 x g, 10 min followed by 10,000 x g, 15 min) of 5% liver homogenate was added to reaction mixture. Enzyme reaction was initiated by adding 0.2 ml of NADH (780 μM), and stopped precisely after 1 min by adding 1 ml of glacial acetic acid. The amount of chromogen formed was measured by recording colour intensity at 560 nm. Results are expressed as units mg-1 protein. Glutathione peroxidase activity (GSH-Px) Glutathione peroxidase (GSH-Px) activity was measured by NADPH oxidation, using a coupled reaction system consisting of glutathione, glutathione reductase, and cumene hydroperoxide (Tappel, 1978). 100 µL of enzyme sample was incubated for five minutes with 1.55 ml stock solution (prepared in 50 mM Tris buffer, pH 7.6 with 0.1 mM EDTA) containing 0.25 mM GSH, 0.12 mM NADPH and 1 unit glutathione reductase. The reaction was initiated by adding 50 µL of cumene hydroperoxide (1 mg ml-1), and the rate of disappearance of NADPH with time was determined by monitoring absorbance at 340 nm. One unit of enzyme activity is defined as the amount of enzyme that transforms 1 µmol of NADPH to NADP per minute. Results are expressed as units mg-1 protein. Thiobarbituric acid reactive substances (TBARS) The product of the reaction between malondialdehyde (MDA) and thiobarbituric acid reactive substances (TBARS) was measured by a modified method of Ohkawa et al., (1979). For each sample to be assayed, four tubes were set up containing 100, 150, 200 and 250 µL of tissue homogenate, 100 µL of 8.1% SDS, 750 µL of 20% acetic acid, and 750 µL of 0.8% aqueous solution of TBA. The volume was made up to 4 ml with distilled water, mixed thoroughly and heated at 95oC for 60 minutes. After cooling, 4 ml of n-butanol was added to each tube, the contents mixed thoroughly, and then centrifuged at 3000 rpm for 10 minutes. The absorbance of the clear, upper (n-butanol) layer was measured using a Shimadzu (Japan) UV-1601 spectrophotometer at 532 nm. The concentration of MDA was calculated by the absorbance coefficient of MDA-TBA complex at 1.56 x 105 cm-1 M-1, and was expressed in µmol TBARS g-1 tissue protein. Determination of serum cholesterol, lipoproteins and triglyceride Blood samples were collected from tail tip veins of the rats after 16 h of fasting, and transferred to sterilized centrifuge tubes at room temperature. The blood samples were centrifuged for 10 min at 4,000 x g to obtain serum. The serum was stored in a freezer at 0o for later analysis of total cholesterol (TC) and triglyceride (TG), high- and low-density lipoprotein (HDL and LDL)-cholesterols. Aliquots of serum were taken for determination of total cholesterol by enzymatic colorimetric assay method of Allain et al., (1974), and triglycerides determined by enzymatic glycerol phosphate oxidase/peroxidase method of Cheng et al., (1988). Autoanalyzer (Express Plus, Ciba Corning, USA) and Elitech kit were used. Serum high density lipoprotein (HDL)-cholesterol was assayed by precipitation of chylomicrons, while very low-density lipoproteins (VLDL) and low-density lipoproteins (LDL) were determined with sodium phosphotungstic acid and magnesium chloride (Rainwater et al; 1995). Centrifugation left only the HDL in the supernatant; their cholesterol content was determined by the method of Virella-Lopes et al., (1977). Estimation of low density lipoprotein (LDL)-cholesterol was done by using empirical formula of Friedewald et al., (1972) for samples with TG levels <4.5 mmol L-1. [LDL-chol] = [Total chol] – [HDL-chol] – ([TG]/2.2); where all concentrations are given in mmol L-1. Statistical analysis The data obtained were expressed as means (±SEM), and analyzed by using repeated measures of variance. The differences between the means were analyzed statistically with one-way analysis of variance (ANOVA; 95% confidence interval), and the Bonferroni correction was applied as post hoc test. Values of p<0.05 were taken to imply statistical significance. Results Acute toxicity testing Intraperitoneal administrations of stepwise, graded doses of Annona muricata leaf aqueous extract (AME, 25–100 mg kg-1) were found to be safe in mice. However, relatively moderate to high doses of the plant’s extract (>200 mg kg-1 i. p.) were found to be toxic and/or lethal to the animals. The LD50 value of the plant’s extract was calculated to be 155±20 mg kg-1 i. p. in mice. The relatively low LD50 value of 155±20 mg kg-1 obtained probably suggests that Annona muricata leaf aqueous extract is only moderately safe in mice. Effects of diabetes on body weight and serum insulin Seventy-two hours after STZ administration, all the rats treated with STZ displayed glucosuria, hyperglycemia, hypoinsulinemia and moderate but insignificant (p>0.05) loss of body weight. At the beginning of this study, the baseline weights of all the rats were similar in all groups. At the end of the study period (60 days), however, the diabetic animals presented with significant (p<0.05) loss in body weight, as well as insignificant liver weight loss. The initial and final body weights were, however, not significantly different (p>0.05) in the ‘control’ and AME-treated rat groups (Table 1). Blood glucose and serum insulin concentrations The