 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Traditional, Complementary and Alternative Medicines, Vol. 6, No. 2, 2009, pp. 139-149 Research Paper EFFECTS OF VARYING CONCENTRATIONS OF THE CRUDE AQUEOUS AND ETHANOLIC EXTRACTS OF Dalbergia sissoo PLANT PARTS ON Biomphalaria pfeifferi EGG MASSES. Adedotun A. Adenusi*1 and Alexander B. Odaibo2 1Parasitology Unit, Department of Plant Science and Applied Zoology, Olabisi Onabanjo University,
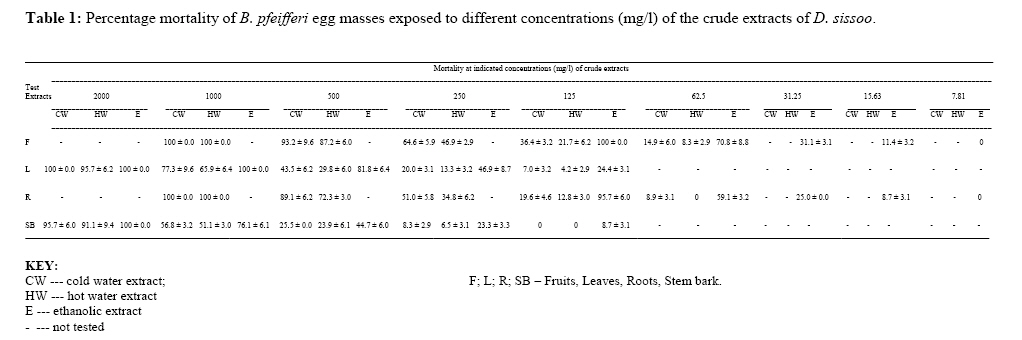
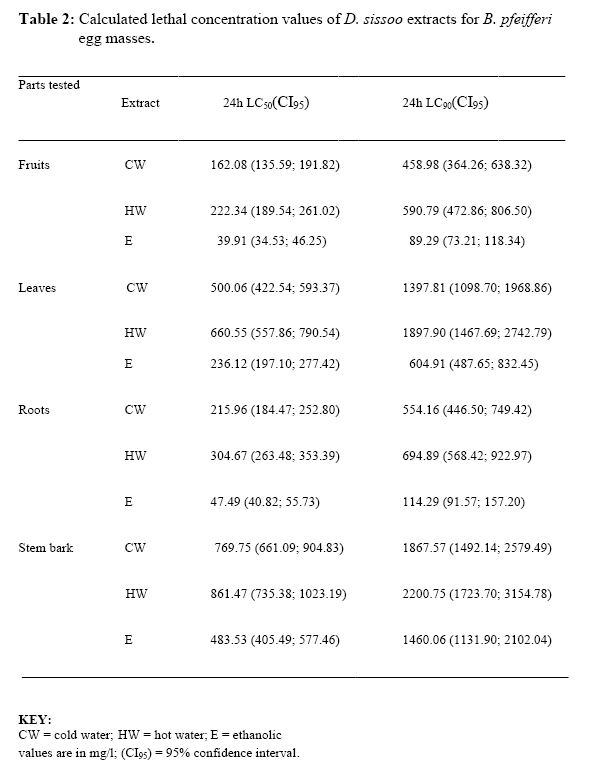
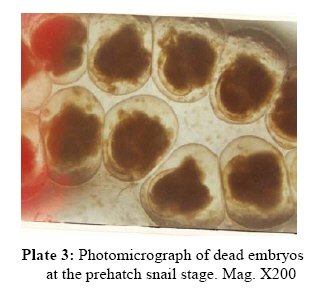
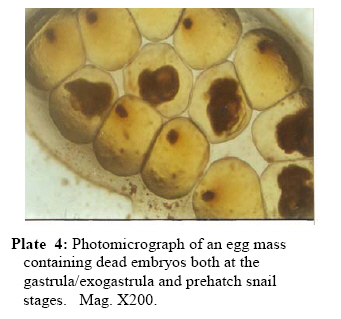
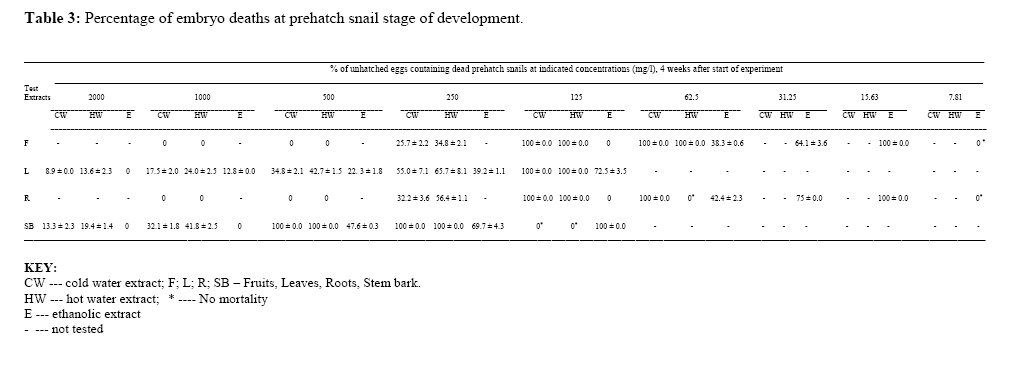
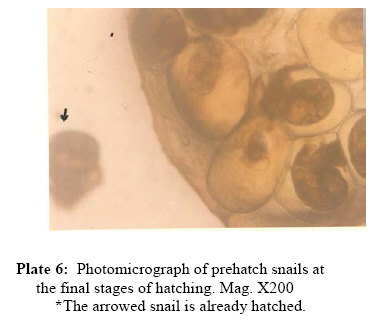
P.M.B. 2002, Ago-Iwoye. Nigeria, Code Number: tc09019 Abstract This study evaluated, using replicated laboratory bioassays, the toxicities of the crude aqueous and ethanolic extracts of Dalbergia sissoo Roxb. 1832 (family Leguminosae) fruits, leaves, roots and stem bark against egg masses of Biomphalaria pfeifferi (Krauss, 1848), the snail intermediate host of Schistosoma mansoni (Sambon, 1907) in Nigeria. Viable 0-24 hr-old embryonated egg masses were separately exposed to five different concentrations (7.81-2000 mg/l) of extracts for 24 hrs, washed in dechlorinated tap water and incubated at room temperature for a maximum of 4 weeks. The LC50 and LC90 values of test extracts for egg masses were calculated by probit analysis. The activities of the tested extracts were concentration-dependent. However, only the ethanolic extract of the fruits demonstrated significant activity (24 hr-LC90 value < 100 mg/l: 89.29 mg/l). Mortalities of eggs were manifested at the gastrula/exogastrula and or the prehatch snail stage of development. The percentage of dead embryos at the prehatch snail stage decreased while the deaths of embryos at the gastrula/exogastrula stage increased, with increasing concentration of extract. Lethality of the ethanolic extract of D. sissoo fruits to embryonated egg masses of B. pfeifferi is an added advantage to its potential development for use as a plant molluscicide, as the overall efficacy of a molluscicide is greatly enhanced if it also shows significant toxicity towards snail eggs. Key words: Dalbergia sissoo, Biomphalaria pfeifferi, Schistosoma mansoni, Egg masses Introduction Human schistosomiasis is a parasitic disease caused by digenetic trematode species of the genus Schistosoma which co-habitate the venous plexuses of the mammalian viscera (Smith and Christie, 1989; Lockyer et al., 2003) and transmitted by freshwater gastropod molluscs which serve as intermediate hosts. In the tropics and subtropics, schistosomiasis is the second most important parasitic disease after malaria in terms of prevalence, public health and socio-economic importance (James and Colley, 1995; UNDP/World Bank/WHO, 1997; Pointier and Giboda, 1999; Chitsulo et al., 2000; Steinmann et al., 2006). Five species; S. mansoni, S. haematobium, S. japonicum, S. intercalatum and S. mekongi are primarily adapted as human parasites and are endemic in different parts of the tropics where they cause significant morbidity. S. mansoni is transmitted by freshwater planorbid snails of the genus Biomphalaria which are amphimictic simultaneous hermaphrodites (WHO, 1965; Madsen, 1985; Rupp, 1996; Woodruff and Mulvey, 1997; Nakano et al., 2003). B. pfeifferi transmits S. mansoni in sub-saharan Africa including Nigeria (Rollinson and Simpson, 1987; WHO, 1993; Brown, 1994). Man, the definitive host, acquires infection by contact with freshwater infested with schistosome cercariae, which actively penetrate