 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Traditional, Complementary and Alternative Medicines, Vol. 8, No. 2, 2011, pp. 196-206 BIDIRECTIONAL FUNCTION OF SHENGHE POWDER ON REPAIR OF RADIATION-INDUCED DNA DAMAGE IN GLIOMA AND ASTROCYTE Yuesheng Xia,* 1 Yongqi Li, *1 Jianhua Wang, 1 Huan Wang, 2 Jing Kang,3 Baoying Wang3 and Zengxia Hou4 1Xijing Hospital, Fourth Ministry Medical University,
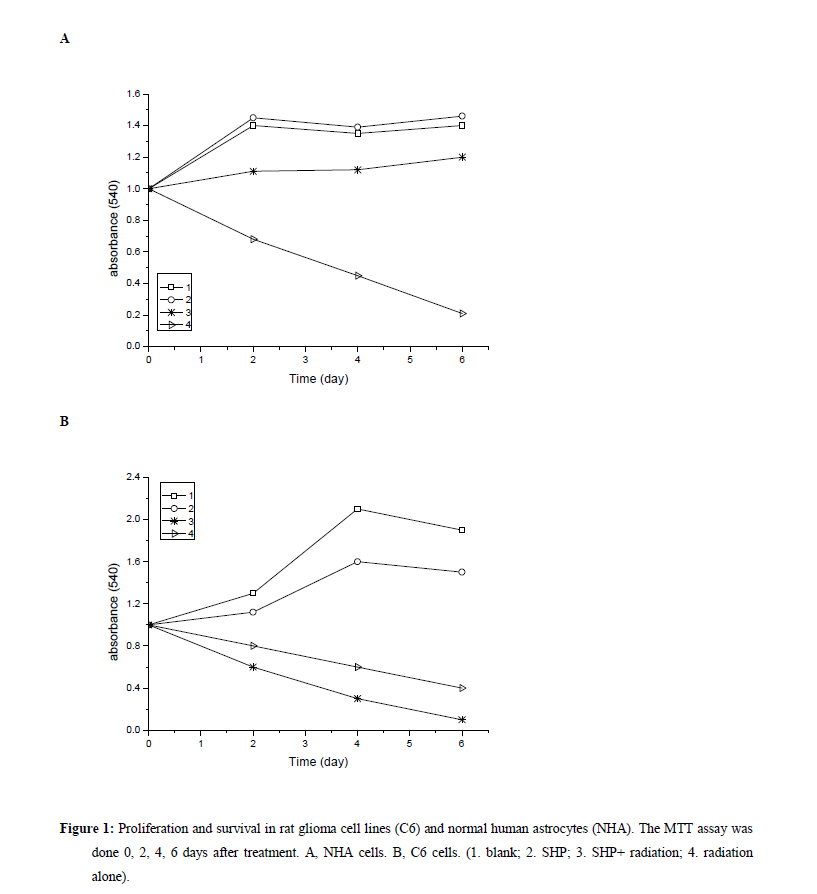
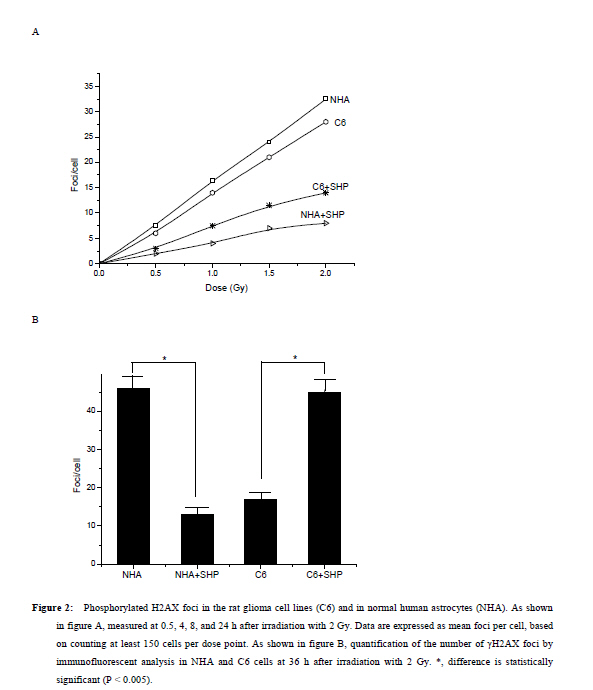
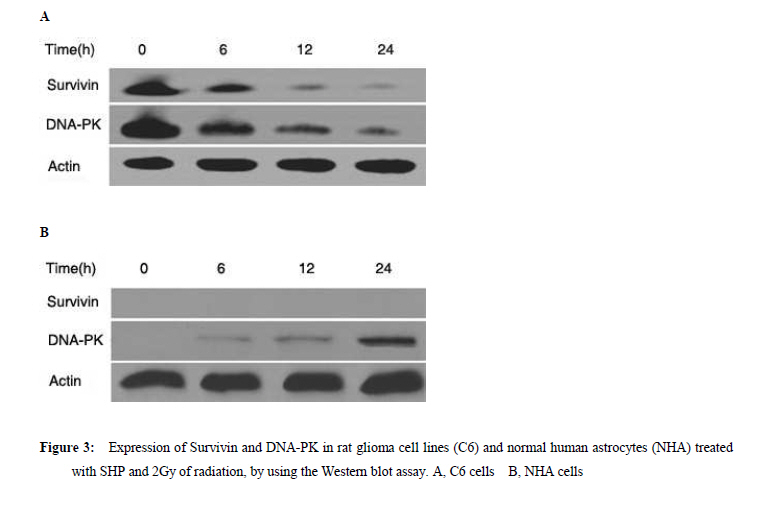
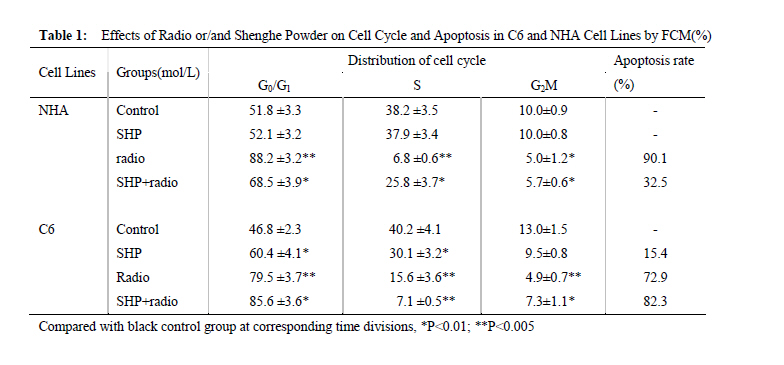
Xi’an City, Shaanxi Province, 710032, China. Code Number: tc11027 Abstract The study assessed the effect of Chinese herbs of Shenghe Powder (SHP) on the repair capacity of gamma-radiation-induced DNA damage in rat glioma cells (C6) compared with normal human astrocytes (NHA). C6 and NHA Cells treated with SHP and irradiated with 2Gy of gamma radiation. Cells growth inhibition were analysed by MTT assay, DNA damage and repair were evaluated using phosphorylated histone H2AX (γH2AX) at the appointed time. Apoptosis was observed by flow cytometry, and the expression of DNA-dependent protein kinase (DNA-PK) and surviving proteins were assessed by Western blot analysis. SHP depressed the radiation-induced DNA double-strand break and enhanced the DNA repair capacity in NHA, which correlated with promotion of DNA-PK phosphorylation. In contrast, SHP enhanced radiosensitivity of C6 cells, the pre-treatment with SHP resulted in reduced numbers of γH2AX foci in irradiated C6 cells, and decreased the expression of DNA-PK and survivn(P<0.005). It significant effect on inhibition of C6 cell proliferation and induced C6 cells apoptosis in a time-depdendent manner than radiation alone (P<0.001). SHP showed a novel bidirectional function to improve the radioresistance of NHA and enhanced radiosensitivity of C6 cells. This implies that SHP can protect the NHA from radiant damage and enhanced the sensitivity of C6 cells to radiation, which could be attributed to the alteration of survivin DNA-PK in DNA repair processes. Keywords: DNA repair, radiation, glioma, astrocyte, Chinese herbs IntroductionHigh-grade glioma is common in primary malignant brain tumor, standard therapy consists of surgical resection followed by radiotherapy, despite the use of conventional therapeutic modalities, the prognosis in patients is very poor (Fogh et al., 2010; Arslan et al., 2006). Attempts to improve outcome by increasing the local radiation dose have in general been unsuccessful because of necrosis in the surrounding brain, which becomes dose limiting before significant improved local tumor control is achieved. On conventional radiation dose, the development of resistance of glioma to radiation is a major problem during the treatment of tumors (Clarke et al., 2009; Stupp et al., 2007). Obviously, a more effective approach would be to develop agents that selectively sensitize these glioma cells to radiation. Studies approved DNA damage and repair capability related to radiation-induced cytotoxicity and radiosensitivity (Manti and D'Arco, 2010). In this study, we examined DNA damage responses and repair in rat glioma cell lines (C6) and normal human astrocytes (NHA) treated with Shenghe powder (SHP) and radiation, and attempted to demonstrate its mechanism. Materials and Methods MaterialsRat glioma cells was purchased from the Second Military Medical University, normal human astrocytes were purchased from Cambrex Bio Science (Wokingham, UK). 