 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
African Journal of Traditional, Complementary and Alternative Medicines, Vol. 8, No. 3, 2011, pp. 312-321 Protective Effect Of Ssanghwa-Tang Fermented By Lactobacillus fermentum against Carbon Tetrachloride-Induced Acute Hepatotoxicity In Rats Hyun-Ae Eum1, Ji-Hye Lee1, Min-Cheol Yang, Ki Shuk Shim, Jae-Hoon Lee, and Jin Yeul Ma*1,2
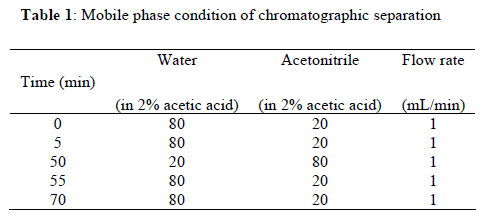
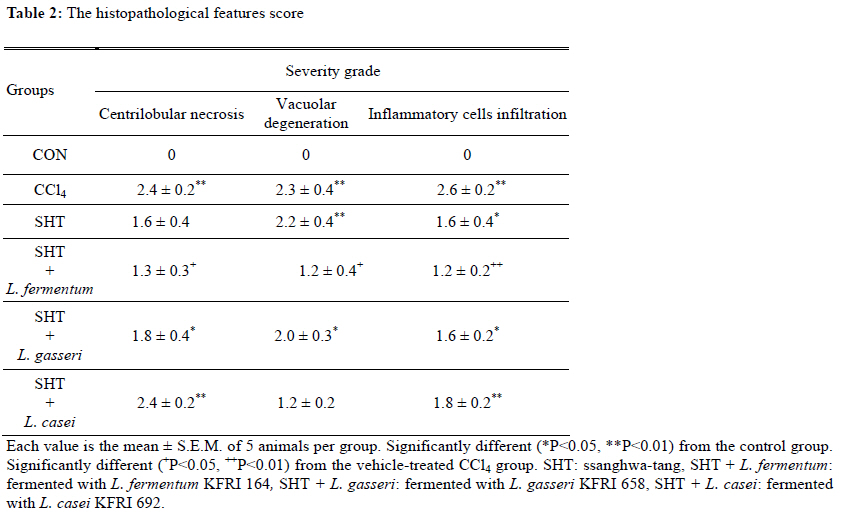
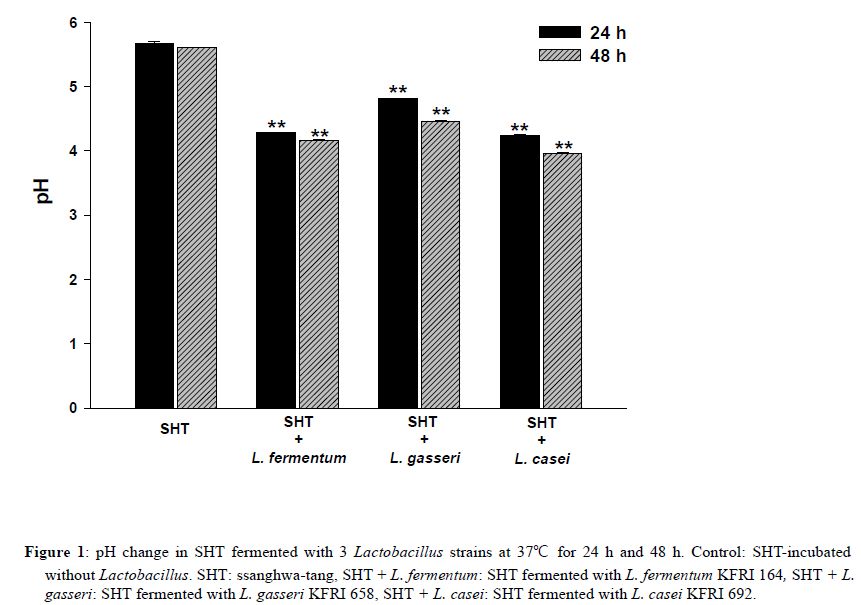
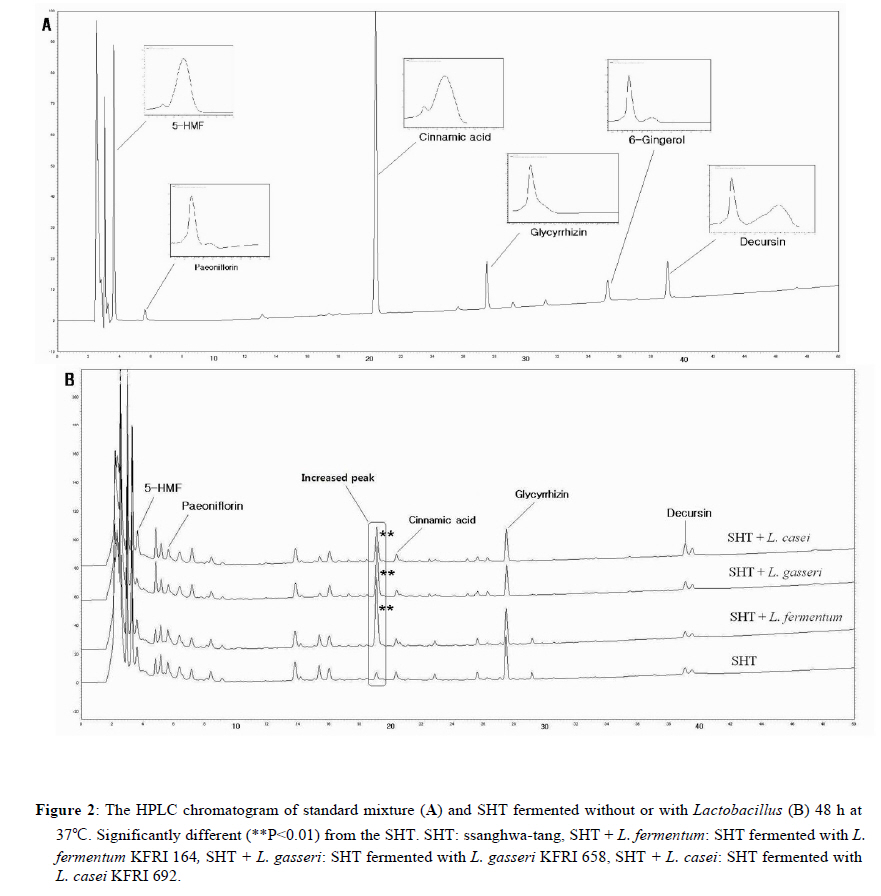
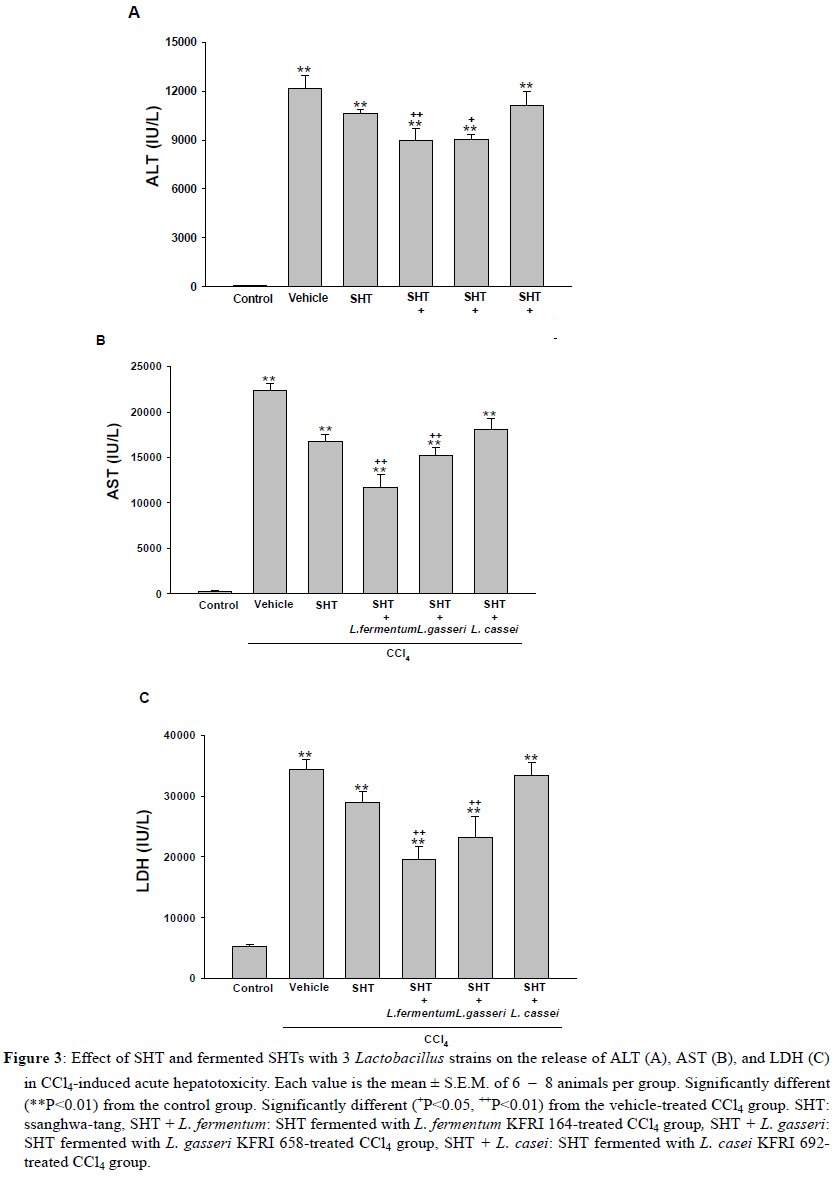
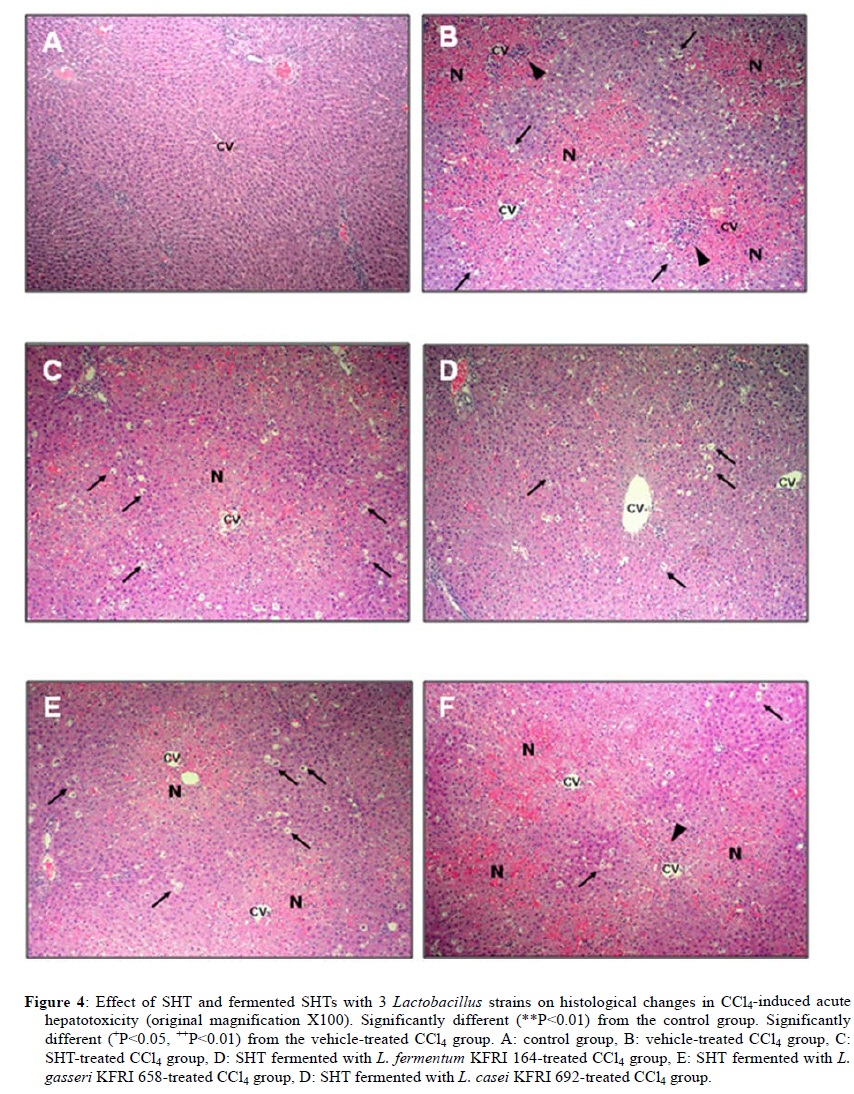
2Center for Herbal Medicine Improvement Research, Korea Institute of Oriental Medicine, 483 Exporo, Yuseong-gu, Daejeon 305-811, Korea Code Number: tc11040 Abstract Ssanghwa-tang (SHT) is a traditional herbal medicine formula that has been used for the development of physical strength, relief of pain, and the reduction of fatigue. In this study, we fermented SHT with Lactobacillus fermentum (L. fermentum), Lactobacillus gasseri (L.gasseri), or Lactobacillus casei (L.casei) to investigate the hepatoprotective effects of SHT and fermented SHT with Lactobacillus on carbon tetrachloride (CCl4)-induced liver injury in rats. Rats were given CCl4 (1 ml/kg, 50% CCl4 in olive oil) intraperitoneally and either SHT or fermented SHTs (15 ml/kg) was administered 30 min before CCl4. At 24 hr after CCl4 injection, the levels of transaminases in the serum were markedly increased. These increases were significantly attenuated by either SHT + L. fermentum or SHT+ L.gasseri. However, SHT and SHT + L.casei showed slight suppression of the increase of transaminases. The liver histological changes were diminished by treatment with SHT + L. fermentum. Additionally, the potential hepatoprotective effect of fermented-SHTs correlated with the amount of unknown metabolite which is produced during fermentation process with L. fermentum, L.gasseri, or L.casei. Therefore, these results suggest that the hepatoprotective effect of SHT may be improved by fermentation with L. fermentum and the intestinal bacterial enzyme activities may likely play an important role in the pharmacological action of herbal medicines. Key words: Carbon tetrachloride, Fermentation, Herbal fomula, Lactobacillus, Ssanghwa-tang. Abbreviations: ALT: Alanine aminotransferase; AST: Aspartate aminotransferase; LDH: Lactate dehydrogenase; Carbon tetrachloride: CCl4; L.casei: Lactobacillus casei; L. fermentum: Lactobacillus fermentum; L.gasseri: Lactobacillus gasseri; SHT: Ssanghwa-tang. Introduction Ssanghwa tang (SHT), a traditional herbal medicine formula, has been used for thousands of years as a therapeutic formula in Korea, and is still widely used. Although SHT has been widely used as a nutritional tonic in commercial medical products in Korea, not much interest has been given to SHT outside Korea (Ha and Park, 1998). SHT consists of 9 herbs: Paeoniae Radix Alba, Rehmanniae Radix Preparata, Angelica Gigantis Radix, Cnidii Rhizoma, Astragali Radix, Cinnamomi Cortex, Glycyrrhizae Radix, Zingiberis Rhizoma, and Zizyphi Fructus. It has been used for the development of physical