his intact skin (El Ridi, 2002) and subsequently develop to the adult worms. These cercariae are released into water by infected snails, in which the parasite undergoes asexual larval multiplication. The snails in turn become infected by miracidia released from schistosome eggs which reach freshwater with human excrement. With probably 500,000 deaths per year out of the over 220 million people estimated to be infected worldwide (WHO, 2001) in endemic developing countries across South America, Africa and the Far East (WHO, 1993), the World Health Organization recommends an integrated control of schistosomiasis. Thus, an appropriately targeted snail control using molluscicide(s) is an important preventive strategy which should be combined with carefully managed and directed chemotherapy, ecological and biological control methods, as well as socio-economic improvements and advances in health education with community participation (WHO, 1993; Clark et al., 1997; Yuan et al., 2000; Schall et al., 2001). Snail control in schistosomiasis is based on the rational assumption that elimination or reduction of snail population density below a certain critical threshold, would reduce transmission to a level at which the rate of new human infections, as measured by disease incidence, is significantly reduced or stopped altogether (WHO, 1965; Thomas, 1973). In addition to controlling the adult snails, control of their egg masses would also result in their eventual decimation. It is thus advantageous, if a molluscicide also kills snail eggs (Marston and Hostettmann, 1985). The high costs of synthetic molluscicides such as niclosamide, their toxicities to non-target aquatic biota and even man (Andrews et al., 1983; McCullough, 1992; Pieri et al., 1995; Rawi et al., 1996), as well as the complex organization required in their application, are a major setback to their continued use, especially in schistosomiasis control programmes. There is thus, the need for cheaper, environmentally friendly, biodegradable and readily available natural molluscicides from plants. Biologically active natural products abound in African medicinal plants (Marston et al., 1993). Phytolacca dodecandra, the best studied plant molluscicide to date, is commonly used to treat intestinal worms in Ethiopia (Esser et al., 2003), while Millettia thonningii and Pavetta owariensis, two other plant species with promising molluscicidal activities are similarly being used as anthelmintics in alternative medicine practice in Ghana (Abbiw, 1990) and Guinea (Balde et al., 1986) respectively. It is possible also, that Dalbergia sissoo Roxb. 1832 (family Leguminosae), a deciduous tree, morphological parts of which are used in alternative medicine as anthelmintic and remedy for diverse forms of ailments in Nigeria and other parts of Africa (Haerdi, 1964; Burkill, 1985, 1995) could offer greater promise for development as a plant molluscicide. In a previous paper, we reported that the crude ethanolic extracts of D. sissoo fruits and roots demonstrated promising molluscicidal activities against adult B. pfeifferi with additional toxicities towards its egg masses (Adenusi and Odaibo, 2008). This paper reports on the toxicities of aqueous and ethanolic extracts of D. sissoo fruits, leaves, roots and stem bark on further development of the embryonated eggs of B. pfeifferi. Materials and Methods Plant materials Fruits, leaves, roots and stem bark of D. sissoo were collected in the fresh state in August-September 2004 (8.00-10.00 a.m.) from the field and promptly transported to the laboratory for identification and authentication. The