 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
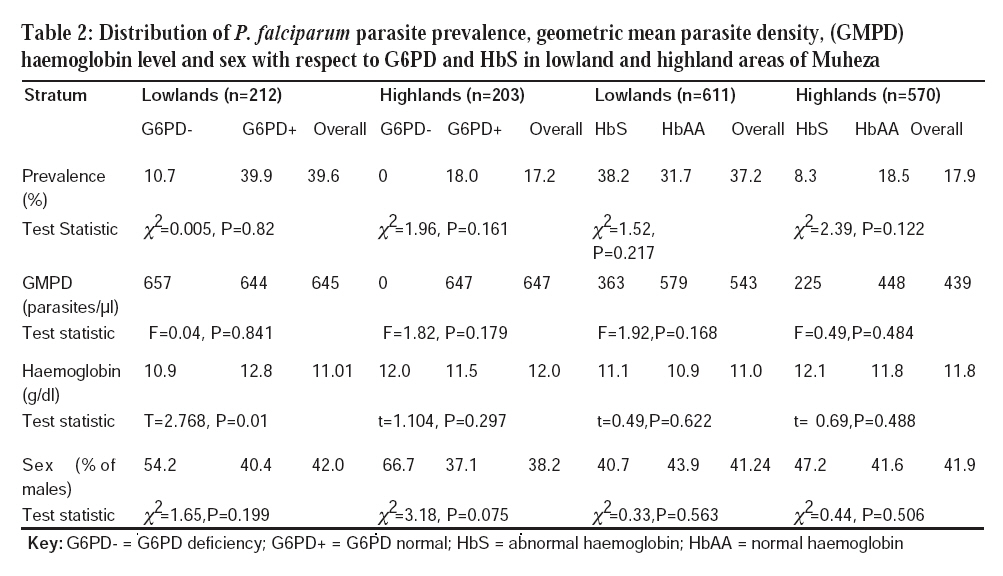
Tanzania Journal of Health Research, Vol. 10, No. 1, January, 2008, pp. 9-13 Prevalence of glucose-6-phosphate dehydrogenase deficiency and haemoglobin S in high and moderate malaria transmission areas of Muheza, north-eastern Tanzania M.D. SEGEJA1*, B.P. MMBANDO, M.L. KAMUGISHA, J.A. AKIDA, Z.X. SAVAELI, D.T. MINJA, H.A. MSANGENI and M.M. LEMNGE National Institute for Medical Research, Tanga Medical Research Centre, P.O. Box 5004, Tanga, Tanzania Received 7 August 2007 Revised 30 November 2007 Accepted 18 December 2007 Code Number: th08003 Abstract Glucose-6-phosphate dehydrogenase (G6PD) deficiency and haemoglobin S (HbS) are very common genetic disorders in sub Saharan Africa, where malaria is endemic. These genetic disorders have been associated with protection against malaria and are therefore under strong selection pressure by the disease. In November-December 2003, we conducted a cross-sectional survey to determine the prevalence of G6PD deficiency and HbS in the population and relate these to malaria infection and haemoglobin levels in lowland and highland areas of differing malaria transmission patterns of Muheza, Tanzania. Blood samples from 1959 individuals aged 6 months to 45 years were collected. A total of 415 (21%) and 1181 (60%) samples were analysed for G6PD deficiency and HbS, respectively. Malarial parasite prevalence was 17.2% (114/1959) in the highlands and 39.6% (49/1959) in the lowlands. Lowlands had higher prevalence of G6PD deficiency and HbS than highlands (G6PD deficiency = 11.32% (24/212) versus 4.43% (9/203), P = 0.01, and HbS = 16.04% (98/611) versus 6.32% (36/570), P = 0.0001). Logistic regression model showed an association between G6PD deficiency and altitude [lowlands] (Odds ratio [OR] 3.4, 95% CI=1.49; 7.90, P=0.004). In the lowlands, G6PD deficient individuals had lower mean haemoglobin (10.9g/dl) than normal ones (12.8g/dl), P = 0.01. These findings show that high malaria transmission in the lowlands might have selected for G6PD deficiency and HbS. Keywords: malaria, haemoglobin S, glucose-6-phosphate dehydrogenase, Tanzania Introduction Glucose-6-phosphate dehydrogenase (G6PD) deficiency and haemoglobin S (HbS) are very common genetic disorders in Sub-SaharanAfrica where malaria is endemic. These genetic disorders have been associated with protection against malaria morbidity (Moormann et al., 2003). Studies have shown that children with sickle cell trait (HbAS) are protected from both mild and severe malaria (Aidoo et al., 2002). HbASis also associated with reduced parasite densities during intercurrent Plasmodium falciparum infections (Williams et al., 2005) and enhanced acquired immunity, which suggests that HbAS probably protects against malaria infection due to increased parasite clearance and induction of antibodies (Cabrera et al., 2005). These two traits are therefore under strong selection pressure by the disease (Francis & Pete, 2006). G6PD deficiency is associated with many clinical disorders, such as neonatal jaundice, haemolytic anaemia and several cardiovascular diseases (Beutler, 1994). Some studies have shown that P. falciparum parasite densities are lower in G6PD deficient individuals than those with normal (Ruwende et al., 1995). The enzyme is very polymorphic in many parts of the world, existing in four most common G6PD variants, namely, G6PD A, A–, B and G6PD Mediterranean. G6PD A has a normal enzyme activity and is commonly found in Africa; G6PD A-has 8 – 20% of normal enzyme activity and is also mainly found in Africa. G6PD B has a normal enzyme activity and is distributed worldwide while G6PD Mediterranean has less than 5% of normal enzyme activity and is found in the Middle East (Mehran, 2003). Some studies in African children have shown that G6PD A- is associated with 46 – 58% reduction in risk of severe malaria for both female heterozygotes and male hemizygotes and probably homozygotes have a similar level of protection from malaria, although this genotype is quite rare (Ruwende et al., 1995). Some individuals, particularly among populations of African, Asian or those of Mediterranean origin are apt to develop haemolysis when given therapeutic doses of the antimalarials 8-aminoquinolines and sulphonamides. This defect is due to diminished activity of the enzyme glucose-6-phosphate dehydrogenase which catalyses the first