strength, the inhibition of inflammation, relief of pain, and the reduction of fatigue. SHT has been also known to have a hepatoprotective effect (Ahn et al., 1984; Han et al., 1984). Most traditional herbal medicines are orally administrated and their components inevitably brought into contact with intestinal microflora in the alimentary tract (Park et al., 2002). The intestinal bacteria transform these components before absorption from the gastrointestinal tract (Kim et al., 1999). Therefore, the intestinal bacteria may contribute to the pharmacological activities of herbal medicines (Kobashi and Akao, 1997; Kim et al., 1998; Hasegawa et al., 1997). It has been suggested that the number of intestinal bacteria varies depending upon the personal traits of the individuals and that the pharmacological actions of herbal compounds are expressed when the herbal components are transformed to bioactive compounds by intestinal bacteria (Lee et al., 2002). According to this viewpoint, the metabolic transformation of herbal components to bioactive compounds may be a factor in the personal variation in therapeutic benefit of theses herbal components. Some studies have shown that some pharmacological activities of herbal medicines are exerted by bioactive aglycones produced upon biotransformation of active glycosides by intestinal bacteria. For example, paeoniflorin, an active gylcoside of Paeoniae Radix Alba, was not absorbed per se, but its aglycon paeoniflorgenin was absorbable and circulating in the bloodstream, thus enabling its pharmacological action (Hsiu et al., 2003). In other study, it was a metabolite of paeoniflorin that existed in high concentration in blood when paeoniflorin was orally administered (Heikal et al., 1997). Also, the bioavailability of paeoniflorin is poor and it was suggested that its metabolite is responsible for the pharmacological actions of Paeoniae Radix Alba (Takeda et al., 1995). In the case of jakyak-gamcho-tang (shaoyao-gancao-tang in Chinese or shakuyaku-kanzo-to in Japanese Kampo Medicine), its antispasmodic action originates from paeonimetabolin-I, which is the metabolite of the active glucoside, paeoniflorin, formed due to biotransformation by intestinal bacteria (He et al., 2003). Similarly, isoliquiritigenin, which is a metabolite of isoliquiritin, is a potent antispasmodic agent, accounting for the antispasmodic action of Glycyrrhizae Radix (Sato et al., 2007). These findings suggest that the intestinal bacterial enzyme activities should play an important role important role in the pharmacological action of herbal medicines and could be a factor for constitutional classification. In this study we selected L. fermentum, L. gasseri, and L. casei as a strain to fementation, based on the change in pH after inoculating into SHT. Previous study showed L. fermentum played an important role in regulating the balance of microflora and it has been one of predominant lactobacilli in human intestine and vaginal tract (Aroutcheva et al., 2001; Park et al., 2005). Many healthy effects of L. gasseri have been reported (Klaenhammer and Kullen, 1999). For example, inhibitory activity against some pathogenic and food-spoilage