plant parts were independently identified in the Department of Plant Science and Applied Zoology, Olabisi Onabanjo University, Ago-Iwoye, the Herbarium Unit, Department of Botany and Microbiology, University of Ibadan and the Herbarium Unit, Forestry Research Institute of Nigeria (FRIN), Ibadan. Voucher specimens of all the plant parts were deposited at the Herbarium of FRIN (FHI 107118). Preparation of extracts All plant parts were air dried to relatively stable weights, chopped into bits and thereafter ground to moderately-fine powders (mesh size, 200μ), using an electric blender (Molyneux). Cold (room temperature; 26 to 29 oC) and hot water extracts of each plant part were prepared by steeping 40 g each, of the powdered plant material in 1L of the water, respectively for 24 hrs, followed by filtration. 4000 mg/l stock concentrations of the extracts were prepared by dilution of the filtrates with the appropriate volumes of dechlorinated tap water. These were subsequently diluted serially to final assay solutions of 2000, 1000, 500, 250, 125, 62.5 … mg/l, as required. Plant material residues were gently dried in the oven at 40 oC to stable weights. The amount of plant material in extract was expressed as a percentage of the original dry weight of powdered plant material (% Yields: Hot water extracts, 13.34 ± 0.13 – 23.56 ± 1.03; Cold water extracts, 12.50 ± 0.41 – 15.00 ± 0.44). Ethanolic extracts were prepared by steeping 40 grams of powdered plant material in 90% ethanol for 48 hrs,$ followed by filtration. Extracts (% Yields: 6.67 ± 0.37 – 8.50 ± 0.55) were concentrated to dryness by evaporation, under reduced pressure, in a rotary evaporator (NYC R-205D) at temperatures below 45 oC. Stock solutions of the extracts were prepared fresh, by dissolving 4 g of dry extract in 20 ml dimethylsulfoxide (DMSO) and then completing to 100 ml volume with dechlorinated tap water. Serial dilutions of the stock solutions with the appropriate volumes of dechlorinated tap water afforded final assay solutions of 2000, 1000, 500, 250, 125, 62.5 mg/l as required. Final concentration of DMSO in both test and control solutions was 0.5%. Assay for ovicidal activity The 0-24 hour old egg masses laid by first generation, laboratory-bred adult B. pfeifferi snails on colourless, transparent polythene sheets lining the inner walls of beakers (after 2 weeks of continuous ovipositioning), were used for the study. Egg masses were removed by cutting out small circles of polythene onto which they had attached and immediately examined with light microscopy to confirm viability (Olivier and Haskins, 1960) and extent of embryonation. Only viable eggs (Plate 1) were used for experimentation. Four egg masses (each with 8-15 embryos) were exposed for 24 hrs, to 200 mls, each of five different concentrations of an extract in 2 replicates. There were two sets of control experiments. Egg masses in the negative controls were exposed to dechlorinated tap water containing 0.5% DMSO while in order to verify the susceptibility of the egg masses under the assay conditions, those in the positive controls were exposed to 0.5 mg/l niclosamide (Bayluscide® WP 70; Bayer, AG, Germany). The 0.5 mg/l concentration of niclosamide followed a dose-range finding test (two-fold serial concentrations of 1.0, 0.5, 0.25, 0.13 and 0.07 mg/l were used) in which all embryos were killed at the gastrula/exogastrula stage at concentrations of 0.5 mg/l and above. Egg masses were removed from the extracts at the end of the 24 hr exposure period and thoroughly washed with dechlorinated tap water before incubation in beakers