3-[4,5-dimethylthiozol-2-yl]-2,5-diphenyltetrazolium bromide (MTT, Sigma Chemicals, St Louis, MO), anti-γH2AX antibody (Upstate Biotechnology, USA), Survivin antibody (Novus biological, Littleton, CO, USA), DNA-dependent protein kinase (DNA-PK) antibody (Upstate Biotechnology, Lake Placid, NY). SHP mainly includes Radix Ginseng, Scrophularia ningpaensis Heml, Atractylodes macrocephala Koidz, Lacrymajobi Linn. var. ma-yuen (Roman.) Stapf, Herba Hedyotis Diffusae, Herba Cistanches, Bufo gargarizans, and Glycyrhiza uralensis Fisch. All Chinese herbs were purchased from Shanxi Company of Chinese Herbal Medicines and identified by professor Yuesheng Xia. Methods SHP Extraction Method Previously described methods was used (Wang et al., 2007). The dried whole herbs (1000g) were degreased by heating under reflux with industrial ethanol in a bath and then extracted 3 times with 10 L boiling distilled water for 1 hr each time. The decoctions were then filtered through carbasus, mixed, and concentrated to 1000 ml. The concentrated extract was mixed with 95% ethanol to make the ethanol content up to 80%. After standing overnight (20±2h), the precipitate was filtered, washed, and vacuum-dried to give a brown power. Before experimentation, it was dissolved in phosphate-buffered saline (PBS) and filtered with a 0.22-μm membrane. The percolate was made up to concentrations of 1.0mg/l and stored at –20°C. Cell Culture C6 and NHA cells were grown in RPMI 1640 (Life Technologies, Inc., Rockville, MD) containing glutamate (5mM) and 5% fetal bovine serum, and maintained at 37 °C in an atmosphere of 5% CO2 and 95% room air. SHP Treatment and IrradiationThe exponentially grown cell lines C6 and NHA were treated with SHP (1mg/l) for 2 hr. Then, culture plates were cooled on ice and irradiated with 0~2 Gy of gamma radiation at a dose rate of 0.78 Gy/min (137cesium irradiator, Atomic Energy, Ottawa, Canada). SHP was not removed from the medium. After irradiation, the NHA and C6 cell lines were placed back into the incubator (37 °C) and the kinetics of DNA repair was assessed, control cells were subjected to similar treatment but without irradiation or SHP. MTT AssaysC6 and NHA Cells were plated in a 96-well plate and treated with lower concentrations of SHP (1mg/l), and some were exposed to radiation. After treatment, cells were stained with MTT, and incubated for 4 hr in a 37 °C incubator. Then the cells were lysed in 150µL of ethanol/DMSO mixture (1:1), and the absorbance read at 540 nm using a 96-well plate reader. Immunofluorescent Analysis for γH2AX An approach similar to method of Camphausen et al. (2004) was adopted. C6 and NHA cells were cultured on coverslips placed in 35-mm dishes. Fractional cells were exposed to 1 mg/l SHP for 24 hr at 37 °C, after which time the cells were rapidly washed three times with PBS and the cells incubated in fresh medium for 30 min to allow time for phosphorylation of histone H2AX. Other cells were were exposed to 1 mg/l SHP for 2 hr