species, lowering of serum cholesterol concentrations, adjustments of the immune system and an enhancement of intestinal functions. L. casei has received attention as a probiotic due to evidence of antimicrobial (Ingrassia et al., 2005; Brink et al., 2006; Nemeth et al., 2006); and anti-diarrheal (Isolauri et al., 1991; Srinivasan et al., 2006). We fermented SHT with lactic acid bacteria and assessed whether the hepatoprotective effect of SHT is enhanced upon oral administration to rats with acute hepatic injury induced by carbon tetrachloride (CCl4). Materials and Methods Preparation and fermentation of SHT SHT is a mixture of 9 medicinal herbs. The herbs were purchased from the special herb market (Kyungdong oriental herb market, Seoul, Korea), and authenticated by Dr. Ok Pyo Zee, college of pharmacy, SungKyunKwan University, Korea. Their voucher specimens (S-CON, S-164, S-658, S-692) have been deposited in Korea Institute of Oriental Medicine (KIOM, Daejeon, Korea). SHT consisted of the hot water extracts from a mixture of 468.5 g of Radix of Paeonia lactiflora (Paeoniae Radix), 187.5 g of Rehmanniae Radix preparate, 187.5 g of Radix of Angelica gigas (Angelica Gigantis Radix), 187.5 g of Radix of Astragalus membranaceus (Astragali Radix), 187.5 g of Rhizoma of Cnidium officinale (Cnidii Rhizoma), 140.5 g of Cortex of Cinnamon (Cinnamomi Cortex), 140.5g of Radix of Glycyrrhiza uralensis (Glycyrrhizae Radix), 74.5 g of Rhizoma of Zingiber officinale (Zingiberis Rhizoma), and 100 g of Fructus of Zizyphus jujube (Zizyphi Fructus). The SHT water extract was obtained by boiling the total 1,674 g of the crude herb mixture in 16,740 ml of distilled water for 3 h using a COSMOS-660 extractor (Kyung-Seo Machine Co., Korea). Then the water extract was filtered through a Whatman No.1 filter paper. Biotransformation of SHT The SHT water extract was used as the culture media for fermentation after adjusting the pH to 7.0 using 1 M NaOH. Thirty of the 36 Lactobacillus strains used were acquired from the Korea Food Research Institute (KFRI, Seongnam, Korea) and the other 6 strains were acquired from the Korean Collection for Type Cultures (KCTC) at Korea Research Institute of Bioscience and Biotechnology (KRIBB, Daejeon, Korea). MRS broth (DifcoTM Lactobacilli MRS Broth, Becton Dickinson, USA) and MRS agar (BactoTM Lactobacilli MRS AGAR, Becton Dickinson, USA) were used as liquid medium for the growth of lactic acid bacteria and as slant medium for subculture, respectively. The 36 strains of Lactobacillus were subcultured a slant cultures and in liquid medium. The bacterial strains were inoculated in slant medium and incubated at 37°C for 24 h. When colonies were formed, the cultures were kept in the refrigerator after blocking oxygen with paraffin film. To retain the activated cultures and protect them from contamination, they were transferred to new slant agar medium every 2-3 weeks interval. The activated cultures were inoculated into MRS broth and incubated for 24 hr and then these served as the inoculum. For the fermentation, 500 ml of SHT in an Erlenmeyer flask with cap was inoculated with 5 ml of the inoculums as described above. This