containing dechlorinated tap water at room temperature. Individual embryos in an egg mass were microscopically examined weekly for development and hatching. Final assessment of mortalities was at the end of 4 weeks. An embryo in an egg mass was considered dead if its cells became opaque, dull or desegregated (dos Santos et al., 2000) or if unhatched at the end of the experiment. Mortalities were recorded as the number and percentage of dead/unhatched embryos. Statistical calculation The LC50 and LC90 values (with 95% confidence limits) of extracts (with 95% confidence limits) for the egg masses were calculated by analysis of the mortality data and logarithm concentration, using a probit analysis computer software programme (SPSS v. 10.0 for Windows). Results Toxicities of tested extracts of D. sissoo, as indicated by the percentage egg mortalities (percentage of dead unhatched embryos after four weeks of incubation) were concentration-dependent and increased with increasing concentration of extracts (Table 1). Ethanolic extracts were generally more active than the corresponding aqueous extracts with lower percentage egg mortalities, at all tested concentrations (Table 1) and whose calculated 24 hr-LC50 and LC90 values for B. pfeifferi egg masses were generally higher (Table 2). Ethanolic extract of the fruits was the most active with 100% mortality at 125 mg/l, followed by those of the roots (95.7% at 125 mg/l), leaves (100% at 1000 mg/l) and stem bark (100% at 2000 mg/l) (Table 1). Their respective 24 hr-LC50 and LC90 values for B. pfeifferi egg masses were 39.91 and 89.29 mg/l (fruits), 47.49 and 114.29 mg/l (roots), 236.12 and 604.91 mg/l (leaves) and 483.53 and 1460.06 mg/l (stem bark) (Table 2). The calculated LC50 or LC90 values of the cold and hot water extracts were not significantly different (P < 0.05) from each other for each of the plant parts tested. Extract toxicities were manifested as embryo deaths either at the gastrula/exogastrula (Plate 2) or prehatch snail (Plate 3) stage of development or both (Plate 4), depending on the concentration of extract. The percentage of dead embryos at the prehatch snail stage of development however, decreased with increasing concentration of extract (Table 3), while the percentage of embryo deaths at the gastrula/exogastrula stage increased with increasing concentration of extract. Eggs exposed to 125 mg/l of either the cold or hot water extracts of the fruits, leaves and roots exclusively contained dead, prehatch snails at the end of the experiment (Table 3). None of the eggs in the positive controls hatched, even after four weeks of incubation and all were at the gastrula/exogastrula stage (Plate 5), while all eggs in the negative control groups hatched (Plate 6) by the second week of incubation. Discussion Results from this study have shown that the activity of a plant extract varies considerably according to the morphological part of the plant (Kloos and McCullough, 1987; Wickens, 1987; Sofowora, 1993). Extracts (aqueous and ethanolic) from the fruits were the most potent, followed by those from the roots, leaves and stem bark. Since active secondary metabolites do not always accumulate to the same degree in plant parts (Duncan and Sturrock, 1987; Farnsworth et al., 1987; Lugt, 1987; Wickens, 1987), their yield in a plant extract and consequently, the activity of the extract will vary considerably with the plant part. It would thus mean that in our study, the fruit extract had the highest yield of bioactive compounds. Since the yield of bioactive metabolites in a plant extract also varies considerably with the