at 37 °C and then irradiated with 0, 1, or 2 Gy and incubated for a further 30 min. The medium was removed, and the cells were washed with PBS and then held in methanol/PBS (50:50 v/v) at room temperature for 10 min before fixing them in methanol at –30 °C for 30 min. After removal of the methanol, the cells were held in PBS at room temperature for 5 min and then blocked with 5% milk powder in PBS for 30 min. The cells were then stained with antiphospho-histone H2AX antibodies for 1.5 hr at room temperature in the dark. After removal of the primary antibody, the cells were washed with PBS, followed by 0.1% Tween 20 in PBS and three more times with PBS, and then incubated with Alexa Fluor 488-labeled goat anti-mouse IgG antibody (Molecular Probes, Eugene, OR) at room temperature in the dark for 45 min. The cells were again washed with PBS, followed by 0.1% Tween 20 in PBS and three more times with PBS and rinsed with water. The coverslips were mounted on microscope slides with 4µl mounting solution (1mg/ml p-phenylenediamine, 3µg/ml 4',6-diamidino-2-phenylindole [DAPI], 90% glycerol in PBS) and stored at 4 °C in the dark. Fluorescent foci were imaged with a Zeiss LSM510 Laser Scanning Confocal microscope (Carl Zeiss, Jena, Germany) mounted on a Zeiss Axiovert 100M microscope. Images were captured by a Photometrics Sensys CCD camera (Roper Scientific) and imported into IP Labs image analysis software package (Scanalytics, Inc.). For each treatment condition, γH2AX foci were determined in at least 300 cells. Western Blot Assay Lysates were generated by placing these cells in RIPA lysis buffer (for Suvivin) or Chaps Lysis buffer (for DNA-PK). Western blotting assays were performed to determine total protein concentrations, which were normalized to 1 µg/µl for all samples. Samples were then prepared in sample buffer and heated to 95 °C for 5 min. Protein lysates (15 ml) in sample buffer from each tissue were loaded within each well. Gels were run at constant current (40 mA) for 3–4 hr for maximum separation. Wet transfer was performed for 4 hr at constant voltage (40 V) using polyvinylidene difluoride membrane presoaked in methanol. The membrane was blocked in 5% milk in 0.2% tetrabutyl antimony (TBST). The membrane was then washed in 0.2% TBST×3 for 15 min each. The membranes were then incubated overnight with primary antibodies directed at survivin and DNA-PK. Subsequently, the membranes were washed in 0.2% TBST×3 for 15 min each. The membrane was then incubated with secondary antibody for 45 min. Chemiluminescent (Bio-Rad, Hercules, CA, USA) detection was then used to detect expression of survivin, DNA-PK, respectively. For each gel, positive and negative controls were loaded for survivin or DNA-PK, respectively. Actin levels served as internal loading controls. Measurement of Apoptosis Annexin-V StainingPhosphotidylserine exposure on the outer layer of the cell membrane was measured using the binding of annexin V–fluorescein isothiocyanate (FITC). Cells were harvested and washed with cold PBS, incubated for 15 min with annexin V–FITC and propidium iodide and analyzed by flow cytometry (Becton Dickinson, Franklin Lakes, NJ, USA). Flow Cytometric Analysis Cells were grown in 6-well plates and were treated as