 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
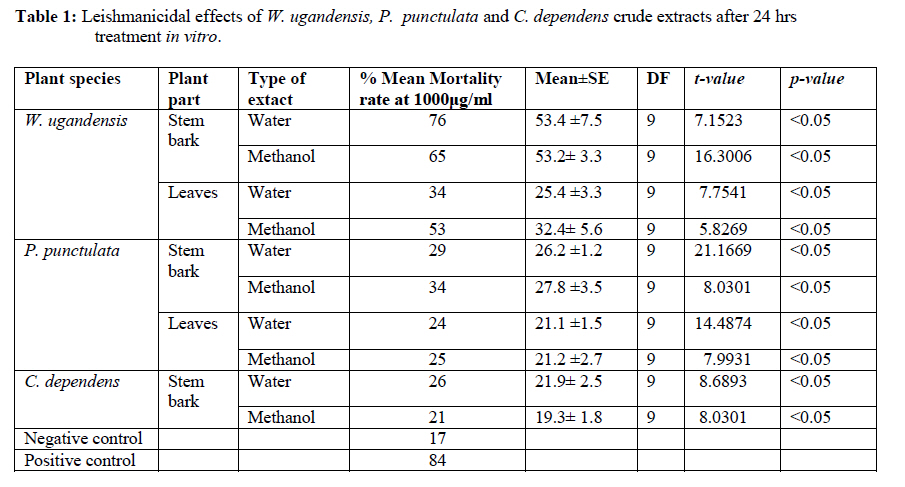
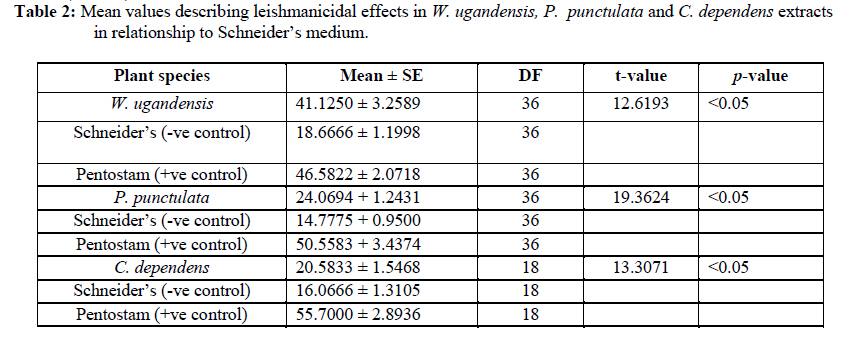
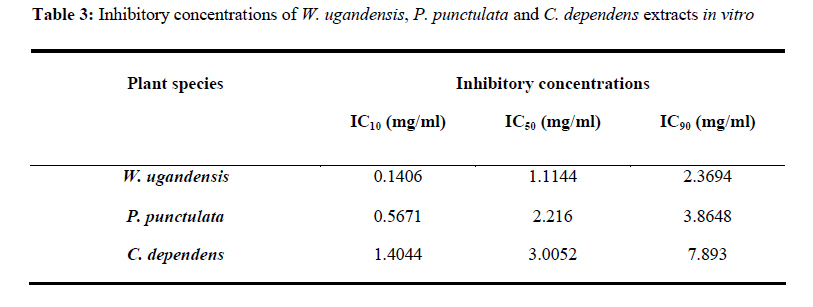
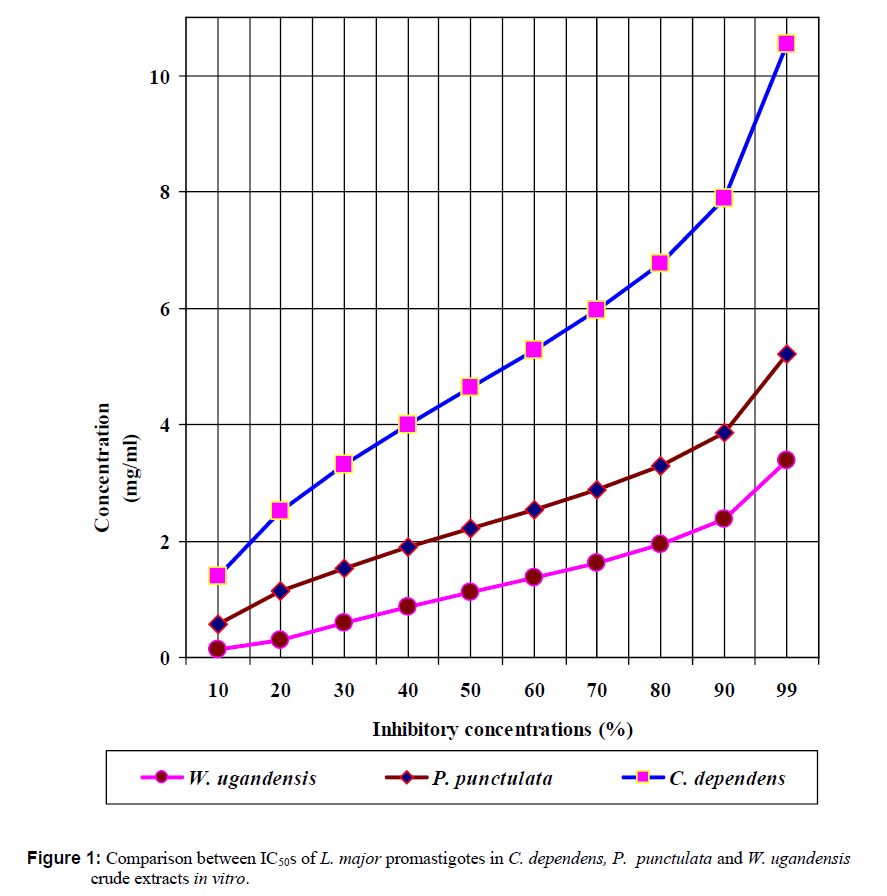
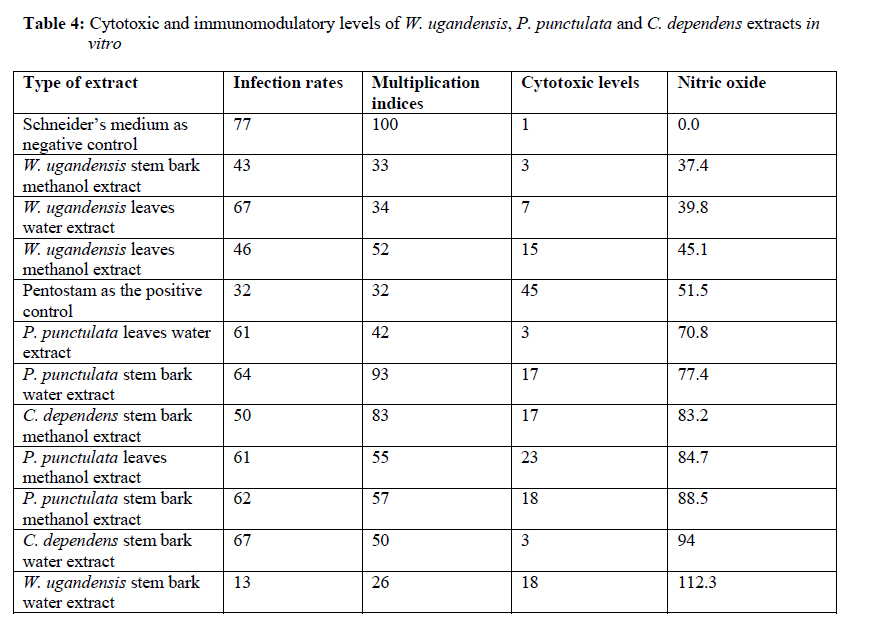
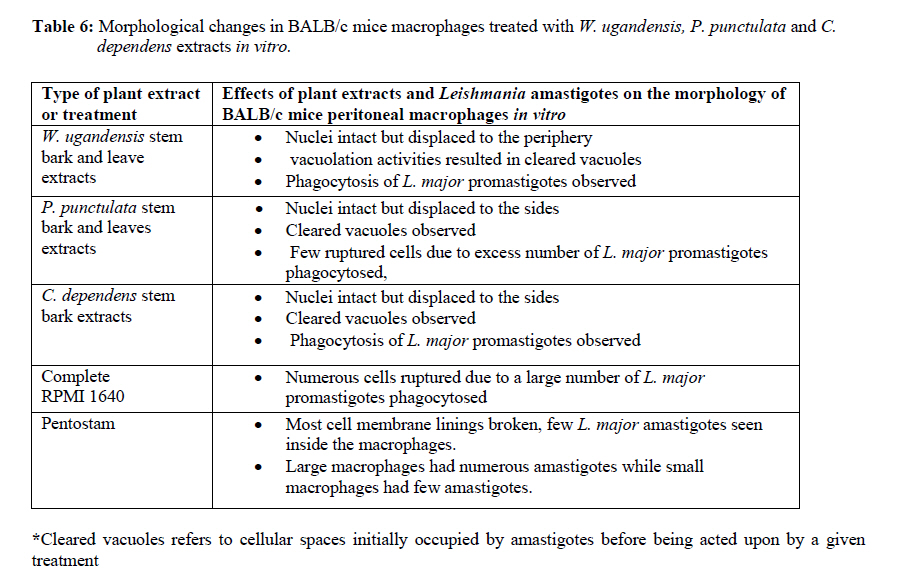
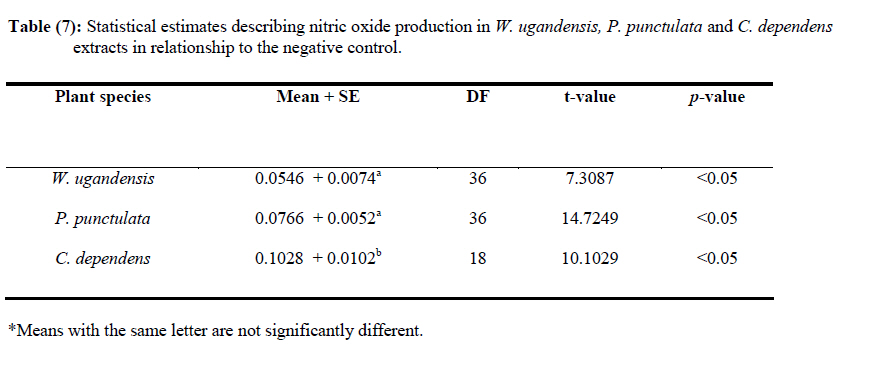
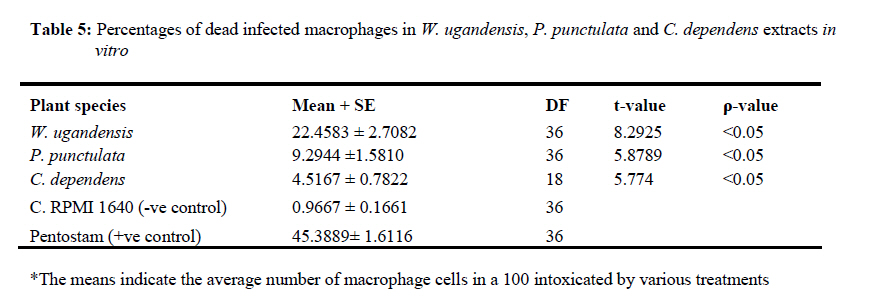
African Journal of Traditional, Complementary and Alternative Medicines, Vol. 7, No. 3, 2010, pp. 264-275 Research Paper IN VITRO EFFECTS OF WARBURGIA UGANDENSIS, PSIADIA PUNCTULATA AND CHASMANTHERA DEPENDENS ON LEISHMANIA MAJOR PROMASTIGOTES Edward K. Githinjia,b*, Lucy W. Irungua, Willy K. Tonuib, Geoffrey M. Rukungac, Charles Mutaic, Charles N. Muthaurab, Reuben Lugaliae, Geoffrey Gikandie, Caroline W. Wainainab, Johnston M. Ingongab and Antony WanjoyadaUniversity of Nairobi, P. O. Box 30197 Nairobi 00100 Kenya; Code Number: tc10037 Abstract Plant extracts from Warburgia ugandensis Sprague (Family: Canellaceae), Psiadia punctulata Vatke (Family: Compositae) and Chasmanthera dependens Hoschst (Family: Menispermaceae) were tested for activity on Leishmania major promastigotes (Strain IDU/KE/83 = NLB-144) and infected macrophages in vitro. Plants were collected from Baringo district, dried, extracted, weighed and tested for antileishmanial activity. Serial dilutions of the crude extracts were assayed for their activity against Leishmania major in cell free cultures and in infected macrophages in vitro. Inhibitory concentrations and levels of cytotoxicity were determined. Warburgia ugandensis, Psiadia punctulata and Chasmanthera dependens had an IC50 of 1.114 mg/ml, 2.216 mg/ml and 4.648 mg/ml, respectively. The cytotoxicity of the drugs on BALB/c peritoneal macrophage cells was insignificant as compared to the highly toxic drug of choice Pentostam®. The supernatants from control and Leishmania infected macrophages were analyzed for their nitrite contents by Griess reaction and nitrite absorbance measured at 540 nm. Warburgia ugandensis (stem bark water extract), Chasmanthera dependens (stem bark water extract) and Psiadia punctulata (stem bark methanol extract) produced 112.3%, 94% and 88.5% more nitric oxide than the untreated infected macrophages respectively. Plant crude extracts had significant (p<0.05) anti-leishmanial and immunomodulative effects but insignificant cytotoxic effects at 1mg/ml concentration. All experiments were performed in triplicate. Statistical analysis of the differences between mean values obtained from the experimental group compared to the controls was done by students’t test. ANOVA was used to determine the differences between the various treatment groups. The analysis program Probit was used to determine IC50s. Key Words: Warburgia ugandensis; Psiadia punctulata; Chasmanthera dependens; anti-leishmanial activity; cytotoxicity and immunomodulative effects. IntroductionLeishmaniases are a group of primarily zoonotic diseases, caused by the trypanosomatids of the genus Leishmania that are transmitted by phlebotomine or Lutzomyia sand fly species (Croft and Vanessa, 2002). Leishmaniases are now endemic in 88 countries in Africa, Asia, Europe, North and South America. Leishmaniases have an estimated 350 million people at risk; incidence of two million new cases each year and a prevalence rate of 12 million people worldwide (Ashford et al., 1992; Desjeux, 2001; Manuel and Luis, 2001). The global burden of leishmaniases caused a morbidity burden of 2.4 million disability adjusted life years (DALYs) and 59,000 deaths in 2001 (WHO, 2002). Four main forms of leishmaniases exist namely: visceral leishmaniasis (VL) or kala-azar cutaneous leishmaniasis (CL), diffuse cutaneous leishmaniasis (DCL) and mucocutaneous leishmaniasis (MCL) (WHO, 1990; Desjeux, 2004). Visceral leishmaniasis is the most severe form of leishmaniasis, involving the internal organs and is fatal if left untreated (Ashford and Bates, 1998). Unlike the other forms of leishmaniasis in which the parasite is histologically localized, VL is caused by parasite