 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
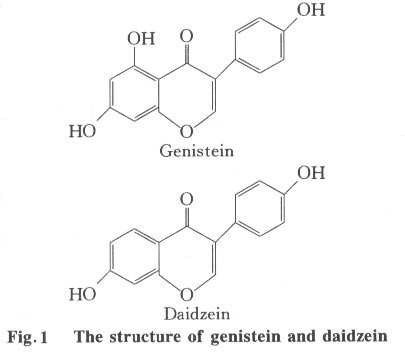
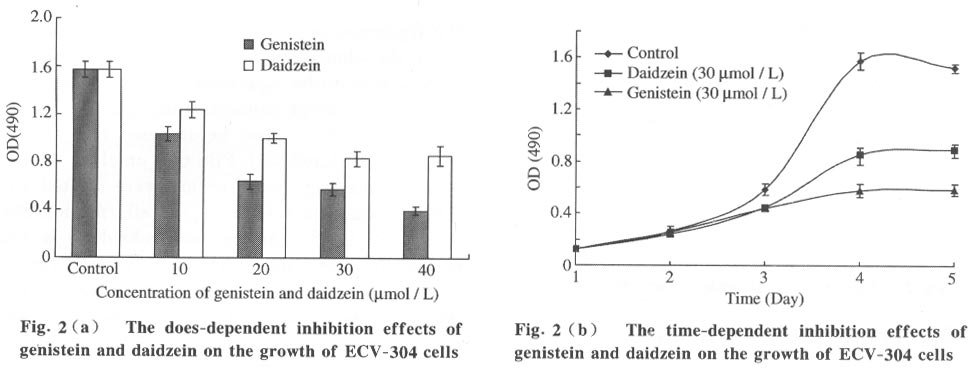
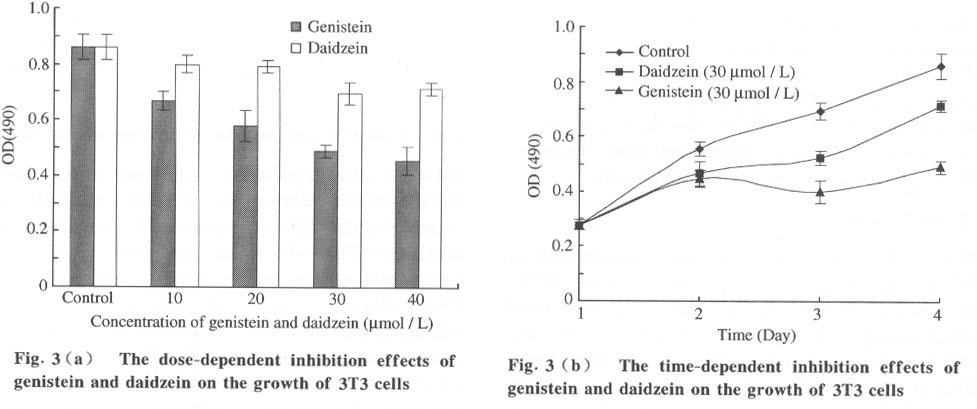
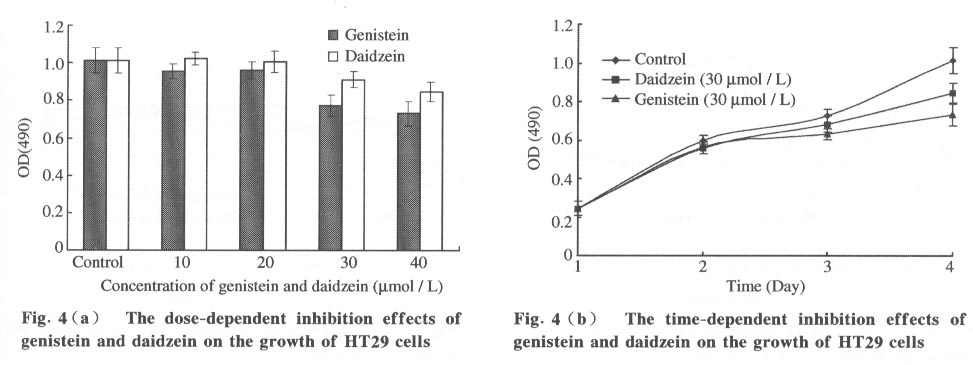
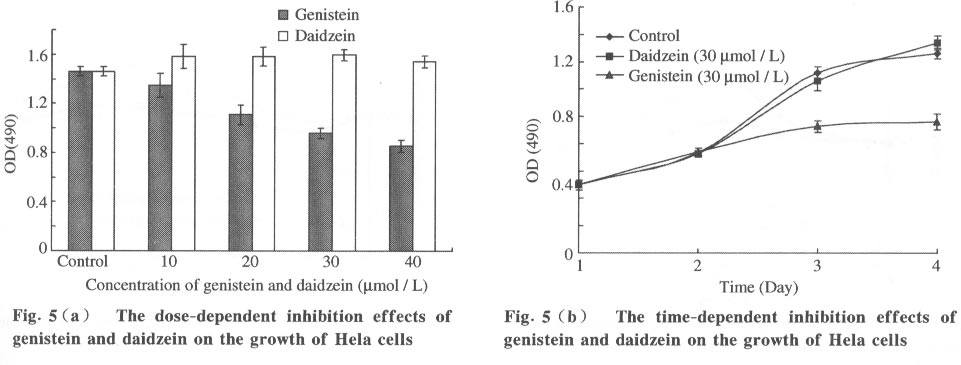
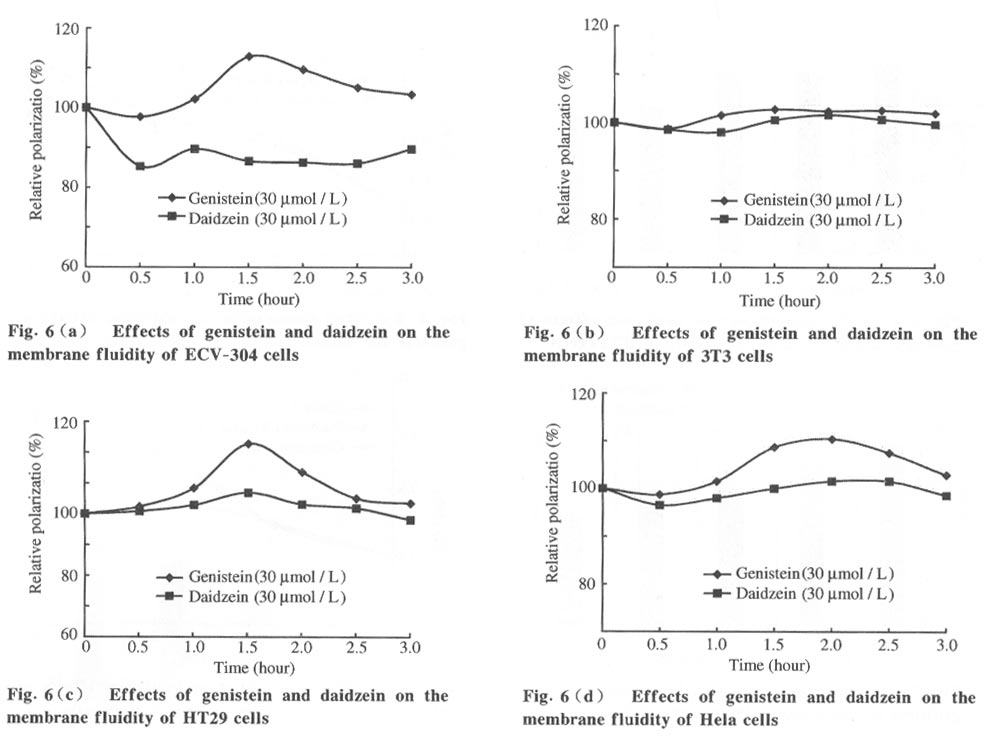
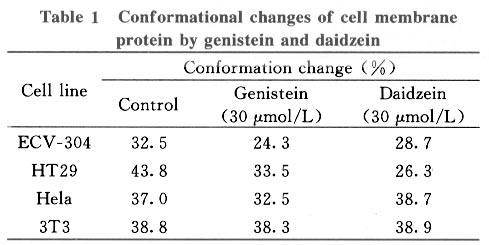
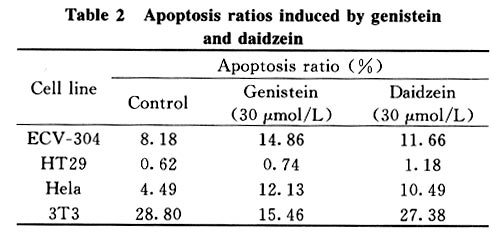
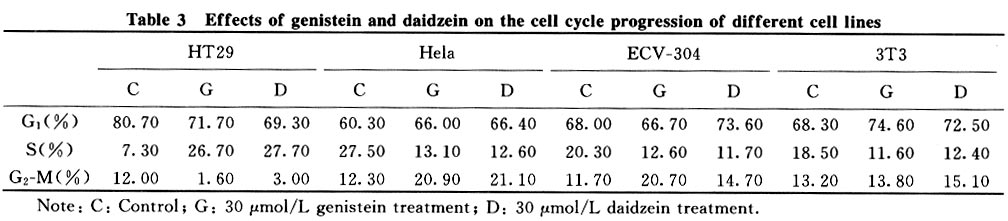
Tsinghua Science and Technology, December 2001, 6(5), pp. 410-416 Genistein and Daidzein Effects on Proliferation, Cell Membranes, Cell Cycles and Cell Apoptosis of Different Cell Lines* LI Chonghua Center for Ocean Science and Engineering, Department of Biological Sciences and Biotechnology, Tsinghua University, Beijing 100084, China * Supported by the Foundation of Combined Research Projects of Tsinghua University,
Hong Kong Baptist University, and Beijing University of Chinese Medicine on
Chinese Traditional Medicine. Received: 2001-09-18 Code Number: ts01089 Abstract: Genistein and daidzein are two principle isoflavonoids in soybeans. They have received increasing attention in the past few years because of their possible roles in cancer prevention. Here are provided experimental evidences that genistein could inhibit the growth of human ladder carcinoma cells (ECV-304), human colon cancer cells (HT29), human uterus cervix cancer cells (Hela), and murine transformed muscle cells (3T3). Different from genistein, daidzein could only inhibit the growth of ECV-304, HT29, and 3T3 cells. To elucidate the mechanisms of the anti-tumor effect of genistein and daidzein, fluorescent polarization, circular dichroism, and flow cytometric analysis were employed to study the influence of genistein and daidzein on membrane fluidity and membrane protein conformation of these cell lines. The results showed that genistein increased the order of membrane protein conformation and reduced the membrane fluidity of ECV-304, HT29, and Hela cells. Daidzein also increased the order of membrane protein conformation of ECV-304 and HT29, but had no effect on the membrane fluidity of all these four cell lines. Also demonstrated was that both compounds affected the apoptosis and cell cycle progression of some cell lines. However, the effects of genistein and daidzein were not the same. These evidences suggested that the effects of genistein and daidzein on malignant cells were multisites and multiapproaches, and there were differences between their functional mechanisms. The amelioration effect on cell conditions may represent one of the mechanisms of the effect of genistein and daidzein on the growth, differentiation, and transference of malignant cells. Key words: isoflavonoid; genistein; daidzein; anti-tumor Introduction Isoflavonoids are found mainly in soybeans, a primary food