 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tsinghua Science and Technology, December 2001, 6(5), pp. 421-425 Enhancement of Aminoacylase Activity by Sodium Citrate YU Fanli Department of Biological Sciences and Biotechnology, Tsinghua University, Beijing
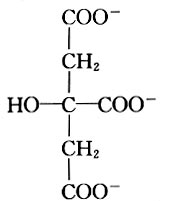
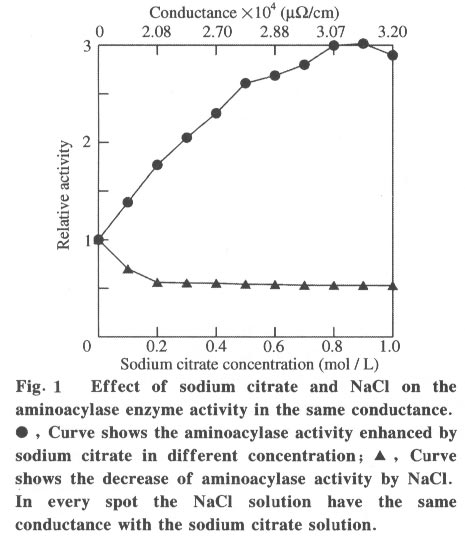
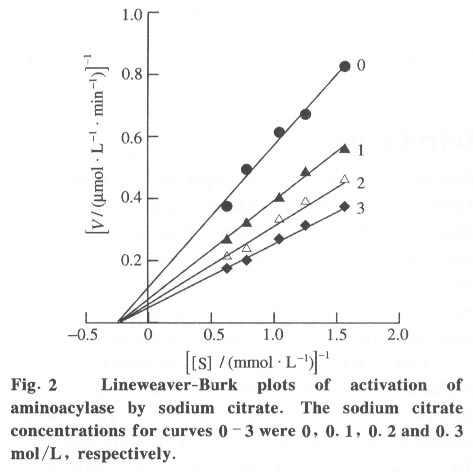
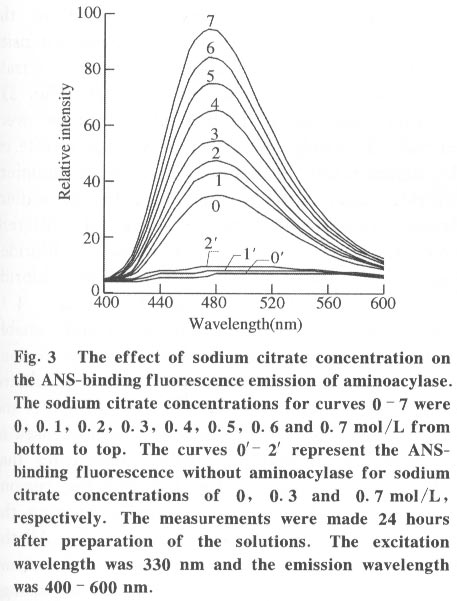
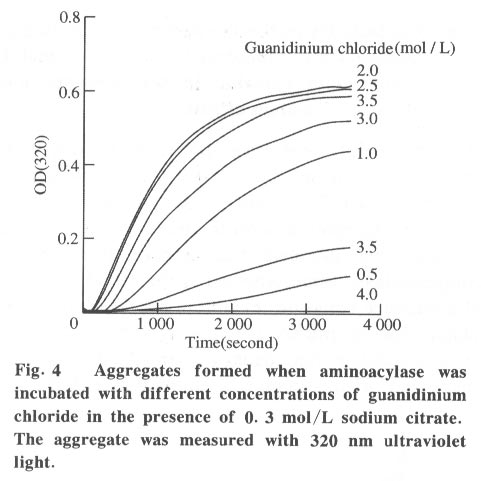
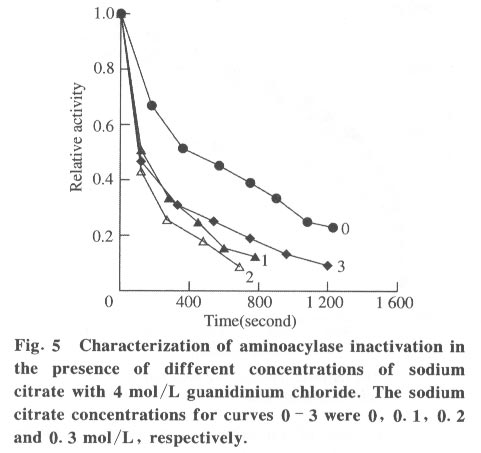
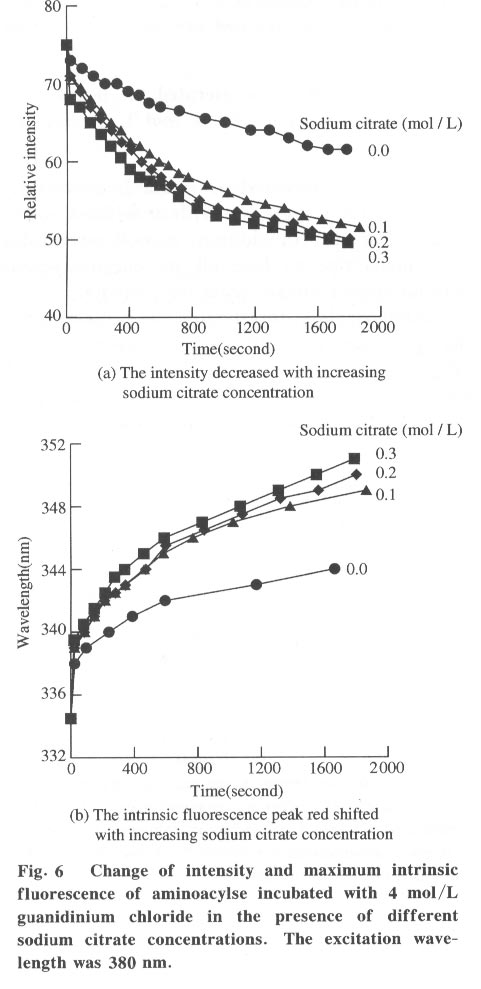
100084, China Received: 2000-12-28 Code Number: ts01091 Abstract: Kidney and other tissues of animals and humans have a high concentration of citrate which is an important intermediate substance in the citrate cycle. Citrate may play an important physiological role in metabolism. In this paper, we studied the interaction of the sodium salt of citrate with aminoacylase which is an important enzyme in metabolism and found sodium citrate can enhance the activity of aminoacylase. The maximum enzyme activity induced by sodium citrate increased approximately 3 folds over the enzyme activity without sodium citrate. The initial reaction rates for different concentrations of sodium citrate were obtained, showing that sodium citrate is a non-competitive activator. The result of the ANS binding fluorescence measurements for aminoacylase indicated that increasing sodium citrate concentrations markedly increased the ANS binding fluorescence with a blue shift of the emission spectra peak. This suggests the formation of more hydrophobic regions. Aggregates formed quickly when aminoacylase was incubated with sodium citrate (0.3 mol/L) and guanidinium chloride (0-3.5 mol/L). Aminoacylase lost enzyme activity in the guanidinium chloride more quickly in the presence of sodium citrate than in the absence of sodium citrate. The intrinsic fluorescence emission intensity decreased more quickly and the red shift of the emission spectra peak was larger than that without sodium citrate Key words: minoacylase; sodium citrate; activation; denaturation; unfolding Introduction Enzyme activation is a common phenomenon in biological studies. Many experiments have suggested that enzyme activation is caused by the conformational change of the enzyme active sites[1-4]. Other papers have showed that the enhancement of enzyme activity is associated with secondary or tertiary structural changes[5]. This study analyses aminoacylase (N-acylamino acid amidohydrolase, EC 3.5.1.14), a dimeric protein, consisting of two identical subunits each with an active site. The zinc ion at the active site of aminoacylase is essential for enzyme activity[6]. The unfolding courses of aminoacylse in guanidinium chloride, urea and SDS solutions as well as thermal unfolding have been studied[7-9]. It was also reported that Mn2+ could activate aminoacylase[10]. Kidney and other tissues of animals and humans have a high concentration of citrate and citrate is an important intermediate substance in the citrate cycle. This paper presents enzyme activity enhancement for aminoacylase as well as the conformational changes in the presence of sodium citrate. 