method/solvent of extraction (Marston and Hostettmann, 1985; Clark et al., 1997), it is plausible that the ethanolic extracts were generally more potent than the corresponding aqueous extracts probably because the active principles in the plant dissolved more readily in and were better extracted by a less polar solvent (ethanol) than water. The literature is replete with reports of differences in the activities of extracts obtained from the same morphological part of a plant using different solvents. For instance, the methanolic extract of the fruits of Tetrapleura tetraptera, is more potent than the aqueous extract (Adewunmi et al., 1982). Although there were no significant differences (P > 0.05) between the LC50 or LC90 values calculated for the cold and hot water extracts of each tested plant part, the cold water extracts were generally more active than the hot water extracts, judging by the mortality data. It would thus seem that the active compounds in the extracts were slightly affected by heat (Sofowora, 1993; Evans, 2002). The viable 0-24 hour-old egg masses used in this study were susceptible in varying degrees, to the toxic action of the tested plant extracts. They were however, less susceptible than the adult snails for which the calculated LC values of the extracts were lower (Adenusi and Odaibo, 2008). The higher lethal doses for embryonated eggs of B. pfeifferi and B. glabrata compared to those for the adult snails have been reported in the literature for other products, including latex of the Crown of Christ, Euphorbia splendens var. hislopii (Schall et al., 1998) and extracts from different parts of some species of Annona (dos Santos and Sant’Ana, 2001). Similarly, Ahmed and Ramzy (1997) reported that the eggs of a related species, B. alexandrina, were less susceptible to the extracts of Solanum nigrum than the adult snails, on account of higher lethal doses. In the present study, the developmental stage of the embryos at which extracts manifested lethality was concentration-dependent. Majority of the eggs exposed to the lowest concentrations of extracts hatched, while those exposed to the highest concentrations were unhatched and contained dead embryos either at the gastrula/exogastrula or prehatch snail stage of development or both. Hatching of eggs exposed to low concentrations of extracts could be due to the fact that these concentrations were not high enough to effectively interfere with normal embryonic development within the eggs, notwithstanding their entry into them. Embryo deaths at the prehatch snail stage in eggs exposed to higher concentrations of extracts could have resulted from interference with the stages involved in the process of hatching. It is plausible that following exposure, such embryos could only develop to the prehatch snail stage, beyond which subsequent developmental processes are inhibited. It is pertinent to note that the prehatch snails were actively mobile before death was a consequence of non-hatching. High as these concentrations may seem, they were not high enough to effectively arrest further development of the embryos beyond the gastrula stage. They were however, potent enough to possibly cause physiological abnormalities which, as soon as the process of hatching was about to take place, were expressed, such that the mechanisms involved in the hatching process were thwarted. Death resulted consequent upon the inability of these prehatch snails to survive further without hatching. Embryo death at the gastrula/exogastrula stage is indicative of acute extract toxicity, as such embryos never developed beyond the stages