source for humans and herbivores. The structure and function of isoflavonoids are similar to 17b-estradiol (17b-E2), and have dual effects as both mammal oestrogenicity and anti-oestrogenicity, so they are also termed isoflavonic phytoestrogens (IPE). It has been reported that these compounds can reduce blood fat, as well as the risk of cancer and osteoporosis[1]. Genistein and daidzein are two principle isoflavonoids in soybeans. In recent years, they have received increasing attention because of their possible roles in cancer prevention. The structure of daidzein is almost identical to that of genistein, except for lacking one 5-hydroxyl group which results in their different hydrophobicity (Fig. 1). The hydrogen bond formed by this 5-hydroxyl group with the 4-ketonic oxygen makes genistein more hydrophobic than daidzein. The difference in their hydrophobicity may lead to their difference in functions and functional mechanisms. Previous studies demonstrated that genistein and/or daidzein has inhibition effects on the growth of leukemia, breast and prostate cancer[2-6]. Earlier studies also showed that genistein and/or daidzein inhibited the growth of human colon tumor (HCT) cells[7]. However, the functional mechanisms of genistein and daidzein remain uncertain. To investigate the effect of genistein and daidzein on the growth of malignant cells, this study used four other cell lines: human bladder carcinoma cells (ECV-304), human colon cancer cells (HT29), human uterus cervix cancer cells (Hela), and murine transformed muscle cells (3T3). This work was an attempt to determine the functional mechanisms of genistein and daidzein, by studying the effects of genistein and daidzein on the membrane characteristics, cell cycles, and cell apoptosis of these cell lines. Materials and Methods 1.1 Chemicals Genistein and 1,6-diphenyl-1,3,5-hexatriene (DPH) were purchased from Sigma Chemical (St.Louis , MO). Daidzein was purified by Dr. Zhang Rongqing. Media DMEM, RPMI1640 and trypsin were purchased from GIBCO (Grand Island, NY). Fetal bovine serum was purchased from Hyclone. All other chemicals were of reagent grade. 1.2 Cell culture HT29 cells and Hela cells were maintained in DMEM containing 5% fetal bovine serum. ECV-304 was cultured in DMEM containing 10% fetal bovine serum. 3T3 was in PRMI1640 containing 10% fetal bovine serum. All the cell lines were cultured in a 37 °C humidified atmosphere of 95% air/5% CO2. 