indicated. Then, attached and floating cells were pooled, pelleted by centrifugation, washed in PBS, and fixed with cold 70% ethanol containing 0.5% Tween 20 at 4 °C overnight. Cells were washed and resuspended in 1.0 ml of propidium iodide solution containing 100 lg of RNase A/ml and 50µg propidium iodide/ml and incubated for 30 min at 37 °C. Apoptotic cells were assayed using FACSort Becton Dickinson Flow Cytometer at 488 nm and data were analyzed with CELLQuest Software. Cells with sub-G1 propidium iodide incorporation were considered as apoptotic. The percentage of apoptotic cells was calculated as the ratio of events on sub-G1 to events from the whole population. Statistical Analysis Data are presented as mean (±SD, or SE). Mean values were compared by the Student’s t test. The threshold of statistical significance was set at P < 0.05. Statistical analyses were performed using SPSS 17.0 package (SPSS, Chicago, IL, USA). Results Proliferation AssayC6 and NHA cells were treated with SHP at a 1mg/l, 24 hrs before single-dose irradiation (0, 1 or 2Gy).The MTT assay was used to determine the effect of SHP on C6 and NHA cell survival and proliferation at 0, 2, 4, and 6 days after cell irradiation. As figure 1 showed, both cell lines when irradiated, but not treated with SHP, showed a time-dependent decrease in cell survival and proliferation compared with cells that received no radiation (P<0.005), the C6 cell line seemed more radioresistant than the NHA cell line. NHA cell lines treated with SHP and radiation showed a time-dependent decrease in cell proliferation before 2 day, which gradually presented radio-resistance compared with cells that received radiation alone after 2 day (P < 0.005)(Figure 1A). Figure 1 displayed the largest decline in cell survival which occurred between 2 and 4 days after radiation. Both cell lines treated with SHP did result in an opposite results. C6 proliferation and cell viability decreased significantly after SHP, the greatest decrease in cell survival and proliferation occurred in the fourth day (P < 0.001). In contrast, NHA treated with SHP reversed the shape of the time-dependent decrease curve and upgraded the curve. It shows that SHP may inhibit the repair of DNA damage in C6 and enhance the repair of DNA damage in NHA. H2AX Phosphorylation in Glioma and Normal Astrocytes CellsImmunofluorescence was used to visualize and quantify phosphorylated H2AX foci in C6 and NHA cells at various times after irradiation in vitro. These foci are thought to represent sites of DNA double-strand breaks (DSBs) where chromatin structural change is occurring. H2AX phosphorylation is recognized to be an early event in break repair and has a role in subsequent recruitment of repair proteins. Foci resolution is thought to occur following DSBs repair, and persistent foci at late time points are thought to represent residual unrepaired DSBs (Mah et al.,2010; Entin-Meer et al., 2007). By comparing the number of foci per cell across the dose range of interest over a repair time from 30 min to 24 hr, we can compare induction and resolution of DSBs and therefore define DSBs repair kinetics in these cell lines. We used the assay in the C6 and NHA cells examined at 0.5, 4, 8, and 24 hr after