 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
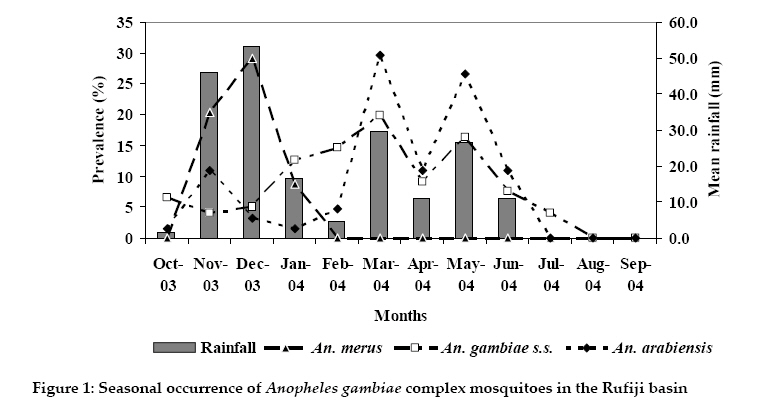
Tanzania Journal of Health Research, Vol. 12, No. 1, January, 2010 Diversity, spatial and temporal abundance of Anopheles gambiae complex in the Rufiji River basin, south-eastern Tanzania E.S.P. KIGADYE1*, G. NKWENGULILA2, S.M. MAGESA3 and S. ABDULLA4 1Faculty of Science Technology and
Environmental Studies, The Open University of Tanzania. P.O. Box 23409, Dar
es Salaam, Tanzania * Correspondence: Dr. Emmanuel S.P. Kigadye; E-mail: ekigadye@uccmail.co.tz Received 27 October
2009 Code Number: th10008 Abstract The Anopheles gambiae complex contains the most efficient malaria vectors in the world. Identification of the species and the concomitant distribution are vital for effective malaria control. The objective of the study was to establish the diversity, spatial and seasonal abundance of malaria vectors in the Rufiji River Basin in Tanzania and the implications on malaria vector control. Mosquitoes were collected using CDC light-traps. Anopheles mosquitoes were identified by conventional keys. Anopheles gambiae complex were speciated using standard PCR method. Of the 562 specimens analysed by PCR 69% produced fragments equivalent to An. gambiae s.s. (390 bp), 23% equivalent to An. arabiensis (315bp), and 7% as An. merus (464 bp). An. gambiae s.s. and An. merus were more abundant on the plateau than on the flood plain (Fisher’s exact test, P< 0001), whereas An. arabiensis was equally abundant between the two sites (Fisher’s exact test, P=1656). The density of the three sibling species of An. gambiae complex also varied with the seasons. An. gambiae s.s. and An. arabiensis were most predominant species during the start of the rainy season, but as season progresses, An. gambiae s.s. predominated. An. merus was only recorded during the short rainy season. In conclusion, both An. gambiae s.s., An. arabiensis and An.merus are malaria vectors in the Rufiji River basin and that An. merus is recorded for the first time in the south-eastern coast of Tanzania. These findings are important in the planning and implementation of malaria vector control activities in the Rufiji River basin, south-eastern Tanzania. Key words: An. gambiae s.s, An. arabiensis, An. merus, abundance, distribution, Tanzania Introduction Studies of malaria transmission in Tanzania are complicated by the number of vector species involved. Of the 45 species of anopheline mosquitoes recorded in the country, 11 have been implicated as malaria vectors (Gillies & De Meillon, 1968). The three species which, are most commonly involved in malaria transmission in Tanzania, are Anopheles gambiae s.s. Giles, An. arabiensis Patton and An. funestus Giles (Mnzava & Kilama, 1986). Another member of the An. gambiae complex, An. merus Diönitz has been shown to be a minor vector (Mosha & Petrarca, 1983; Temu et al., 1998). In addition, An. rivulorum Leason, a member of the An. funestus group and An. marshalii have also been implicated in malaria transmission in north-eastern Tanzania (Magesa et al., 1991; Wilkes et al., 1996; Malima, 2000). As in most areas of Africa, Anopheles gambiae complex is the major vector species in Tanzania, with the sibling species An. gambiae s.s. and An. arabiensis Paton being the most important within the complex (White, 1974). An. gambiae s.s. predominates or is the only species in humid coastal and lacustrine areas, while An. arabiensis is predominant in or exclusive in dry and semi-arid areas (Mnzava & Kilama, 1986). The occurrence of An. gambiae s.s. is restricted to 128-240 km along the coastal belt of Tanzania. However, in a study in the Kilombero Valley, the species was also found about 500km from the coast (Mboera, 2000). Despite several decades of malaria research, locality-specific information on the diversity, spatial and temporal distribution of Anopheles gambiae complex, is still wanting. Species within the complex differ in host biting preference, abundance and vector competence, identification of the mosquito vectors to species level and mapping species distribution in heterogeneous environments is