growth within reticuloendothelial cells throughout the body (Boelaert et al., 2002). It is characterized by irregular fever, weight loss, hepatosplenomegaly and anaemia (Manuel, 2002). Cutaneous leishmaniasis or oriental sores, produces skin lesions on the face, arms and legs, and is often self-healing (Ashford and Bates, 1998). DCL is a disabling disease whereas CL is only disabling when lesions are multiple (Desjeux, 2004). Mucocutaneous leishmaniasis (a mutilating disease) or espundia, as it is known in South America, causes disfiguring lesions to the face and destroys the mucous membranes of the nose, mouth and throat (Walton, 1987). Reconstructive surgery of deformities is an important part of therapy (Conjivaram et al., 2002). Traditional uses of medicinal plants consist of oral administration of crude plant extracts for visceral leishmaniasis and as topical preparations of the corresponding extracts for the treatment of cutaneous leishmaniasis (Croft and Vanessa, 2002). Natural products reported to have antileishmanial activity include quinones, alkaloids (quinolines and isoquinoline analogues, indole analogues and steroidal alkaloids), terpenes (iridoids, monoterpenes, sesquiterpenes, diterpenes, triterpenes and saponins), phenol derivatives (chalcones and flavonoids) and other metabolites such as acetogenins (Manuel and Luis, 2001). In 2001, a chloroform fraction (CLF) and a purified indole alkaloid obtained from crude stem extract of Peschiera australis was shown to have leishmanicidal effects against Leishmania amazonensis, a causative agent of cutaneous leishmaniasis in the New World. In a bioassay-guided chemical fractionation, the leishmanicidal activity in CLF completely and irreversibly inhibited promastigote growth. This fraction was also active against amastigotes in infected murine macrophages. Chemical analysis of CLF identified an iboga-type indole alkaloid coronaridine as one of its major compounds. Promastigotes and amastigotes treated with CLF or coronaridine showed pronounced alterations in their mitochondria as assessed by transmission electron microscopy (Delorenzi et al., 2001). The recommended drugs used for the treatment of visceral leishmaniasis (VL) and cutaneous leishmaniasis (CL), the pentavalent antimonials, were first introduced 60 years ago. Over the past decade alternative drugs or new formulations of other standard drugs have become available and registered for use in some countries, whilst other drugs are on clinical trial for both forms of the disease. Although the ambition to develop a single drug or drug formulation effective against all forms of leishmaniases is unlikely to be fulfilled, the advances have been significant as the concept of choice for treatment is now real. Therefore, we set out to investigate whether extracts from W. ugandensis, P. punctulata and C. dependens have inhibitory effects on L. major promastigotes in vitro. Methodology (methods and materials) Plants materials Plants were collected in Marigat Division, Baringo District in Kenya where Kala azar is endemic in August 2005. Voucher specimens; Tonui TFJ 001-5, Tonui TFJ 003-05 and Tonui TFJ 005-05 were deposited at the Herbarium of National Museum of Kenya. Authentication was achieved by comparison with Herbarium specimens by taxonomists and a voucher specimen from each plant was deposited at KEMRI. Plants extractionStem bark and leaves at the secondary stage of growth were washed to remove physical impurities, airdried, shred, and ground with an industrial blender and weighed. Water extraction was done using distilled water. Fifty grammes of ground powder was dissolved and soaked in 500 ml of water for 3 hrs, placed in warm water bath for 1 hr, filtered and freeze dried. Methanol extraction was done using distilled methanol as organic solvent, 20gms of ground powder was soaked overnight and concentrated using rotor evaporator. Two to 5gms of extract were obtained in each type of solvent and plant part. Extracts were put in sterile vials, weight recorded and stored at -20oC until required for reconstitution. Cell