1 Materials and Methods Aminoacylase was prepared from pig kidney as in Birnbaum's procedure[11] up to the step of acetone fractionation. The crude preparation was then purified first by gel filtration through Sephadex G-150 then by ion-exchange with DEAE-cellulose as described by Kordel and Schneider[12]. The final preparation was homogeneous on polyacrylamide gel electrophoresis (PAGE) in the presence and absence of sodium dodecanesulphonate (SDS). N-chloroacetyl-L-leucinea and ANS (1-anilino-8-naphthalenesulfonate)(1/2 magnesium salt) were purchased from Sigma. Other chemicals were local products of analytical grade. Enzyme concentration was determined by measuring absorbance at 280 nm and using the absorption coefficient A1% 1cm = 13.5 [12]. Enzyme activity was determined at 37 °C by measuring the absorbance change at 240 nm accompanying the hydrolysis of the substrate and using the molar absorption coefficient e238 =185(mol/L)-1 cm-1 as reported by Kordel and Schneider[6]. The fluorescence emission spectra were measured with a Hitachi-850 spectrofluorimeter with an excitation wavelength of 280 nm. All measured emission spectra were corrected spectra. ANS was added to the protein solutions with different concentrations of sodium citrate to study the interaction with ANS with the measurements made at 20 °C after incubation with sodium citrate for 24 h. The excitation wavelength was 380 nm. All measurements were carried out in 0.1 mol/L Tris-HCl buffer (pH 7.5). 2 Results 2.1 Aminoacylase enhanced enzyme activity by sodium citrate Sodium citrate (0 - 1.0 mol/L) enhanced the activity of aminoacylase (Fig. 1). At 0.9 mol/L sodium citrate, the enzyme activity reached its maximum value of about 3 times that without sodium citrate. When the sodium citrate concentration was higher than 0.9 mol/L , the aminoacylase enzyme activity decreased with increasing sodium citrate concentration. The effect of NaCl on aminoacylase enzyme activity was measured to determine whether the effect of sodium citrate was due to the increased ionic strength or the increased Na+ concentration (Fig. 1). The results showed that the aminoacylase activity enhancement was not due to the ionic strength. 2.2 Type of activation in aminoacylase by sodium citrate The type of activation of aminoacylase by sodium citrate was monitored by measuring the initial reaction rates (V) in different concentrations of the substrate, while the sodium citrate concentration held at different constant values. The plots of 1/V vs. 1/[S]for different concentrations of sodium citrate (Fig. 2) suggest that sodium citrate is a noncompetitive activator. Increasing sodium citrate concentrations resulted in a family of lines with different slopes and intercepts on the ordinate. These lines have a common intercept on 1/[S]axis, indicating that the Michaelis-Menten constant remained unchanged in the presence of different concentrations of sodium citrate, while the value of VM increased with the increasing sodium citrate concentration. 