critical (Coetzee, 2004). Moreover, strategies for malaria control require a solid understanding of vector dynamics and factors influencing their spatial and temporal distribution (Mbogo et al., 2003). Such information would help to develop early warning systems for predicting malaria epidemics, and planning control programmes based on accurate prediction of their effects (Thompson et al., 1997). Given the need for locally adapted, integrated vector control strategies, a better understanding of the ecology, biology and population structure of the vectorial units is necessary (TDR, 2002). The objective of the study was therefore; to establish the identity of the Anopheles gambiae complex present in the Rufiji basin, determine their abundance, spatial and seasonal dynamics and their implications to vector control. Materials and Methods Study site and mosquito sampling Mosquitoes were collected from six wards in the Rufiji District (7.47°8’S, 38.62°39.17’E), located 50 km south east of Dar es Salaam in Tanzania. Mosquito sampling was confined to an area under the Rufiji Demographic Surveillance System (RDSS) that covers six contiguous wards with 31 villages (about 60 x 30 km wide). The district lies below 500 metres above sea level and receives an annual rainfall of 800-1000mm. According to the perceptions of local people, and data on malaria incidence from health care facilities, malaria is the most important public health problem for both adult and children in Rufiji District (Goodman, 2002; Mwageni et al., 2002). Monthly mosquito catches were carried out in each of the selected households for two consecutive nights for over a period of one year. Mosquito were caught using standard Centre for Disease Control light traps (CDC, Atlanta, GA) hung from the ceiling at the foot end of the bed occupied by a person sleeping un-treated net (Mboera et al, 1998). Light traps were operated between 18.00-0600 hours. An. gambiae complex were separated from other anophelines and culicines using the keys by Gillies & Coetzee (1987), counted and then stored in labelled eppendorf tubes with desiccant (silica gel) for laboratory processing at Ifakara Health Institute laboratory. Differentiation of the sibling species of the An. gambiae complex The PCR method of Scott et al. (1993) was used with minor modifications. One leg from each mosquito was placed in a separate 1.0 mL eppendorf tube to which 12.5µL of PCR master mix was added. This was centrifuged for one minute at 1600 rev/minute in a micro-centrifuge. All quantities given by Scott et al. (1993) were halved. The PCR procedure included an initial cycle of denaturation at 94°C for 2 minutes followed by 30 cycles of denaturation at 94°C for 30 seconds, and a final extra extension step at 72°C for 8 minutes using a HybaidTM thermal cycler. The resulting amplified Deoxyribonucleic Acid (DNA) was run on an ethidium bromide stained 2.5% agarose gel and photographed under ultraviolet light illumination as described by Scott et al. (1993). Data analysis Analysis was performed using SPSS® software. Fishers exact test was used to test for difference in mosquito abundance between the plateau and flood plain areas. Regression analysis was carried out to test for significant difference between rainfall and mosquito abundance. Ethical considerations The study received ethical approval from the Medical Research Coordination Committee, Institute for Medical Research, Tanzania (Certificate No. NIMR/HQ/R.Sa/Vol. IX/192). Informed consent was obtained from Local Health Authorities and Village Committees and house occupants. Results Of the total 14134 An. gambiae complex females collected between October 2003 and September 2004, 562 specimens were analysed by PCR in order to differentiate the sibling species and the results were as follows: An. gambiae s.s. was the most abundant (69%) followed by An. arabiensis (24%) and the least abundant was An. merus (7%). An. gambiae s.s. and An. merus were more abundant on the plateau than on the flood plain (P<0001), that of An. arabiensis was similar between the two areas (P=1656). The density of An. gambiae s.s. was generally