free, macrophage and nitric oxide assays were used to test these extracts for their different effects. Promastigotes cultivationLeishmania major (strain IDUB/KE/83 = NLB 144) which was originally isolated in 1983 from a female P. duboscqi collected near Marigat, Baringo district, Rift Valley province in Kenya, was used (Beach et al., 1984). This strain had since been maintained by cryopreservation in in vitro cultures in liquid nitrogen and also through periodic passage in BALB/c mice at KEMRI (Mbati et al., 2000). Fifteen infected BALB/c mice were used as the source of promastigotes. Stationary phase promastigotes cultures were used in this study. The mice were kept in insect proof rooms at KEMRI animal house. Promastigote forms of L. major (NLB 144) were cultured in complete Schneider’s medium prepared with 20 % heat inactivated fetal calf serum (FCS) and filtered in sterile conditions. Promastigote concentrations were diluted to 1x106cell per ml of medium. The mortality rates of promastigotes were determined using Trypan blue stain exclusion principle whereby the promastigotes permeable to the blue dye were the dead ones. The parasites were counted using a Neubauer chamber. Antileishmanial activityPromastigotes (1x106cell in 100ml) were loaded into each culture well of negative control group in complete Schneider’s media, positive control group in Pentostam and experimental groups treated with plant extracts. The 24 well plates were then wrapped with Para film and incubated in a plastic chamber at 280 C for 2-4 days, making observations in the course of the incubation. Serial dilutions at 1000µg/ml, 500µg/ml and 250µg/ml were smade from each of the stock solutions of W. ugandensis, P. punctulata and C. dependens plant extracts were then introduced into the same wells containing L. major parasites. The control group was incubated in the absence of the extracts. Parasite viability was assessed before and after incubation by staining with Trypan blue and examining under a microscope. After establishing several inhibitory concentrations, the minimum inhibitory concentration that was able to eliminate 50% of the parasites was identified as the IC50. Macrophage assayMacrophage cells were obtained from BALB/c by stimulating with 2% starch injection into peritoneal cavity with 27-gauge hypodermic needle. After 3 days the macrophages were harvested by anaesthetizing the mouse (one at time) with chloroform in an aesthesia jar with a lid, till it was unconscious, swabbing the abdomen with spirit then snipping the skin and pulling it to expose the peritoneum completely. Four millilitres of washing media Hanks buffered salt solution (HBSS) was introduced into the peritoneal cavity and the abdomen softly massaged. A lot of this media was withdrawn using 19 gauge needle into a plastic centrifuge tube placed on ice, centrifuged at 2000 rpm for 10 minutes at 40C and the supernatant was aspirated leaving about 0.5 ml in the tube. An overdose of pentobarbitone sodium (1ml) was used to euthanize the mice and their carcasses incinerated. Concentrations of macrophages was adjusted to 105 live cells per ml. 0.5 ml of the suspension was added into each cell-culturing chamber and incubated at 37 oC, 5% CO2, and 95% humidity for 24 hrs after which 1 ml of reconstituted test compounds was introduced to the experimental group of cultures at 250 µg/ml, 500 µg/ml and 1000 µg/ml concentrations and not into the control group. The negative control group was incubated in complete RPMI 1640 media while the positive control group was incubated in Pentostam. Both the experimental and the controls were incubated for further 48 hrs, after which the experiment was terminated, dried, fixed stained and macrophages counted. Percentages of infected macrophages were determined by counting 100 cells in triplicate slides. Cytotoxic levels were determined by counting the number of ruptured and/or blue macrophages (the dead ones) in a hundred. The study was carried out according to the institutional, national and international rules in regard to animal experiments