irradiation, when resolution of foci was occurring. These data are expressed as foci resolution with time; a representation of DSBs repair kinetics is shown in Figure 2. Clearly much higher amounts of γH2AX foci were observed in NHA control group and C6 treated group. All cells tested showed radiation dose–dependent increase in residual γH2AX foci reflecting unrepaired DNA double-strand breaks. However, SHP pretreatment further increased the number of residual γH2AX foci in C6, and continued to increase steadily up to 24 hr. Higher amounts of γH2AX foci were observed in C6 compared with NHA cells (P<0.005). The remaining foci at 24 hr are thought to represent unrepairable damage, which occurred at a rate of approximately two lesions per cell per gray. The data indicate that in C6 cells, more DSBs were produced in C6 and left unrepaired, whereas in NHA cells, less DSBs were produced and/or were subject to repair. The data are in line with the enhanced sensitivity of C6 cells compared with NHA cells. Express of DNA-Dependent Protein Kinase and SurvivinDNA-PK activity has been shown to be essential for DNA repair and cell survival, previous studies have demonstrated that loss of function of survivin or DNA-PK results in increased cellular sensitivity to DNA-damaging agents (Meek et al, 2004; Karimi-Busheri et al., 2007). Survivin can enhance tumor cell survival upon radiation exposure by enhancement of double-strand DNA break repair (Chakravarti et al., 2004; Djuzenova et al.,2007). To investigate whether SHP plus radiation affected the expression level of phosphorylated proteins in DNA-PK and survivin, Western blot analysis was performed using antibodies raised against survivin and DNA-PK. As shown in Figure 3, SHP pretreatment strongly inhibited the DNA-PK phosphorylation and downregulated the level of survivin protein in C6 in a time-depdendent manner (Figure 3A), and difference responder in NHA cells, phosphorylation of DNA-PK was strongly increased by irradiation in a time-depdendent manner, non-significant expression of survivin in the whole course (Figure 3B ). Apoptosis in C6 and NHA Cell LinesCells apoptosis was performed by flow cytometric analysis (Table 1). Treatment of SHP combined radiation induced an increase in the sub-G0/G1 peak in C6 and NHA (P<0.01), an indicator of apoptosis cells, which caused a significant increase in the percentage of C6 cells in G0/G1-phase and a modest increase in cells in S phase. By comparison, the cell cycle distribution of the NHA cells appeared to be little affected by SHP combined radiation treatment. Apoptosis rate in C6 is significantly higher than that in NHA (P<0.01). Survivin is expressed in the G2/M phase of the cell cycle in a cycle-regulated manner, however, it looks like the decrease and increase in survivin levels in NHA and C6 cells respectively fall short of transformation through the cell cycle. Perhaps SHP presents more complicated mechanism and causes the change of cell cycle . Discussion High-grade glioma remain virtually incurable with current therapeutic regimens, despite considerable advances during the last two decades in neurosurgical techniques, radiation, and chemotherapy, treatment of malignant gliomas remains mostly palliative, median survival is