2.3 ANS binding fluorescence with different concentrations of sodium citrate The fluorescence emission of ANS increases when the dye binds to hydrophobic regions of a protein[13]. The fluorescence emission spectra for ANS banded by aminoacylase were measured in the presence of different sodium citrate concentrations while the aminoacylase concentration was held constant (Fig. 3). The data show that the ANS binding fluorescence emission maximum of ANS shifted from 480 nm to 474 nm with increasing sodium citrate concentrations, indicating the transfer of ANS molecules to a hydrophobic environment[14]. At the same time, the fluorescence intensity increased markedly, suggesting that more hydrophobic regions were formed. Control experiments showed that sodium citrate had little effect on the fluorescence of ANS for the sodium citrate concentration range used in this investigation (0-0.7 mol/L). 2.4 Sodium citrate accelerated aggregation of aminoacylase Aminoacylase did not aggregate in guanidinium chloride solution without sodium citrate after several hours (data not shown). However, the aggregate quickly formed when aminoacylase was incubated with guanidinium chloride (0-3.5 mol/L) in the presence of different sodium citrate concentrations (Fig. 4). The aggregation depends on the guanidinium chloride concentration with maximum aggregation reached at 2.0 mol/L guanidinium chloride. When the guanidinium chloride concentration was higher than 4 mol/L, aggregation did not occur. 2.5 Sodium citrate accelerated the denaturation of aminoacylase by 4 mol/L guanidinium chloride Aminoacylase incubated in 4 mol/L guanidinium chloride did not aggregate with or without sodium citrate (Fig. 4). In addition, it took several days for aminoacylse to lose all its enzyme activity without sodium citrate (data not shown). Aminoacylase inactivation was measured in the presence and absence of sodium citrate (Fig. 5) and the results suggested that the aminoacylase activity decreased more quickly in the presence of sodium citrate in the solution. 2.6 Intrinsic fluorescence spectra of unfolding of aminoacylase incubated with 4 mol/L guanidinium chloride in the presence and absence of sodium citrate The intrinsic fluorescence spectra of aminoacylase were measured in the presence and absence of sodium citrate in 4 mol/L guanidinium chloride solution. Increasing sodium citrate concentrations reduced the intrinsic fluorescence intensity of aminoacylase. Simultaneously, the red shift of the fluorescence emission maximum became larger and changed more quickly (Fig. 6(a) and 6(b)). The results indicate that denaturation of aminoacylase incubated with 4 mol/L guanidinium chloride in the presence of sodium citrate was faster and more serious than without sodium citrate. 