1.3 In vitro growth inhibition assay Cells in logarithmic growing periods were plated into 96-well flat-bottom microplates (Costar Corp.) at a density of 103 - 104/well. Genistein and daidzein were added separately at various concentrations to each sample when the cells began growing exponentially. (Equal amounts of DMSO, instead of genistein and daidzein were added to the controls). After incubation for several days, 20 mL 3-(4,5-dimethy-2-thiazolyl)-2,5-diphenyl tetraz- olium bromide (MTT, 5 mg/mL) was added to each well. After incubation for another 4 h, the supernatants were removed and the formazan crystals were dissolved by adding 200 mL DMSO. The plate was then read on a microplate reader (Bio-Rad, model 550) at 490 nm. An MTT colorimetric assay was also conducted after treatments with 30 mmol/L genistein and 30 mmol/L daidzein for 0, 24, 48, 72, and 96 h. 1.4 Membrane fluidity assay The fluorescence polarization method was used to detect the change in membrane fluidity. A DPH fluorescence probe was used. It was dissolved in THF to the storage concentration of 2 mmol/L and held at 4 °C . Each time before use, this solution was freshly diluted with PBS to 2 mmol/L. Cells in the logarithmic growing period were diluted with PBS to a concentration of 2x106 cells/mL and then 2 mmol/L. DPH solution was added to a final concentration of 1 mL DPH/mL. The solution was held at 37 °C for half an hour and then washed with PBS to remove any free DPH. The cells were resuspended with PBS, then genistein and daidzein were added separately at various concentrations to each sample. Samples were incubated at 37 °C and the polarization of the solution was measured every half hour using a Hitachi-850. The wavelengths of excitation and emission were 360 nm and 403 nm, respectively. 1.5 Assays of cell membrane protein conformation Cells in the logarithmic growing period were diluted to the density of 2x106 cells/mL and treated with different doses of genistein and daidzein. The cells were incubated at 37 °C for 2 h , and then washed by PBS to remove DMSO. The changes in cell membrane protein conformation were investigated using a spectrometer (model J-500C CD) and a quartz colorimetric cup with 1-mm light distance at a time constant of 2 seconds. Autoscan was repeated 4 times and the results were collected and analyzed by computer. 1.6 Assays of cell cycle and cell apoptosis Genistein and daidzein were added to cells that had been incubated for 24 h. After being incubated for another 4 days, the cells were washed with PBS and diluted with PBS to the density of 106 cells/mL. RNaseA was added to the solution at the level of 3000 units/mL. The solution was held at 37 °C for 30 min, then filtered with a 30 mm nylon mesh. The sample was stained by PI before being measured by flow cytometry. 2 Results 2.1 Inhibition effect of genistein and daidzein on the growth of different cell lines 2.1.1 Growth inhibition of ECV-304 cells Both genistein and daidzein inhibited the proliferation of ECV-304 cells in a dose-dependent manner (Fig.2(a)). A significant inhibition effect was also observed with the increase of treatment time ( Fig.2(b) ). The median effective concentrations (EC50) of genistein and daidzein were 10 mmol/L and 15 mmol/L, and the suppression efficiencies were 63.4% and 45.6%, respectively. 