about 1 year from the time of diagnosis (Arslan et al., 2006; Sathornsumetee and Rich, 2006). To enhance the effects of radiation, two major strategies have been proposed. The first approach is based on the reduction of the treatment volume, the second strategy makes use of the increase of the differential response between the tumor and normal tissue to, for example, chemotherapeutic drugs, biologic agents, and genetic or proteomic techniques (Muni et al.,2010; Bidros et al.,2010; Both,2009). However, complex molecular determinants of tumor response to radiation have limited the success of these approaches to date. Obviously, it is important to find a more effective approach which would be to develop agents that selectively sensitize these glioma cells to radiation. We previously demonstrated that Shenghe powder could induce tumor cells apoptosis and reverse multidrug resistance (Wang et al., 2007), inhibit the proliferation of C6 and decrease malignant degree (Xia et al., 2007), improve the survival time of the patients with postoperative high-grade glioma (Xia, 2005). Its complex components may offer a novel antitumor mechanism. The objective of this study was to determine the effect of Shenghe powder on the repair of DNA damage induced by radiation. Radiation causes a broad spectrum of DNA lesions such as alkali-labile lesions, single-strand breaks and DSBs. DSBs is the major threat to the genomic integrity of cells, which is biologically the most significant lesion produced by radiation and other exogenous cytotoxic agents (Lavin et al.,2005; Singh et al.,2009; Sarcar et al.,2010 ). It has been shown that histone H2AX becomes phosphorylated immediately after irradiation, which in turn is believed to recruit DNA repair factors to sites of DNA double-strand breaks (Kanaar et al., 1998; Momota et al.,2003). Because γH2AX is closely associated with DNA double-strand break repair mechanisms, we studied the radiation response of C6 and NHA lines in vitro, using γH2AX expression as a marker of DNA double-strand breaks. Of the various techniques currently available to assay double-strand breaks, we chose an immunofluorescence approach involving quantification of phosphorylation of histone H2AX because of its sensitivity, which can be readily detected by fluorescent immunostaining with antibodies to the phosphorylated form of the protein. The fluorescent intensity correlates well with the number of DNA double-strand breaks (Sedelnikova et al.,2002). This research showed that Shenghe powder enhanced radiosensitivity of C6 cells, the pre-treatment with Shenghe powder resulted in reduced numbers of γH2AX foci in irradiated C6 cells, the capacity of repair was markedly decreased. In contrast, Shenghe powder could depress the radiation-induced DNA double-strand break and enhance the DNA repair capacity in NHA. DNA-PK is overexpressed in brain glioma, it has been firmly established that DNA-PK plays an important role in DNA end joining, especially DNA double-strand break repair after radiation, prior reports have shown that inhibition or deficiency in DNA-PK leads to decreased DNA double-strand break repair and increased radiosensitivity both in vitro and in vivo (Meek et al., 2004; Veuger et al.,2003; Chaudhry et