mean blood glucose concentrations and serum insulin levels of the STZ-treated animals are shown in Table 2. In our ‘control’ set of experiments, pretreatment of the rats with distilled water alone did not significantly modify (p>0.05) the animals’serum insulin and blood glucose concentrations. As shown in Table 2, induction of diabetes resulted in a significant increase in the blood glucose levels of the rats. There was a gradual rise in the blood glucose concentrations of the animals as from day 2 following injection of STZ, and the values were significantly higher (p<0.05) than those of ‘control’ animals (Table 2). Furthermore, high levels of blood glucose concentrations of the STZ-treated rats were persistently observed throughout the study period (22.3±0.6 mmol L-1). AME treatment significantly reduced (p<0.05-0.001) the blood glucose concentrations of the AME-treated group C diabetic rats. AME treatment also significant increased (p<0.05) serum insulin levels of the group C rats. Biochemical findings Figure 1 shows the effect of A. muricata aqueous leaf extract on hepatic hexokinase and glucokinase activities. In the STZ-treated diabetic rats, both hexokinase and glucokinase activities significantly decreased (p<0.05), but the levels returned to almost normal, after AME treatment. Table 4 shows the effects of A. muricata aqueous leaf extract on biochemical variables in STZ-treated animals. There was a clear evidence that STZ-induced hepatic injury was associated with free radical injury and oxidative stress. Oxidative stress was characterized by increased lipid peroxidation and/or altered non-enzymatic and enzymatic antioxidant systems. The effects of STZ and STZ + AME treatments on hepatic tissues’ ROS, GSH, SOD, GSH-Px and TBARS are presented in Table 3. The hepatic antioxidant activities of CAT, GSH-Px, SOD and GSH significantly decreased (p<0.05), while GSSG, ROS and TBARS significantly increased, in the STZ-treated, diabetic rats. The ‘control’ group of rats maintained optimal values of the antioxidants studied. AME treatment significantly (p<0.05) decreased STZ-induced elevated GSSG, ROS and TBARS, and also significantly increased (p<0.05) STZ-induced reduced antioxidant enzyme activities. Furthermore, AME treatment restored the altered activities of antioxidant enzymes like GSH-Px, SOD and GSH, TBARS towards their normal values in the liver. Serum total cholesterol, triglycerides, HDL and LDL cholesterols, and (T-chol/HDL-chol) in the ‘control’, STZ-treated, STZ + AME-treated, and AME-treated rats are shown in Tables 4 and 5. Serum total cholesterol, triglycerides, LDL cholesterol and (T-chol/HDL-chol) were significantly elevated (p<0.05) in STZ-treated Group B diabetic rats as compared to ‘control’ Group A rats. Similarly, HDL cholesterol was significantly reduced (p<0.05) in STZ-treated group B diabetic rats (Table 5). All the lipid parameters examined were improved towards normal values after AME treatment in Group C rats. Discussion Medicinal plants have been used for centuries in the treatment of diabetes mellitus. Therefore, we have investigated the effects of A. muricata leaf aqueous extract on lipid profiles in serum and biomarkers of oxidative stress in hepatocytes of diabetic rats. In diabetes, hypoinsulinaemia increases the activity of fatty acyl coenzyme-A oxidase, which initiates β-oxidation of fatty acids, resulting in lipid peroxidation (Baynes, 1995). Also, protein glycation and glucose auto-oxidation can lead to the formation of free radicals, and this, in turn, can induce lipid peroxidation (Baynes, 1991). Increased lipid peroxidation impairs membrane functions by decreasing membrane fluidity and changing the activity of membrane-bound enzymes and receptors (Baynes, 1995). Oxidative stress in diabetes mellitus could cause disturbances at the level of subcellular organelles, especially in the liver, which is the metabolic ‘power-house’ of the body. Evidence of mitochondrial alterations in diabetic rats has been noticed for a long time (Gerbitz et al., 1996). Mitochondrial damage can, in turn, generate a further oxidative stress inside the cell; therefore, liver mitochondria from streptozotocin-treated rats are likely to generate increased levels of reactive oxygen species (Kristal et al., 1997). Along with hyperglycaemia and abnormalities in serum lipids, diabetes is usually associated with microvascular and macrovascular complications which are the major causes of morbidity and mortality in diabetic individuals (Virella-Lopes and Virella, 2003). Diabetes can be managed by exercise, diet and drugs. Hypoglycaemic drugs are either too expensive, or possess undesirable side-effects and/or contra-indications. Therefore, the search for more effective and safer hypoglycaemic agents from plants and other natural sources has continued to be an area of interest for many researchers (Krishna et al., 2004). In the present study, we noticed elevated serum lipids in STZ-treated diabetic rats. Lipids play an important role in the pathogenesis of diabetes mellitus. The level of serum lipids is usually raised in diabetes, and such an elevation represents a risk factor for coronary heart diseases (Mironava et al., 2000). However, in this study, a significant decrease in STZ-induced elevated LDL, TG and TC; and an increase in STZ-induced reduced HDL levels, were observed in AME-treated rats. These alterations could be beneficial in preventing diabetic complications as well as in improving lipid metabolism in diabetics. The results of the present study also showed an increase in the levels of ROS, GSSG and MDA; and a decrease in GSH, CAT, GSH-Px and SOD contents of hepatic tissues of STZ-treated diabetic rats. Continuous treatment of Group C rats with AME caused significant decreases in the elevated blood glucose and ROS, GSSG and MDA levels of the diabetic rats. A significant elevation of hepatic activities of GSH-Px, CAT, SOD and GSH level were also observed in the AME-treated diabetic rats. It is thought that reactive oxygen free radicals could inactivate and reduce hepatic CAT, SOD, and GSH-Px activities. This speculation is in agreement with the findings of Wohaieb and Godin (Wohaieb and Godin, 1987). Furthermore, the decrease in hepatic GSH and increase in hepatic GSSG, could be due to decreased synthesis, or increased degradation of GSH by oxidative stress in diabetes. The marked decrease in MDA, ROS and GSSG levels in the hepatocytes of AME-treated rats probably suggests that AME exerts antioxidant activity that protects the tissues from the destructive effects of lipid peroxidation (Nicola et al., 1996). Most of the glucokinase (GCK) in a mammal is found in the liver, and GCK provides approximately 95% of hexokinase activity in hepatocytes. GCK plays an important role in diabetes. It is involved in glucose uptake in the pancreas and liver, which are defective in type 2 diabetes mellitus. Hypoglycaemia or hyperglycaemia may also reduce or alter the functional efficiency of the GCK enzyme molecule, resulting in increasing or decreasing sensitivity of β-cell insulin response to glucose (Zelent et al., 2005). Because insulin is one of, if not the most important, regulators of GCK synthesis, diabetes of all types diminishes GCK synthesis and activity by a variety of mechanisms. Furthermore, it has been shown recently that insulin has a direct stimulatory effect on mitochondrial protein synthesis in isolated rat hepatocytes (Memon et al., 1995). In the present study, GCK activity was lower in STZ-treated diabetic rats as compared with the ‘control’ rats. The decrease in hepatic GCK could result from hypoinsulinaemia, decreased synthesis, or increased degradation of GCK by oxidative stress in diabetes (Matschinsky and Magnuson, 2004). However, the ability of AME to significantly increase GCK activity of the hepatocytes to optimal level would appear to suggest insulin releasing ability of the plant’s extract in AME-treated Group C rats. The results of the present study also revealed a highly significant decrease in serum insulin levels of STZ-treated diabetic rats. Single daily doses of AME significantly reduced the blood glucose concentrations of diabetic rats, and caused a significant increase in serum insulin levels. The present data, therefore, shows that treatment of diabetic rats with AME caused marked amelioration of hyperglycaemia, with pronounced increase in serum insulin levels. Improvement in insulin action in diabetic rats after AME administration might be attributed to its ability to improve the physical state of plasma membrane through increment of hepatic GSH levels, thereby interfering with the progression of lipid peroxidation. Although the exact mechanisms of action of AME on the different biochemical variables examined in this study could not be established, a number of earlier investigators have shown that tannins and other polyphenolic compounds (e.g., coumarins), flavonoids, triterpenoid saponins, and a host of other plant secondary metabolites possess hypoglycaemic, hypolipidaemic, hypotensive, anti-inflammatory, and other pharmacological and biochemical properties in various experimental animal models (Ojewole, 2005). A. muricata is known to contain ellagic acid, tannis, flavonoids, polyphenolic compounds, triterpenoids, β-sistosterol, and so on (Watt and Breyer-Brandwijk, 1962; TDRG, 2002; Chang, 2001). It is, therefore, not unreasonable to speculate that some of the above chemical constituents of the plant, especially the coumarins, flavonoids and triterpenoids, are probably responsible for the altered biochemical variables in the hepatic tissues, as well as the antidiabetic property of AME, observed with plant’s leaf aqueous extract in this study. Conclusion Based on our findings, we conclude that STZ treatment is associated with oxidative stress in hepatic tissues, and that A. muricata leaf aqueous extract possesses antioxidant activity which is able to inhibit and/or prevent hepatic oxidative damage produced by STZ treatment. Acknowledgements The authors are grateful to Messrs Adeogun Oludele and Doherty O. Wiston for their technical assistance. References
© Copyright 2009 - African. Journal. Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc09005t5.jpg] [tc09005t3.jpg] [tc09005t4.jpg] [tc09005t2.jpg] [tc09005t1.jpg] [tc09005f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}