2.1.2 Growth inhibition of 3T3 cells Figures 3(a) and 3(b) show that both genistein and daidzein inhibited the proliferation of 3T3 cells in a dose- and time-dependent manner. The EC50 of genistein and daidzein were 10 mmol/L and 15 mmol/L, and the suppression efficiencies were 43.1% and 17.0%, respectively. 2.1.3 Growth inhibition of HT29 cells As shown in Fig. 4, both isoflavonoids exhibited no inhibition on the growth of HT29 at low concentrations. However, they displayed stronger inhibition effects at higher concentrations. The EC50 of genistein and daidzein were 25 mmol/L and 30 mmol/L, and the suppression efficiencies were 27.7% and 16.5%, respectively. 2.1.4 Growth inhibition of Hela cells The dose-dependent effect and time-dependent effect of genistein and daidzein on Hela cells are shown in Fig.5. Genistein had an obvious inhibition effect on Hela cells; the suppression efficiency was 34.3% and the EC50 was 20 mmol/L. But daidzein had no obvious influence on the growth of Hela cells. 2.2 Effects of genistein and daidzein on membrane fluidity Membrane fluidity was evaluated by measuring the microviscosity of the cell membrane with a fluorescence spectrophotometer. High microviscosity (represented by polarization of light) indicated low membrane fluidity. The experimental data showed different effects of genistein and daidzein on different cell lines (Fig. 6). When the ECV-304, Hela and HT29 cells (except for 3T3 cells) were incubated with genistein (30 mmol/L), the microviscosity of the cell membrane increased with time in the beginning and decreased gradually after about 1.5 - 2.0 h. However, no obvious effect on the four kinds of cells was observed with a 30 mol/L daidzein treatment. 2.3 Effects of genistein and daidzein on membrane protein conformation With genistein and daidzein treatment, the coil proportion of the ECV-304 cell and the HT29 cell membrane protein decreased. The effect of genistein was stronger than that of daidzein. The coil proportion of Hela cell membrane protein also decreased with the treatment of genistein, but daidzein did not show such effects. Also, both compounds had no obvious effects on 3T3 cell membrane protein (Table 1). 2.4 Effects of genistein and daidzein on cell apoptosis Both genistein and daidzein enhanced the apoptosis ratios of ECV-304 cells, HT29 cells and Hela cells, but had no obvious effects on 3T3 cells. Genistein had a stronger effect on cell apoptosis of ECV-304 and HT29 than daidzein (Table 2). 