was then incubated at 37°C for a period of 48 hr. At 24 hr intervals, samples were collected and analyzed for pH. L. fermentum (KFRI 164); L. gasseri (KFRI 658); and L. casei (KFRI 692) were selected as the best candidates for animal experiments from among the 36 strains, after observing changes in pH after 24 hr and 48 hr, respectively, upon inoculating into SHT. As the control, SHT was also incubated at the same condition without inoculation. After fermentation or incubation, the SHT and fermented SHT were filtered with a 60μm nylon net filter (Millipore, MA, USA), and stored at −4 ◦C for further experiments. High Performance Liquid Chromatography Analysis In order to prepare samples for high performance liquid chromatography (HPLC) analysis, 5 ml of fermented-SHTs were taken and lyophilized. An extract of the lyophilized sample was then obtained by applying 5 ml of 70% EtOH to the samples and embedding them in the water bath held at 70° C for 8 hr. Next, the insoluble residue of the extract was eliminated after centrifugation and the supernatant was concentrated using a vacuum evaporator. Finally, the concentrate was triturated with an ultrasonic processor after applying 5 ml of MeOH, followed by filtration through a 0.45μm filter. The filtered sample was then used for HPLC analysis. All solvents (HPLC-grade water, acetonitrile, and methanol) used in this study were obtained from SK chemicals (Ulsan, Korea). The HPLC system (Hitachi Co. Japan) consisted of a pump (L-2130), autosampler (L-2200), column oven (L-2350), and diode array UV/VIS detector (L-2455). The output signal of the detector was recorded using an EZchrom Elite software for Hitachi. For separation of sample, an OptimaPak C18 column (5 μm,100Å, 4.6 mm × 250 mm, RS tech, Korea) was used and UV wavelength was 254 nm. The mobile phase was water and acetonitrile with gradient elution at a flow rate of 1.0 mL/min and the column temperature was maintained at 40℃ (Table 1). The injection volume of samples was 10μl. HPLC analysis was performed for six marker compounds, decursin of Angelica gigantis Radix, paeoniflorin of Paeoniae Radix Alba, 5-HMF of Rehmaniae Radix Preparata, cinnamic acid of Cinnamomi Cortex, glycyrrhizin of Glycyrrhizea Radix, and 6-gingerol Zingiberis Rhizoma in SHT. The conditions for operating the HPLC system are summarized in Table 1. Animals and treatment regimens Male SD rats weighing 250-270 g (Orient bio Ltd. Seoul, Korea) were fasted overnight but given tap water ad libitum. All animals were treated humanely under the Korea Institute of Oriental Medicine Care Committee Guidelines. The animals were randomized into the following treatment groups: control vehicle (control), CCl4 (1.0 ml/kg body wt, 50% CCl4 in olive oil), SHT (15 ml/kg body wt) + CCl4, SHT + L. fermentum (SHT fermented with L. fermentum KFRI 164, 15ml/kg body weight) + CCl4, SHT + L. gasseri (SHT fermented with ; L. gasseri KFRI 658, 15 ml/kg body weight) + CCl4, and SHT + L. casei (SHT fermented with L. casei KFRI 692, 15ml/kg body weight) + CCl4. CCl4 was administered by intraperitoneal injection, and SHT or fermented SHTs were administered orally at 30 min prior to the injection of