high during the long rains between March and May 2004 and was virtually undetectable in August-September 2004, a relatively dry period. As for An. gambiae s.s. the density of An. arabiensis reached a peak during the long rains in March-May and dropped to zero during the dry season, July-October 2004. The density of both An. gambiae s.s. and An. arabiensis positively correlated with rainfall but not significantly (P>0.05). The density of An. merus was considerably lower than that of the other two species. The species was only detected between October and January 2004 (Figure 1). Discussion Three members of the An. gambiae complex namely An. gambiae s.s. Giles, An. arabiensis Patton and An. merus Diönitz were recorded during the present study. This is the first report on the speciation of An. gambiae complex in the Rufiji River basin in south-eastern Tanzania. In addition, the occurrence of An. merus is reported for the first time in the south-eastern coast of Tanzania. Previous studies have reported An. merus from the north-eastern coast (White, 1972; Bushrod, 1981; Mosha & Petrarca, 1983; Temu et al., 1998). The presence of An. merus in Rufiji River basin is somehow surprising as the species is typically known to breed in brackish water associated with mangroves which do not occur in the area. An. merus has been recorded inland as far as Segera in north eastern Tanzania (Bushrod, 1981), more than 100 km from the coast. However, the distribution range of An. merus is still not clearly known in Tanzania. The propensity for inland distribution is well documented in southern Africa, where the species is believed to breed in fresh water (Coetzee et al., 1993). The abundance of An. gambiae s.s. and An. merus on the plateau than the flood plain observed in the present study could probably be attributed to the variation in environmental and ecological factors such as rainfall and breeding habitats (Kigadye, 2006). One of the limitations of the present study is the absence of data on larval ecology, which could have otherwise, revealed the breeding ecology of the three members of An. gambiae complex in the Rufiji basin. The results of the present study showed that the population densities of the three sibling species vary with the seasons. These results confirm the findings of previous studies on the population dynamics of the An. gambiae complex mosquitoes (Charlwood et al., 2000). Typically, at the start of the long or during the short rains, the number of An. gambiae s.s. and An. arabiensis undergo explosive growth. As the rains progresses, An. gambiae s.s. becomes predominant while at the end of the rains the density of both species declines. In a study in north-eastern Tanzania, An. gambiae s.s. was found to predominate over An. arabiensis during the main rains and throughout the cool dry season, after which An. arabiensis multiplied in excess of An. gambiae s.s. (White et al., 1972). Similar to our findings, studies elsewhere in the dry savannah of Africa (Smith et al., 1995) have shown that the numbers of mosquitoes drop to very low levels. Three sibling species of the An. gambiae complex; An. gambiae s.s., An. arabiensis and An. merus occur in sympatry in the study area. It is therefore, important to find their specific breeding habitats as they might have some divergence in niche or other part of their life history that can be exploited for control. Conclusively, the study contributes to the understanding of the ecology and distribution of malaria vectors with respect to species composition, spatial and seasonal heterogeneities that could guide vector control operations in the Rufiji basin. Acknowledgements The authors wish to thank the Open University of Tanzania, the National Institute for Medical Research and the Ifakara Health Institute for financial and material support. Prof. Maureen Coetzee of the National Institute for Communicable Diseases, Johannesburg, South Africa is thanked for technical support. References
Copyright 2010 - Tanzania Journal of Health Research The following images related to this document are available:Photo images[th10008f1.jpg] |
| |||||||||

{kind=link}