and rate controlling step in the hexose monophosphate shunt metabolic pathway (Luzzatto et al., 2001). Despite that, very few studies on these two genetic disorders have been conducted in north-eastern Tanzania. Therefore, we conducted this study to determine the prevalence of G6PD deficiency and HbS in the population and to relate these to malaria infection and haemoglobin levels. Results from this study will provide baseline information on G6PD deficiency and HbS in an area with difference transmission intensity which will be useful in designing and implementing different malaria interventions. Materials and methods Study area and population Muheza district is topographically divided into highland and lowland strata. Malaria is holoendemic in the lowlands with prevalence of over 75% (Lyimo, et al., 1991). In the highlands, there is a modest varied level of malaria transmission whereby in some areas prevalence is 33% (Ellman et al., 1998). In both strata, malaria transmission is highest during and following long rainy season, which usually extends from March through July. Ethnically, the population of Muheza comprises of the Wasambaa in the highlands and Wabondei and Wadigo in the lowlands. The study involved four villages (Kizerui, Kwelumbizi, Shambangeda and IBC Msasa) situated within the East Usambara Mountains in the highlands (600–1000 m) and four villages (Moa, Kwangena, Mwandusi and Jirihini) in the lowlands (0– 300 m). Both the highlands and lowlands communities practice subsistence farming of maize, cassava, and banana. Production of coconuts and fishing is common among some communities in the lowland villages. Study design and sample collection This was a cross-sectional study which was conducted during the short rains in November-December 2003.A random sample of 1959 individuals aged 6 months to 45 years was selected for the survey and sub-samples were later chosen for laboratory analysis. Capillary blood samples of about 300 -500μl were collected into EDTA anticoagulant tubes. Due to limitations of laboratory reagents, a random sample of 21% (415/1959) and 60% (1181/1959) were analyzed for G6PD deficiency and HbS, respectively. Thick and thin blood smears for malaria parasite diagnosis were prepared. Identification and quantification of Giemsa stained blood slides for malaria parasites were done using a light microscope. Haemoglobin levels were estimated using Haemocue machine. Determination of G6PD deficiency was done using fluorescent screening test (Dacie & Lewis, 1995). Briefly, 10 μl of EDTA anticoagulated blood was added to 50 μl of the reagent mixture and then 10 μl of the reaction mixture (D-glucose-6-phosphate, NADP, Saponin, oxidized glutathione, Tris aminomethane, HCl, MgCl2, MTT, PMS, Azide) was applied immediately onto Whatman No. 3 filter paper at the beginning of the reaction and after 5 and 10 minutes. Control samples of known G6PD deficient and normal were run together with the tested samples. The spots were examined under Ultra Violet (UV) light. The G6PD normal samples fluoresced while deficient ones did not fluoresce. Haemoglobin S was determined using sickling test (Dacie & Lewis, 1995). Briefly, 5 μl of freshly prepared reagent was added to 1 μl of EDTA anticoagulated blood on a slide and then sealed with cover glass using petroleum jelly. Control samples of known HbS and HbA were also included with test samples and then kept overnight in a moistened chamber. Slides were examined using light microscope. Sickling took place in HbS blood samples but not on HbA blood samples. Data analysis All data were entered and verified in Epi Info 6.04d while statistical analysis was done using STATA software version 8 (STATA Corp., College station, TX). Prevalence of G6PD and HbAS in bivariate variables was compared using χ2or Fisher’s exact tests. Geometric mean density (GMI) of malaria parasites and Hb concentration in G6PD deficiency and HbAS groups were compared using T and F tests. Multivariate logistic regression model was used to estimate risk factors associated with the prevalence of G6PD deficiency. Results Malarial parasite prevalence was 17.2% in the highlands and 39.6% in the lowlands. Among individuals examined for G6PD deficiency, the Wasambaa was the dominant ethnic group accounting for 76.9% (n = 156) in highlands while in the lowlands Wadigo accounted for 49.5% (n = 105) of the population followed by Wasegeju (20.3%; n = 43) and Wamakonde (15.6%; n=33). In the lowlands, the Wasambaa contributed to 3.3% (n = 7) only. The prevalence of G6PD deficiency was significantly higher in lowland compared to highland areas (G6PD = 11.32% vs. 4.43%, P= 0.01) (Table 1). Similarly, the prevalence of HbS was significantly higher in lowland compared to highland areas (16.04% vs. 6.32%, P< 0.001) (Table 1). Individuals with HbS had lower level of parasites density than normal ones in both strata, although the difference was not statistically significant (Table 2). There was no significant relationship between HbS and Hb levels and sex (Table 2). Among results showed that in highlands, sex (male) was more associated with G6PD deficiency although the difference was not significant (Table 2). Discussion Like many other studies in Muheza, Tanzania, malaria parasite prevalence was higher among individuals living in the lowlands than highlands (Kamugisha, 2005; Maxwell et al., 2003). The prevalence of G6PD deficiency and HbS was significantly higher in lowland individuals sampled for G6PD analysis (Table 2), P. falciparum parasite prevalence was lower in G6PD deficient than in normal G6PD individuals in both strata. Parasite density was lower in G6PD deficient individuals in the highland. In the lowlands, haemo globin levels were significantly higher in individuals with normal G6PD (Table 2) compared with those with G6PD deficiency (12.8 vs. 10.9g/dl; P=0.01). Results from a logistic regression model fitted for strata, sex, Hb levels and parasite prevalence, showed that the significant variable associated with G6PD deficiency was strata [lowlands](Odds ratio [OR] 3.4, 95% CI=1.49; 7.90, P=0.004). Sex (female) had reduced effect on the G6PD deficiency and Hb levels. However, the difference was not statistically significant (OR=0.49, P=0.069 and OR=1.19, P=0.081) for sex and Hb, respectively. A logistic regression model was also fitted for the two strata separately where compared to highland areas. Individuals with HbS had lower level of parasites density than normal ones in both strata, although the difference was not statistically significant. In both the highlands and lowlands villages, P. falciparum parasite prevalence was lower in G6PD deficient than in normal G6PD individuals. Parasite density was lower in G6PD deficient individuals in the highland. The G6PD deficiency is known to provide partial protection against malaria, by providing defective environment in the affected red cells. In the lowlands, haemoglobin levels were significantly higher in individuals with normal G6PD. A previous study (Awah & Uzoegwu, 2006) showed that less severe clinical malaria symptoms were observed more in G6PD deficient HbAS when compared to G6PD normal HbAA subjects during malaria attack. Therefore inheriting both genetic disorders reduces malarial anaemia, parasitaemia and severe malarial symptoms. Like in our study, a recent study in Kenya (Moorman et al., 2003) observed higher prevalence of HbS and G6PD deficiency in the lowlands compared to the highlands.Another study in north-eastern Tanzania, using a more robust screening technique (Enevold et al., 2007) found prevalence of HbS of around 16.0% in Mgome village in the lowlands of Muheza district. Using the rapid screening test, we observed a similar prevalence rate of HbS in lowland Muheza. The observation of a higher prevalence of HbS and G6PD deficiency genes in the lowlands is likely to be directly linked to the high malaria transmission. The reduced parasites density seen in individuals with HbS in both lowland and highland areas suggest that malaria has already exerted a strong selective pressure in the human populations at both sites. In conclusion, G6PD deficiency and HbS genes may provide a natural protection against malaria. Acknowledgements This study was supported by a grant from the Ministry of Health and Social Welfare through the National Institute for Medical Research (NIMR), Tanzania. We thank all individuals who agreed to participate in the study and the village leadership for their cooperation. Dr Deus Ishengoma is thanked for useful discussion and editing of the manuscript. We would like to thank Dr. Andrew Kitua, Director General NIMR, for permission to publish the manuscript. References
© Copyright 2008 - Health User's Trust Fund (HRUTF) The following images related to this document are available:Photo images[th08003t2.jpg] [th08003t1.jpg] |
| |||||||||

{kind=link}
{kind=link}