and biodiversity rights. Infection of macrophages and nitric oxide productionMouse peritoneal macrophages were cultured in 24 well plates for 72 hrs. Adherent cultured macrophages were washed once and re-suspended in fresh culture medium. Dead parasites were removed from the complete RPMI 1640 medium by centrifugation (1,000 x g, 5 min), and intact Leishmania promastigotes in their stationary growth phase were added to the macrophage culture plate wells at a ratio of 10 promastigotes to 1 macrophage for 4hrs. The parasite-macrophage interaction studies were performed at 37oC. After 4hrs, the wells were washed again to remove unphagocytosed parasites. The infected macrophages were then incubated with the test compounds for further 72 hrs, after which the supernatants were tested for NO production levels. Griess reaction testThe supernatants from control and Leishmania infected macrophages were analyzed for their nitrite content by Griess reaction (Green et al., 1990). One hundred microlitres of supernatant was put in the 96 well plates. Sixty microlitres of Griess reagent A (1% sulfanilamide in 1.2N HCL) and 60 µl of Griess Reagent B (0.3% N- [1naphthyl] ethylenediamine) were added to the supernatant, mixed thoroughly and incubated in the dark at room temperatures for 10 minutes. Plates were then read at 540 nm in an enzyme-linked immunosorbent assay plate reader (Ignacio et al., 2001). Sodium nitrite (NaNO2) in RPMI was used to construct a standard curve for each plate reading. The nitrate content of samples was determined by comparison of the nitrate standards and nitrate standard curve (Green et al., 1990; José, 1998). Results Antileishmanial activity The water and methanol extracts of the stem bark of W. ugandensis caused 76% and 65% mortality rates at a concentration of 1000 µg/ml respectively as shown in Table 1. At 1000 µg/ml, Pentostam, the current drug of choice caused 83% mortality rates in L. major promastigotes in vitro. A 17% mortality rate was observed in L. major parasites cultivated in complete Schneider’s medium. Mean mortality rates in P. punctulata and C. dependens were lower compared to W. ugandensis. Stem bark extracts in W. ugandensis and P. punctulata registered higher mean mortality rates than leaves (Table 1). Table 2 shows that mortality rates caused by W. ugandensis and Pentostam were in the same range across the three concentrations. But extra cellular antileishmanial potential in W. ugandensis stem bark water and methanol extracts exceeds that in Pentostam. Mortality rates in L. major promastigotes exposed to P. punctulata and C. dependens were second and third highest while mortality rates in complete Schneider’s medium were the lowest. The difference between mortality rates in W. ugandensis, P. punctulata and C. dependens treated experimental groups of L. major promastigotes and the negative control ones was statistically significant hence the plants’ extracts had leishmanicidal effects (Table 2). Warburgia ugandensis had an IC10 of 0.1406 mg/ml with a 95 % confidence interval and IC90 of 2.3694 mg/ml respectively (Table 3). A concentration of 1.1444 mg of W. ugandensis crude extract per ml was able to kill 50 % of the promastigotes in vitro. Psiadia punctulata had an IC10 of 0.5671 mg/ml, IC50 and IC90 of 2.2160 mg/ml and 3.8648 mg/ml respectively. Chasmanthera dependens extracts had IC10 of 1.4044 mg/ml, IC50 and IC90 of 3.0052 mg/ml and 7.893 mg/ml respectively (Table 3). Figure 1 below indicates that 2mg/ml, crude extracts of W. ugandensis were able to cause 77% parasite clearance rate while P. punctulata and C. dependens could only cause 39% and 10% parasite clearance rate in vitro. Chasmanthera dependens was the weakest crude extract; killing 90% of the L. major promastigotes in vitro at 7.893mg/ml. Psiadia punctulata killed 90% of the L. major promastigotes in vitro at a lower concentration of 3.864mg/ml (Figure 1). Macrophage assay for cytotoxic effects Infection rates: Warburgia ugandensis leaves and C. dependens stem bark water extracts had the highest mean infection rate of 7 