al.,2010; Burdak-Rothkamm et al.,2007). The repair of DNA double-strand breaks after radiation may account for the poor apoptotic response and ultimate radioresistance of malignant glioma cells. This study showed that Shenghe powder pretreatment strongly inhibited the DNA-PK phosphorylation, downregulated the level of DNA-PK in a time-depdendent manner in C6 cells. Nevertheless, DNA-PK was strongly increased in NHA. It suggested that Shenghe powder improved the radioresistance of NHA and enhance radiosensitivity of C6 cells which correlated with DNA-PK phosphorylation. Survivin is a novel member of the inhibitor of apoptosis (IAP) family, which is located on human chromosome 17q25.3, encoding a 16.5-kDa protein comprising amino acids. Survivin is the smallest member of the mammalian IAP family, which is highly expressed in the G2/M phase of the cell cycle, which enhances survival of tumor cells primarily through suppression of apoptotis-related cell death perhaps via direct inhibition of caspase-related proteins (Dohi et al.,2004; Shin et al.,2001). Survivin may enhance tumor cell survival upon radiation exposure such as regulation of double-strand DNA break repair and tumor cell metabolism, which were most evident in the radiation-resistant cell lines (Shirai et al.,2009; Ogura et al.,2008). Uniform, Survivin expression is associated with glioma progression from low-to high grade, which appeared to be a marker for radiation resistance, with strongest expression in radiation-resistant glioma phenotypes (Zhen et al.,2005; Jiao et al.,2004). Further, radiation appeared to increase levels of surviving (Chakravarti et al.,2004). In the presence of radiation, Survivin appears to enhance double-strand DNA breaks repair, suppressing radiation induced cell death; thereby suppressing the expression of survivin is indispensable, which plays a crucial role in tumorigenesis and progression of glioma (Saito et al.,2008; Söling et al.,2007). This study showed that SHP pretreatment strongly inhibited the expression of survivin in C6 in a time-depdendent manner. As the biological underpinnings of such phenomena as radioresistance are often multi-rather than unifaceted. the multiple factors focus on seeking multi-approach to enhance radiosensitivity of glioma, include down-regulation resistance genes or up-regulation of target genes. Shenghe powder, its complex components may offer multitarget treatment, which anastomose to requirement of multi-approach treatment in glioma. In conclusion, our studies revealed that Shenghe powder had a novel bidirectional function: (1) Shenghe powder diminished the formation of γH2AX foci in C6 cells exposed to radiation, decreased the capacity of DNA repair, enhanced radiosensitivity of C6 cells, which correlated with the changed of DNA-PK and survivin. (2) Shenghe powder increased the DNA-PK phosphorylation, improved the radioresistance of normal human astrocytes and prevented the normal astrocytes from radiation. Acknowledgments This work was supported by Health Science Foundation of Shanxi (200914). References
The following images related to this document are available:Photo images[tc11027f1.jpg] [tc11027t1.jpg] [tc11027f2.jpg] [tc11027f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}