2.5 Effects of genistein and daidzein on cell cycle progression The cell cycles of the four cell lines under genistein and daidzein treatment were investigated using FCM (Flow Cytometry). As shown in Table 3, ECV-304 cells were accumulated in the G2-M phase with the treatment of genistein and accumulated in the G1 phase with the treatment of daidzein. HT29 cells were accumulated in the S phase or G2-M phase under the effects of genistein and daidzein, respectively. As to the Hela and 3T3 cells, the number of cells in the S phase decreased while the number of cells in the G1 phase and G2-M phase 3 Discussion Genistein and daidzein, two isoflavonoids in soybeans, have been proposed as contributing significantly to the anti-tumor effect of soybeans. Previous studies have demonstrated that genistein and/or daidzein could inhibit the growth of human colon tumor (HCT) cells[7]. Here evidence is provided that genistein could inhibit the proliferation of ECV-304, HT29, 3T3 and Hela cell lines as well (EC50 was 10 mmol/L , 25 mmol/L , 20 mmol/L, and 10 mmol/L, respectively). Daidzein could inhibit the proliferation of ECV-304, HT29, and 3T3 cell lines (EC50 was 15 mmol/L, 30 mmol/L and 15 mmol/L, respectively), but had no effect of growth inhibition on Hela cells. The evidence implies potential uses of genistein and daidzein in treating cancer. Of all the cell lines, the results with the ECV-304 cell line were most attractive, because this kind of cell line is a vascular endothelia cell. How to inhibit angiogenesis during the growth of a tumor is critical to curing cancer. Here it was found that genistein and daidzein could inhibit the proliferation of ECV-304, thus they are potential anti-angiogenesis medicines. Further- more, the fact that the growth of HT29 can be inhibited by both of these isoflavonoids is another new discovery of this experiment. Although the anti-proliferation effect of genistein and daidzein on malignant cells has been extensively studied, the mechanisms of their anti-tumor function remain uncertain. Several possible mechanisms have been proposed for the chemopreventive effect of genistein, including genistein functioning as an estrogen receptor agonist/antagonist, causing DNA strand breaks, altering cell cycle distribution, inducing differentiation and/or oxidation events, inhibiting the activities of PTK and/or topoisomerase II[8-13]. Recent studies have demonstrated that daidzein at a high dose is able to enhance multiple immunologic functions in mice[14]. Also, genistein and/or daidzein affects the membrane characteristics of HCT cells, including membrane fluidity, density of cell surface charge, and the conformation of membrane proteins[7]. Fluidity of the biomembrane plays an important role in cell physiology and biochemical activity. Generally, the membrane fluidity of tumor cells is much greater than that of normal cells, so it has been used to determine whether cells have taken pathologic changes. Genistein