amastigotes per macrophage while the lowest infection rate was observed in W. ugandensis stem bark water extract at 1 amastigote per macrophage cell at a concentration of 1000mg/ml (Table 4). Macrophages in complete Schneider’s media and Pentostam had a mean infection rate of 8 and 1 amastigotes per macrophage cell respectively (Table 4). Multiplication index: Chasmanthera dependens stem bark methanol extract had the highest multiplication index of 83% at 1000 µg/ml hence 8 amastigotes per macrophage, down from the initial 10 promastigotes per macrophage at infection stage (Table 4). Most of the plant extracts inhibited the rate at which amastigote forms multiplied inside macrophages. Stem bark water and methanol extracts of W. ugandensis caused 74% and 68% inhibition. Cytotoxic levels: Crude extracts of W. ugandensis, P. punctulata and C. dependens had effects on the morphology of BALB/c macrophages after 72 hrs exposure in vitro. Such macrophages had nuclei intact but displaced to the periphery, cleared vacuoles and some of them raptured. Warburgia ugandensis, P. punctulata and C. dependens registered low numbers of intoxicated blue and purple macrophages that were below 23% at a concentration of 1000µg/ml. Warburgia ugandensis extracts had a mean number of 10 dead macrophages in a hundred at 1000µg/ml concentration. Psiadia punctulata leaves methanol extract registered a mean of 23 dead macrophages in a 100 macrophages at 1000µg/ml concentration. P. punctulata stem bark methanol extract was 18% cytotoxic. Psiadia punctulata stem bark water and methanol extract and P. punctulata stem bark water extract had a mean number of less than 18 dead macrophages in a hundred at 1000µg/ml concentration. An average of 1 purple or blue stained intoxicated macrophage in a 100 was found in cultures grown in complete Schneider’s medium. C. dependens stem bark methanol killed 17 out of a 100 BALB/c peritoneal macrophages at 1000µg/ml concentration. W. ugandensis stem bark methanol and C. depedens stem bark water extracts had a mean count of 3 cells each while Pentostam mean counts were the highest at 45 cells in a 100 cells at 1000µg/ml. Table 4 shows that the difference in cytotoxic levels between plants extracts and the control was significant p<0.05. Therefore, cytotoxic effects of plant extracts in macrophage cells were not significant. Crude extracts of W. ugandensis, P. punctulata and C. dependens had effects on the morphology of BALB/c macrophages (Table 6). The colour of the macrophage cell after staining, integrity of the cell membrane, number and distribution of L. major promastigotes were the parameters used to determine whether a cell was intoxicated or not (Table 6). Nitric oxide production for immunomodulative effects Infected BALB/c peritoneal macrophages treated with 1000 µg/ml of W. ugandensis stem bark water extract produced 112.3% more nitric oxide than in infected macrophage cells cultured in complete Schneider’s media in the negative control group (Table 4). Chasmanthera dependens stem bark water and methanol extracts produced 94.0% and 83.2% more nitric oxide than the negative control, respectively. Macrophages in C. dependens extracts, P. punctulata extracts and W. ugandensis stem bark water extracts produced over 50% more NO than in the negative control group while Pentostam treated cells produced 51.5 %. Levels of nitric oxide in C. dependens were the highest among the three plant species studied. The differences between optical densities in W. ugandensis and P. punctulata and C. dependens treated infected macrophages were significant as compared with optical densities in untreated infected macrophages. Means in W. ugandensis and P. punctulata were significantly different to the mean in C. dependens extracts (Table 7). In W. ugandensis and P. punctulata extracts, there were significant positive correlations between mortality rates in cell free assay and nitric oxide production levels in NO assay as shown in Table (7). There was insignificant correlation between mortality rates in C. dependens extracts and nitric