CCl4. Animals were sacrificed 24 hr after CCl4 injection, and blood and liver samples were collected for further assessments. The dose of SHT and fermented SHTs was decided according to the clinical dose used in humans. The clinical treatment dosage of SHT per day for an adult was 3 times the unit intake of SHT, which corresponds to 900 ml of the SHT water extract prepared in this study. Blood biochemical analysis The serum alanine transaminase (ALT), aspartate transaminase (AST) and lactate dehydrogenase (LDH) activities were determined by standard spectrophotometric procedures using ChemiLab ALT, AST, and LDH assay kits (IVDLab Co., Ltd., Uiwang, Korea), respectively. Blood was collected from the abdominal aorta of each rat. The blood was centrifuged at 5000 rpm at 4°C for 10 min to separate the serum. Histopathological analysis Liver tissues were fixed in 10% neutral buffered formalin, embedded in paraffin, and cut serially into 5-μm sections. The hematoxylin and eosin (H&E)-stained sections were evaluated at 200× magnification using an Olympus CKX 41 microscope (OLYMPUS OPTICAL Co., Tokyo, Japan). To evaluate the degree of liver injury after CCl4 injection we created an injury grading score as 0 (unremarkable), 0.5 (minimal), 1 (mild), 2 (moderate), and 3 (marked) based on severity of necrotic lesions in the liver parenchyma (Table 2). Statistical analysis All results are presented as the mean ± S.E.M.. One-way analysis of variance (ANOVA) followed by Student– Neuman–Keuls test was used to determine the significance of the differences between the experimental groups. A P-value <0.05 was considered significant. Results Acid development of Lactobacillus strains in SHT fermentation The acid developments in SHT fermented by 3 Lactobacillus strains are shown in Figure 1. At 48 hr after incubation, the Lactobacillus strains increased acid development when compared with SHT. The greatest drop in pH occurred in L. casei. The acid development of SHT fermented with Lactobacillus strains increased rapidly until 24 hr of fermentation and slowly increased thereafter until 48 hr. However, there were no changes on pH in SHT incubated without Lactobacillus. Each value is the mean ± S.E.M. of 5 animals per group. Significantly different (*P<0.05, **P<0.01) from the control group. Significantly different (+P<0.05, ++P<0.01) from the vehicle-treated CCl4 group. SHT: ssanghwa-tang, SHT + L. fermentum: fermented with L. fermentum KFRI 164, SHT + L. gasseri: fermented with L. gasseri KFRI 658, SHT + L. casei: fermented with L. casei KFRI 692. Bioconversion of SHT constituents by Lactobacillus fermentation The HPLC results for SHT and fermented SHTs are shown in Figure 2. In comparing SHT incubated without Lactobacillus (Figure 2A), fermented SHTs (Figure 2B, C, D) showed different chromatogram profiles. All of fermented SHTs, one of the unidentified peak (RT: 19.1 min) was significantly larger than SHT incubated without Lactobacillus. When compared the areas of this peak, the increasing rates of this peak were different depending on the Lactobacillus strain (L. fermentum > L. gasseri > L. casei). This result suggests that the constituents of SHT were changed