they were before exposure. This demonstrates the potency of the extracts at such concentrations, at arresting, within the 24-hr exposure period, embryonic development beyond the gastrula/exogastrula stage. Activity against both adult and egg stages of vector snails is considered one of the most important aspects of any efficient molluscicide that is to be used in the control of schistosomiasis (Tang et al., 1995). In the present study, only the ethanolic extract of D. sissoo fruits could be considered as being reasonably active against B. pfeifferi eggs (LC50 and LC90 values < 100 mg/l; 39.91 and 89.29 mg/l, respectively). The lesser susceptibility of B. pfeifferi egg masses to the toxic action of the ethanolic extract of D. sissoo fruits in the present study compared to that of the adult snails (Adenusi and Odaibo, 2008), though very important practically, does not appear to be an insurmountable obstacle to its effective use in snail control. It only means that multiple applications of the molluscicidal extract would probably be required to completely clear the snails from schistosomiasis transmission sites, as some of the eggs would persist at molluscicidal concentrations, subsequently hatch and eventually develop to mature snails capable of continuing transmission of infection. Furthermore, the molluscicide will have to be applied before the newly hatched snails start to lay eggs. Our results are reliably guaranteed by the susceptibilities of the viable 0-24 hour old egg masses to the prescribed dose of niclosamide (used as the positive control) which was within the limits reported in the literature for Biomphalaria species, including B. pfeifferi (Webbe, 1961; Shiff et al., 1970; Sarquis et al., 1997; Giovanelli et al., 2002). The findings of this study have once again re-emphasized the need to explore the possibility of using allelochemicals derived from plants, particularly those used in alternative medicine practice, as supplementary and complementary measures in the control of schistosomiasis. As the overall efficacy of a molluscicide is greatly enhanced if it also shows significant toxicity towards snail eggs, an added advantage to the potential development of the ethanolic extract of D. sissoo fruits for use as a plant molluscicide is its lethality to the embryonated egg masses of B. pfeifferi. Even Endod, the most promising plant molluscicide, is devoid of ovicidal properties (Lemma, 1970; Lemma and Yau, 1974). The practicability of the ethanolic extract of D. sissoo fruits in the control of schistosomiasis rests upon the fact that the fruit, being a regenerative plant part, does not require destructive harvesting, as would other plant parts. Conflict of interest The authors declare and affirm that there are no conflicting interests, both personal and financial, with any group of people or organisation(s). Acknowledgements The authors give special thanks to Dr. M.O. Soladoye of the Department of Plant Science and Applied Zoology, Olabisi Onabanjo University, Ago-Iwoye, Dr. A.E. Ayodele of the Herbarium Unit, Department of Botany and Microbiology, University of Ibadan, Ibadan, and Mr. Felix Usang (Late) of the Herbarium Unit, Forestry Research Institute of Nigeria (FRIN), Ibadan for identifying the plants. This study includes part of the doctoral thesis of A.A. Adenusi. References
© Copyright 2009 - African. Journal. Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc09019f3.jpg] [tc09019t2.jpg] [tc09019f6.jpg] [tc09019f2.jpg] [tc09019t3.jpg] [tc09019t1.jpg] [tc09019f5.jpg] [tc09019f1.jpg] [tc09019f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}