oxide production levels. Chasmanthera dependens stem bark water and methanol extracts produced 94.0% and 83.2% more nitric oxide than the negative control, respectively. DiscussionAll the three species of W. ugandensis, P. punctulata and C. dependents showed significant anti-leishmanial and immunomodulative activity but insignificant cytotoxic levels at the highest concentration of 1000μg/ml. Strength in activity was dependent on species of plant, part of plant and type of solvent used during extraction. Epipolygodial, mannitol, muzigadial, polygodial, tannin, uganensidial, ugandensolide and warburganal are examples of alkaloid group of metabolites present in the bark of W. ugandensis (Bekalo et al., 1996). A range of biological effects, such as trypanocidal, fungicidal, antiviral and antibacterial activity has been ascribed to them. This paper describes for the first time the activity of W. ugandensis against promastigote forms of L. major. The antileishmanial activity of a different isoquinolinic alkaloid, isoguattouregidine isolated from Guatteria foliosa (Annonaceae), causes a total lysis of L. donovani, L. amazonensis and L. donovani (Manuel and Luis, 2001). In those studies, IC100 against the promastigotes of the three species was 100μg/ml, a dose approximately thirty eightfolds smaller than found here for W. ugandensis crude extracts against L. major (3787μg/ml). Leishmanicidal effects in W. ugandensis extracts were dose and part of plant dependent due to differences in concentrations of the various active metabolites. The stem bark extracts had a higher leishmanicidal effect than leaves extract. A sesquiterpene characterized as muzigadial, isolated from W. ugandensis showed trypanocidal activity against Trypanosoma brucei in vitro (Olila et al., 2001). Trypanosomes are evolutionary close to Leishmania parasites hence likely to respond similarly to similar ethno botanicals (Maslov et al., 2001). It has been confirmed that the intracellular mammalian stage amastigote is intrinsically more drug sensitive than promastigote stage of L. major (Croft and Vannessa, 2002). More recently a screen of plant products identified a broad-spectrum antileishmanial extract, PX6581, which was active against Leishmania amastigotes at 20ng/ml but had no activity against promasigotes. The active antileishmanial ingredient has been identified as a titerpene saponiside. A single parenteral dose of 1mg/kg was sufficient to clear established L. donovani infections in mice. Two 1mg/kg doses cured L. panamensis and L. mexicana lesions in mice (Maes et al., 2000) Use of relatively drug insensitive promastigotes stage in our study probably explains why inhibitory concentrations by the crude extracts were high. PX6518 is currently in pre-clinical development. Probably in vitro tests on these plant products using amastigotes rather than promastigotes can produce even lower inhibitory concentrations. Extra cellular antileishmanial potential in W. ugandensis stem bark water and methanol extracts exceeds that in Pentostam, although the latter is also known for killing intracellular Leishmania parasites indirectly by activating macrophage mitochondrial mechanisms such as NO production (Philippe et al., 2002). Oval shaped slender L. major promastigotes forms with short less rigorous flagella after 24 hrs of treatment with W. ugandensis crude extract suggests that the extracts interfered with the normal growth and development of the parasites. Polygodial’s ability to inhibit the mitochondrial ATPase has been demonstrated (Lunde and Kubo, 2000). The results on cytotoxicity in this study imply that crude extracts from W. ugandensis, P. punctulata and C. dependens caused no significant adverse effects in BALB/c mice peritoneal macrophages in vitro. We, therefore, suggest that the leishmanicidal efficacy at these concentrations was not due to in vitro cytotoxicity. Weak toxicity towards macrophage host cells and strong activity against the extracellular promastigotes and intracellular amastigote form of Leishmania were observed for stem bark extracts