 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tsinghua Science and Technology, December 2001, 6(5), pp. 426-431 Activation of Calf Intestinal Alkaline Phosphatase by Trifluoroethanol* CAO Zhifang 1. Department of Biological Sciences and Biotechnology, Tsinghua University,
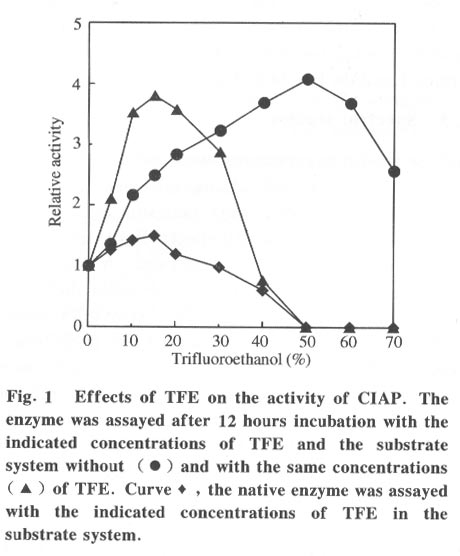
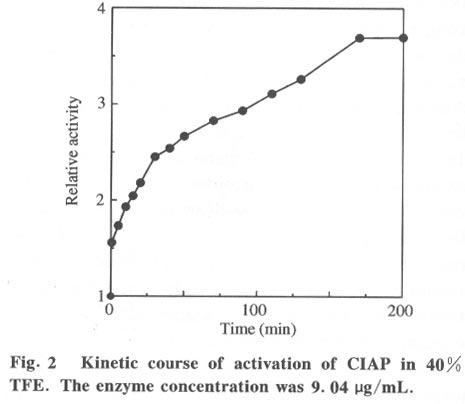
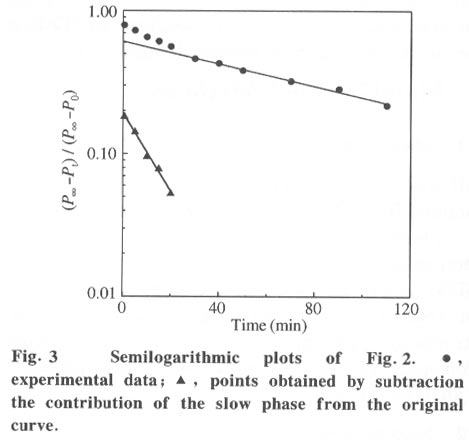
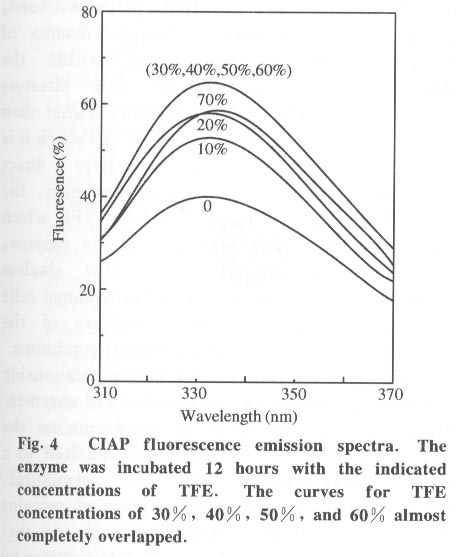
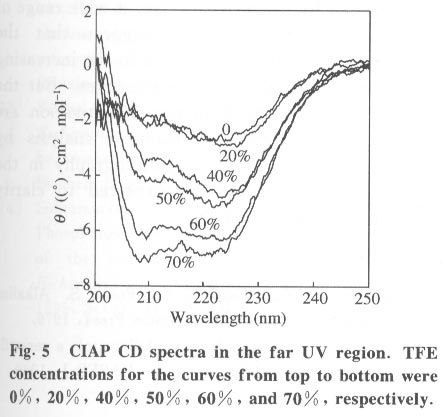
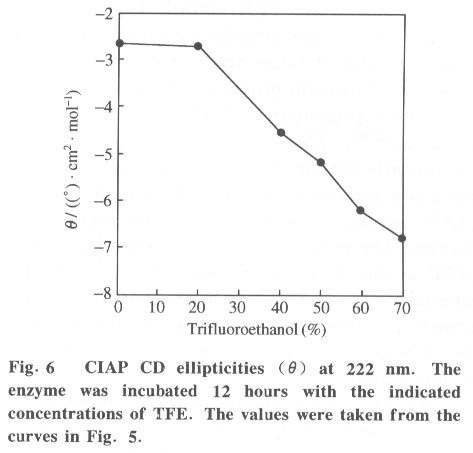
Beijing 100084, China; * Supported by the National Key Basic Research Specific Foundation of China No. G1999075607 Received: 2001-02-13; revised: 2001-05-26 Code Number: ts01092 Abstract: Alkaline phosphatase is a stable enzyme which is strongly resistant to urea, guanidine hydrochloride, acid pH, and heat. But there have been few studies on the effect of organic cosolvents on the activity and structure of alkaline phosphatase. The activity of calf intestinal alkaline phosphatase (CIAP) is markedly increased when incubated in solutions with elevated trifluoroethanol (TFE) concentrations. The activation is a time dependent course. There is a very fast phase in the activation kinetics in the mixing dead time (30 s) using convential methods. Further activation after the very fast phase follows biphasic kinetics. The structural basis of the activation has been monitored by intrinsic fluorescence and far ultraviolet circular dichroism. TFE (0-60%) did not lead to any significant change in the intrinsic fluorescence emission maximum, indicating no significant change in the tertiary structure of CIAP. But TFE did significantly change the secondary structure of CIAP, especially increasing a-helix content. We conclude that the activation of CIAP is due to its secondary structural change. The time for the secondary structure change induced by TFE preceds that of the activity increase. These results suggest that a rapid conformational change of CIAP induced by TFE results in the enhancement of CIAP activity, followed by further increase of this activity because of the further slightly slower rearrangements of the activated conformation. It is concluded that the higher catalytic activity of CIAP can be attained with various secondary structures. Key words: alkaline phosphatase; trifluoroethanol; activation; conformation Introduction Alkaline phosphatase (EC3.1.3.1) is a nonspecific phosphomonoesterase that is found in both procaryotes and eucaryotes as a dimer with identical subunits[1]. It belongs to the group of enzymes attached to the lipid bilayer of membranes by a glycosylphosphatidylinositol anchor[2 - 4]. It is a metalloprotein that has two Zn2+ ions and one Mg2+ ion in each active site. Both its catalytic mechanism and structure have been thoroughly studied and reviewed[5,6]. In mammals, alkaline phosphatase is a particularly ubiquitous enzyme that is very important in physiological functions and medical diagnosis[1,7]. The deduced amino acid sequences for alkaline phosphatase from both mammalian and bacterial sources are aligned with the sequence of the enzyme[8]. The high resolution X-ray crystal structure of E.coli alkaline phosphatase is available[8] and many site-directed mutagenesis studies to replace several of the amino acid side chains have been employed to understand the roles and the functional importance of the amino acid residues at the active site region of alkaline phosphatase[9-12]. The unfolding and refolding of alkaline phosphatase have also been well described. Alkaline phosphatase is a stable enzyme which is strongly resistant to urea, guanidine hydro- chloride, acid pH, and heat[13-16]. The complete inactivation of alkaline phosphatase