can significantly reduce the membrane fluidity of ECV-304, HT29, and Hela cells, which means that genistein can improve the conditions of the cell membranes. On the other hand, daidzein has few or even no effects on the membrane fluidity of these four cell lines. The difference between genistein and daidzein was reflected in their different mechanisms of function. Daidzein is less hydrophobic than genistein and may react only with surrounding proteins and carbohydrate side chains, so its effect on membrane fluidity was limited. Since genistein is more hydrophobic than daidzein, its molecule could penetrate the cell membrane and change the fluidity of the membrane. It is known that genistein and daidzein have no effect on the membrane fluidity of normal cells. The fact that genistein and daidzein had no effect on the membrane fluidity of 3T3 is possibly due to that 3T3 is a transformed cell line and thus maintained many characteristics of normal cells. The proportion of random coiling of cell membrane proteins represents the order property of membrane proteins. It was demonstrated that membrane proteins of tumor cells have more random coils than those of normal cells. Genistein treatment drastically reduced the proportion of random coiling in membrane proteins of ECV-304, HT29, and Hela cells, while daidzein treatment only improved the status of ECV-304 and HT29 cells. These results provide evidence that enistein and daidzein have amelioration effects on cell conditions. The reason for both compounds having no effect on 3T3 cells might be that the content of random coils in 3T3 cells is more like that of normal cells. The changes that occurred in 3T3 cells in the process of plasmid transformation were too small to significantly change the characteristics of the cell membrane proteins. Both genistein and daidzein affected the apoptosis ratio and cell cycle progression of certain malignant cell lines. However, they had different effects on different cell lines. These evidences suggest that there were different functional mechanisms in different cell lines. The reason for the differences requires more detailed study. In summary, this study demonstrated that genistein and/or daidzein had multiple effects on four malignant cell lines. More detailed studies suggested that they functioned by changing the fluidity of the membrane - the conformation of membrane proteins - as well as by cell cycle arrest and by increasing the apoptosis ratio of malignant cells. References
Copyright 2001 - Tsinghua Science and Technology The following images related to this document are available:Photo images[ts01089t1.jpg] [ts01089f4.jpg] [ts01089f3.jpg] [ts01089f5.jpg] [ts01089f6.jpg] [ts01089f2.jpg] [ts01089t2.jpg] [ts01089f1.jpg] [ts01089t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}