in W. ugandensis in vitro. However a cytotoxic sesquiterpene, characterized as muzigadial, was isolated from W. ugandensis (Olila et al., 2001). It was highly toxic in the brine shrimp assay. Our study did not indicate what concentration W. ugandensis extracts is cytotoxic to infected BALB/c peritoneal macrophages. Factors such as part of plant and type of extract did not affect levels of toxicity. The positive control (Pentostam) cytotoxic levels were statistically insignificant thus confirming its already known toxic effects (Grogyl et al., 1992). Warburgia ugandensis is a medicinal plant whose bark has been used for ages as treatment against stomach worms and malaria in Baringo (Henke, 1994). Decoction of leaves of P. punctulata is used as a remedy for colds, headache and abdominal pains (Kokwaro, 1993). Maasai use a decoction of leaves to rid calves of fleas (Henke, 1994). Known active ingredients in P. punctulata include flavones and phenyl propenoids (Agnew and Agnew, 1994). It’s used for treating venereal disease, general tonic for physical and nervous debilities (Maurice et al., 1999). In C. depedens, the known active ingredients are berberine type alkaloids, palmatine, colombamine and jateorhizine (Maurice et al., 1999). It has anti-inflammatory and analgesic effect (Morebise et al., 2001). Consumption of these extracts by various communities in the world is a hint about their non-cytotoxic effects to humans. The water and methanol extracts of the stem bark of W. ugandensis had the lowest mean infection rates less than five L. major promastigotes phagocytosed by each macrophage. These findings suggest that W. ugandensis extracts were either quick in causing death to the extra cellular parasites before they were phagocytosed, or impaired the phagocytosis process. The transformation of flagellated promastigotes to nonflagellated amastigotes in the macrophage phagolysosomal compartment of the mammalian host is a critical step for the establishment of infection (Zhang and Matlashewski, 1997). Phagolysis of engulfed parasites could have led to cleared vacuoles observed in infected macrophage cells as reported in Table 5. Electron microscopy on treated infected macrophages could have shown detailed ultrastructural changes in nucleus, endoplasmic reticulum, Golgi complex and mitochondria. A reduction in infection rate was evident in all the plant extracts hence an indication that they suppressed either the establishment or the development processes of L. major promastigotes in BALB/C peritoneal macrophages. Concentration dependent multiplication indices in all plant extracts imply that high concentrations inhibited rate of replication in amastigotes inside macrophage cells. Lowest levels of inhibition in P. punctulata stem bark and leaves, water and methanol and C. dependens stem bark confirms their weak chemotherapeutic potential. Nitric oxide production in macrophage activities was determined as biomarkers for immunostimulation (Philippe et al., 2002). Cytostatic and/or cytotoxic activities by activated macrophages on various parasites depend on nitric oxide production. Warburgia ugandensis stem bark water extract produced 112.3% more nitric oxide than the negative control. This means that apart from the normal nitric oxide usually produced by Leishmania infected macrophages naturally, the crude extracts from W. ugandensis actually stimulated the macrophages in vitro, to produce more nitric oxide to kill the foreign bodies indirectly. Warburgia ugandensis stem bark water extract produced more than twice as much nitric oxide than negative control at 1000µg/ml concentration. Pentostam, an immunomodulator also produced more nitric oxide than the control. The extracts of W. ugandensis could have acted indirectly by activating macrophage effector’s functions and that certain configurations entered the host cell and remained active even in the intracellular environment. The use of polygodial to facilitate the transmembrane transport of exogenous chemicals into cells has been successfully experimented (Kubo and Taniguchi, 