in 6 mol/L guanidine hydrochloride needs about one hour. The addition of organic cosolvents to an aqueous solution of an enzyme generally decreases the enzyme activity and unfolds or aggregates the protein[17]. But there have been few studies on the effect of organic cosolvents on the activity and structure of alkaline phosphatase. These considerations led us to study the change of activity and structure of alkaline phosphatase in the organic cosolvent systems. This study demonstrates that alkaline phosphatase activity is greatly increased by trifluoroethanol (TFE), an organic cosolvent, and the activation of alkaline phosphatase by TFE is due to the secondary structure change. 1 Materials and Methods 1.1 Materials Calf intestinal alkaline phosphatase (CIAP) was obtained from Promega. The enzyme was found to be homogenous by polyacrylamide gel electrophoresis and sodium dodecanesulphonate (SDS) polyacrylamide gel electrophoresis and was not treated further. TFE was from Sigma. p-nitrophenyl phosphate (p-NPP) was from Promega. All other chemicals were local products of analytical grade. 1.2 Enzyme assays Enzyme concentration was determined using the absorption coefficient A1% 278 of 7.60[18]. The enzyme (0.724 mg/mL) was diluted 100 fold in the incubation buffer containing TFE of different concentrations and the mixture incubated at 25 °C for 12 hours, a time long enough to achieve complete change of the parameters being followed, and then assayed for enzymatic activity. Enzyme activity was determined at 30 °C by following the increasing absorbance at 405 nm, as p-nitrophenyl phosphate is hydrolyzed to p-nitrophenol using a molar absorption coefficient of 1.73x104 (mol/L) -1 · cm-1 [19]. The reaction system contained p-nitrophenyl phosphate, 4 mmol/L Na2CO3 /NaHCO3 buffer, 0.1 mol/L (pH 10.0).The extent of activity change was expressed as a percentage of the native activity. Note that all activity values were normalized to the activity of the native protein at the same concentration. The carryover TFE in the incubation buffer had no effect on the activity of the native enzyme. The progress-of-substrate-reaction method previously described by Tsou[19] was used to study the effect of TFE on the CIAP activity in the substrate system. In this method, enzyme was added to 1.0 mL of reaction mixture containing 4 mmol/L substrate in 0.1 mol/L Na2CO3 /NaHCO3 buffer (pH 10.0) containing different concen- trations of TFE. The steady-state activity was recorded when the kinetic course of the substrate reaction approached a straight line for each TFE concentration. The reaction was carried out at a constant temperature of 30 °C . Absorption and kinetics measurements were recorded on a Perkin-Elmer Lambda Bio UV/VIS spectrometer. 1.3 Spectral studies All spectral measurements were taken at a protein concentration of 212.4 mg/mL and at 25 °C . Changes in the fluorescence emission spectra were measured on a Hitachi 850 spectrofluorometer with an excitation wavelength of 295 nm. A Jasco 500C spectropolarimeter was used for circular dichroism (CD) measurements in the far ultraviolet region from 200 nm to 250 nm in a 0.2 cm path length cuvette. Mean residue ellipticities (q) were calculated on the basis of a mean residue relative molecular mass of 115. The time course of the secondary structural change of CIAP in TFE solution was monitored by measuring changes in ellipticity at 222 nm. Complete and rapid mixing was accomplished by vortex mixing. All the spectral measurements were corrected for the spectra of the buffer solution in the absence of the protein. 