3 Discussion This study shows that the enhancement of aminoacylase activity by sodium citrate was not due to changes of the ionic strength (Fig. 1). At the same time, no significant change of the intrinsic fluorescence emission of aminoacylase was found, which indicates that the aminoacylase conformation incubated with sodium citrate did not change. However, when ANS was added to the solution, the ANS-binding fluorescence intensity increased with increasing sodium citrate concentration, together with a blue shift (Fig. 3), showing that some hydrophobic regions were formed. To study the effect of sodium citrate on the denaturation of aminoacylase in guanidinium chloride, aminoacylase was added to the sodium citrate solution in the presence of different concentrations of guanidinium chloride. Aggregates formed for guanidinium chloride concentrations of 0 - 3.5 mol/L (Fig. 4), suggesting that sodium citrate could enable aminoacylase to generate more hydrophobic regions to form aggregates. Moreover, when incubated with 4 mol/L guanidinium chloride (Figs.5 and 6), the denaturation of aminoacylase increased in the presence of sodium citrate, indicating that sodium citrate exposed the hydrophobic regions and destroyed the tertiary structure causing the enzyme activity to decrease more quickly. All the data suggest that the enhancement of enzyme activity was accompanied by conformational change of aminoacylase, and the conformational change is the cause of the enhanced activity. The results show that aminoacylase was partly unfolded in the presence of sodium citrate and the conformation became more flexible with increased sodium citrate concentrations. More- over, Figs. 4, 5, and 6 show that aminoacylase incubated with sodium citrate was not in a stable state compared with the enzyme in the natural state, and it was more susceptible to the denaturant. The enzyme activity reached its maximum at a sodium citrate concentration of 0.9 mol/L, but the emission intensity and the maximum ANS-binding fluorescence of aminoacylse with different concentrations of sodium citrate continued to change, suggesting that the flexibility of the enzyme is a function of the enzyme activity, and there is an optimal flexibility for the enzyme activity. Citrate/sodium citrate is a useful buffer. However in this experiment sodium citrate enhanced the activity of aminoacylase accompanied with conformational change of the enzyme. The citrate structure is:
Since sodium citrate has three COO- , it was surmised that these three COO- could destroy the hydrogen-bond in aminoacylase and the - OH could form new hydrogen-bonds with aminoacylase replacing the hydrogen-bonds inside the enzyme, slightly disturbing the conformation which makes active sites of aminoacylase more flexible. Measurements of the activity of other enzymes would provide further data to support this theory. Other enzymes in sodium citrate solution may also be more flexible than in their natural state, so citrate/sodium citrate is used as the enzyme buffer, the effect of sodium citrate on the conformation of the enzyme should be considered. References
Copyright 2001 - Tsinghua Science and Technology The following images related to this document are available:Photo images[ts01091f1.jpg] [ts01091f4.jpg] [ts01091f3.jpg] [ts01091f2.jpg] [ts01091f5.jpg] [ts01091f6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}