1988). Stimulation of NO production by W. ugandensis could be due to different alkaloids present in this extract. Psiadia punctulata stem bark methanol and water extracts and P. punctulata leaves methanol and water extracts produced over 70.8% more nitric oxide than the negative control. Apart from the normal nitric oxide usually produced by Leishmania infected macrophages naturally, the crude extracts from P. punctulata actually stimulated the macrophages in vitro, to produce more nitric oxide to kill the foreign bodies indirectly. Methanol soluble components of P. punctulata extracts were more immunomodulative than the water soluble ones. Production of more nitric oxide in C. dependens stem bark water and methanol extracts probably means that apart from the normal nitric oxide usually produced by Leishmania infected macrophages naturally, the crude extracts from C. dependens actually stimulated the macrophages in vitro, to produce more nitric oxide to kill the foreign bodies indirectly. Both water and methanol soluble active compounds in C. dependens extracts were responsible for the cell death process induced by NO against the intracellular protozoan L. major parasite. Significant positive correlation between mortality rates in L. major promastigotes and nitric oxide production levels in infected macrophages in W. ugandensis and P. punctulata extracts probably suggests that both direct leishmanicidal activity and indirect killing of the parasites through activation of macrophages to produce NO could have contributed to the overall antileishmanial effects. Most of the killing of parasites in C. dependens was as a result of either direct leishmanicidal effects or indirectly through NO production by infected macrophages but not through both mechanisms of selective cytotoxicity. ConclusionsThe results of the present study confirmed that natural products are potential sources of new and selective agents for the treatment of important tropical diseases caused by protozoans. The killing of L. major promastigotes by all the three plant species was dependent on part of plant, type of extract and its concentration. Warburgia ugandensis, P. punctulata and C. dependens crude extracts have antileishmanial properties. W. ugandensis stem bark water and methanol extracts, P. punctulata stem bark methanol extract and W. ugandensis leaves water extract had the highest significant (p<0.05) leishmanicidal effect on L. major promastigotes in vitro therefore they are potential chemotherapeutic agents for leishmaniasis. Warburgia ugandensis, P. punctulata and C. dependens plant extracts had no significant (p<0.05) cytotoxic effects on infected BALB/c macrophages in vitro at 1000µg/ml, thus safe for use in further experimental studies in vivo. Warburgia ugandensis stem bark water extract, P. punctulata stem bark and leaves methanol extracts and C. depedens stem bark water extract had the highest significant (p<0.05) levels of nitric oxide produced by infected macrophages hence they are strong immunomodulatory agents in vitro. AcknowledgementsWe thank Dr. Davy Koech, Director of Kenya Medical Research Institute (KEMRI) and Dr. Gerald Mkoji, the Director of the Centre for Biotechnology Development and Research for availing the Leishmaniases laboratory and other facilities of KEMRI for our use. Financial and/or material assistance from the International Federation for Science (IFS), University of Nairobi (UoN) in conjunction with Kenya Medical Research Institute (KEMRI) is gratefully acknowledged. Logistical and technical support provided by KEMRI Leishmaniases laboratory staff is also highly appreciated. Many thanks also go to Prof. Lucy Irungu, Dr. Willy Tonui for supervising the work. Part of this work made up the M.Sc. research thesis of the main author. References
Copyright 2010 - Afr. J. Trad. CAM The following images related to this document are available:Photo images[tc10037t4.jpg] [tc10037t1.jpg] [tc10037t5.jpg] [tc10037t7.jpg] [tc10037t2.jpg] [tc10037t6.jpg] [tc10037t3.jpg] [tc10037f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}