by Latobacillus fermentation and the extent of fermentation by-products are different depending on Lactobacillus strains. Serum ALT, AST, and LDH activities The effects of SHT and fermented SHTs on serum transaminases levels in CCl4-intoxicated rats were assayed and the results are shown in Figure 3. In the control group, serum levels of ALT, AST, and LDH were 46.7 ± 4.2, 306.7 ± 4.2 U/L and 5280.0 ± 348.3 U/L, respectively (Figure 3). In the CCl4-intoxicated group, the ALT, AST, and LDH levels were markedly increased to approximately 261-, 73- , and 7-fold when compared with the normal control animals, respectively. This increase was not significantly inhibited by SHT. In the CCl4-intoxicated rats with the SHT + L. fermentum or SHT + L. gasseri, however, the ALT and AST levels were significantly inhibited by 26.4% (P<0.007) and 25.8% (P<0.019), respectively, and by 47.7% (P<0.0004) and 31.8% (P<0.035), respectively. The increased serum LDH level also significantly decreased by either SHT + L. fermentum (P<0.0003) or SHT + L. gasseri (P<0.011). There was no significant difference between CCl4 alone and SHT + L. casei group. Therefore, these results show that the administration of SHT fermented with L. fermentum significantly decreased the levels of serum transaminases more effectively than the other test substances. The histology features of the rat livers were assessed by light microscopy. Table 2 summarizes the data of liver damage induced by CCl4 in pathological histology. A liver section from control rats showed normal liver lobular architecture and cell structure. In contrast to this, the liver tissues from rats treated with CCl4 showed marked histological changes including centrilobular necrosis, vacuolar degeneration neighboring the necrotic portion, and inflammatory cell infiltration. The reduction in the liver injury score after treatment with SHT, SHT + L. gasseri, or SHT + L. casei was not statistically significant. However, treatment with SHT + L. fermentum significantly ameliorated liver injury compared with treatment with CCl4 alone (Figure 4). Changes in histology were accompanied by less elevated alanine aminotransferase levels (Figure 3 A, B, C). Discussion Recently, herbal therapy has become increasingly popular due to its successful use over extended time periods in Asia and Europe. A complex range of different herbal medicines are known to be beneficial in the treatment of chronic diseases, although the mechanism by which these herbal medicines function is not fully established (Yuan and Lin, 2000). In addition, several studies have shown that the fermentation of herbal extracts with lactic acid bacteria may improve the therapeutic benefits of these compounds by converting glucoside to aglycone, which can increase systemic absorption. Lactic acid bacteria play an important role in fermentation processes and have a great influence on the quality and preservation of the end products. As they produce lactic acid, thus causing the lowering of pH. Therefore, the change in pH of the culture medium during fermentation is one of the most important fermentation parameters. We assessed changes in pH of SHT at 24 hr and 48 hr after incubation with 36 different strains of lactobacillus. After 48 hr, significant decrease in pH was seen in 3 strains: in L. casei, L. fermentum and L. gasseri, indicating good candidate as a starter strain to fermentation. In this study, therefore, we selected L. fermentum, L. gasseri, and L. casei as the best candidates for animal experiments. We found that the acid development of SHT fermented with Lactobacillus strains increased rapidly until 24 hr of fermentation and slowly increased thereafter until 48 hr. These results are not consistent with the previous result (Pyo et al., 2005), where they reported that acid development of L. plantarum and L. delbruekii sub. Lactis increased until 24 hr, and then leveled off after 24 hr of incubation in soymilk fermentation. The difference between the previous and present study might be due to the differences in the fermentation substrate and lactobacillus strains used. It was also reported that the growth of L. fermentum varies depending on the nitrogen and carbon sources of the fermentation medium (Gao et al., 2009). Next, we compared the protective effects of SHT with fermented SHTs in liver damage induced by CCl4 in rats. CCl4-induced hepatotoxicity in rats represents an adequate experimental model of cirrhosis in man and it is used for the screening of hepatoprotective drugs (Al-Shabanah et al., 2000; Pérez-Tamayo, 1983; López-Novoa and Rengel, 1977). The biotransformation of CCl4 occurs in the endoplasmic reticulum (ER) and is mediated by Cytochrome P450 (Castro et al., 1968), with CYP2E1 being the principal isoform implicated as the catalyst. Cytochrome P450 is inhibited suicidally by the reactive metabolites of CCl4 (Athar et al., 1997) with CCl3•, radical initially formed, being relatively unreactive until it reacts very rapidly with oxygen to yield a highly reactive trichloromethyl peroxy radical (CCl3OO•), which is the probable initiator of lipid peroxidation (Bhat and Madyastha 2000). The lipid peroxidation process results in the release of soluble products that may affect the cell membrane. Cell membrane integrity is thus broken and the enzymes (such as ALT, AST, etc.) in the cell plasma leak out. Thus, measurement of serum ALT and AST activities is one of the most widely used means for assessing the extent of hepatic damage. Our results show that in the CCl4 alone group there is a marked hepatocytolysis, shown especially by the highest level of ALT, AST, and LDH activity. Liver tissue isolated from the CCl4 alone group also shows destruction of the liver lobular architecture and cell structure, indicating severe hepatocellular damage. However, SHT shows a decreased release of ALT, AST, and LDH in the serum. Interestingly, the serum activities of serum transaminases were decreased by treatment of fermented SHTs, but most of all, SHT + L. fermentum shows the strongest hepatoprotective effect on the release of serum tansaminase after CCl4 injection. The hematoxylin-eosin stained liver sections also showed significantly fewer histological changes in the SHT + L. fermentum-treated CCl4 group than the vehicle-treated CCl4 group. The severity scores of SHT + L. fermentum , SHT + L. gasseri, and SHT + L. casei are sorted in the order SHT + L. fermentum < SHT + L. gasseri < SHT + L. casei in terms of centrilobular necrosis and inflammatory cell infiltration, showing a positive correlation with serum ALT, AST, and LDH content. It is well known that phytoestrogens found in medicinal herb are compounds known as isoflavones. Isoflavones are poorly absorbed in the small intestine as compared to the more bioactive aglycones. The aglycones are considered more bioactive because they have smaller molecular weight and are hydrophilic, thus they are more readily absorbed (Chang and Nair 1995; Setchell and Cassidy 1999). When isoflavones are ingested, these conjugated isoflavones undergo hydrolysis by enzymes such as β-glucosidases in the intestine. It has been shown that Lactobacillus possesses β-glucosidase, which plays a major role in the intestinal hydrolysis of numerous plant β-glucosides (Coward and others 1993; Choi and others 1999; Tsangalis and others 2004). Thus, during the fermentation process, the isoflavones in herb are enzymatically hydrolyzed by probiotic bacteria, leading to changes in the concentration of all the isoflavone forms in herb. In addition, the production of βglucosidase by the lactic acid bacteria was strain-dependent: some strains could produce this enzyme constitutively while in others the enzyme was not produced at all. In this study we found that an increased HPLC peak (RT: 19.1 min) at 48 hr after fermentation, and the amount of yield was different depending upon Lactobacillus strain. Interestingly, the potential hepatoprotective effect of fermented-SHTs correlated with an increased HPLC peak detected at 17.3 min. The height of the 19.1 min-peak developed as SHT + L. fermentum > SHT + L. gasseri > SHT + L. casei in magnitude, which can be related to the yield of the unidentified metabolite formation upon fermentation. From these results, we suggest that fermentation of SHT with Lactobacillus caused the formation of the unidentified metabolite during fermentation process, which could be shown by the hepatoprotective activity against CCl4-induced liver injury. In conclusion, our study demonstrated that the hepatoprotective effects of SHT increased by fermentation with L. fermentum as demonstrated by decreases in serum transaminases levels and histological changes compared with the CCl4 model group. This protective effect of fermented SHT may be correlated to the yield of the unidentified metabolite formation upon fermentation. However, further work is needed to isolate the metabolite and study its pharmacological actions, in order to see whether it is a bioactive metabolite expressing hepatoprotective activity. Acknowledgments This work has been supported by the grant K10050 awarded to Korea Institute of Oriental Medicine (KIOM) from Ministry of Education, Science and Technology (MEST), Korea. References
Copyright 2011 - African Journal of Traditional, Complementary and Alternative Medicines The following images related to this document are available:Photo images[tc11040f4.jpg] [tc11040f2.jpg] [tc11040f1.jpg] [tc11040t1.jpg] [tc11040t2.jpg] [tc11040f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}