2 Results 2.1 Effect of TFE on the CIAP activity The CIAP activity change as a function of TFE concentration is shown in Fig. 1. The remaining activity of the enzyme incubated for 12 hours at 25 °C increased greatly in the presence of TFE and reached a maximum at 50% TFE ( 4.0-fold enhancement compared to 0% TFE). Even in 70% TFE, the enzyme is still considerably more active than the native enzyme. The study of the effect of TFE on the CIAP activity in the substrate system showed that with increasing TFE concentrations, CIAP is first activated in the 0 - 15% TFE region and reaches maximum activity at 15% TFE (about 1.5 fold), and is then gradually inactivated with a further increase in TFE concentration. It is completely inactivated at TFE concentrations higher than 50%. Interestingly, when the enzyme incubated in the buffer containing different TFE concentrations for 12 hours at 25 °C was added to the substrate reaction system containing the same concentrations of TFE, the enzyme is further activated in the 0-15% TFE region. The activated enzyme activity then decreased with further increases of the TFE concentration and reached 0 in 50% TFE. 2.2 Time course of the CIAP activation by TFE The time course of the CIAP activation during incubation in 40% TFE is shown in Fig. 2. The activity change of CIAP in 40% TFE is a multiphase process. The first phase is the activation reaction which is too fast to be measured by conventional methods. The subsequent activation process is apparently biphasic (Fig. 3). The activity change ended after about 170 min. The calculated activation rate constant for the fast phase was 1.049 (10-3 /s) which included 19.2% of the total change. The rate constant for the slow phase was 0.151 279 (10-3 /s) which included 61.1% of the total change. Thus, the first very fast phase included 19.7% of the total change. The results clearly show that the effect of TFE on the activation of CIAP is time-dependent. 2.3 Effect of TFE on the CIAP conformation The intrinsic fluorescence spectra of CIAP incubated in 0-70% TFE are shown in Fig. 4. The addition of increasing TFE (0 - 30%) concentrations caused a progressive slight increase of the intrinsic tryptophan fluorescence intensity. For TFE concentrations between 30%-60%, there was no significant change in the intrinsic tryptophan fluorescence spectra. Further increasing the TFE concentrations, up to 70%, caused a slight decrease of the intrinsic tryptophan fluorescence intensity. But there was no change in the fluorescence emission maximum wavelength at TFE concentrations lower than 60%. We also observed a slight red shift of the fluorescence emission maximum wavelength for higher TFE concentrations (70% TFE, from 331.5 nm to 334.5 nm) . TFE did not lead to any significant change in the intrinsic fluorescence emission maximum, indicating no significant change in the tertiary structure of CIAP. Measurement of the far-ultraviolet CD was employed to monitor the secondary structural changes of CIAP in TFE solutions of different concentrations (Fig. 5). For TFE concentrations from 0 to 20%, the average molecular ellipticity had only slight changes. But for further increases in the TFE concentration, the CD spectra of the protein changed noticeably and the average molecular ellipticity at 222 nm decreased in a concentration dependent manner (Fig. 6). The circular dichroism signal measured at 222 nm is mainly due to the a-helix. Therefore, the results indicate that TFE can significantly change the secondary structure of CIAP and increase the a-helix content. The time course of the change in the secondary structure of CIAP in 40% TFE was monitored by changes in the ellipticity at 222 nm.The results show that the change follows a very fast phase which achieves the final q222 value of the CD spectra in the dead-time of mixing (15 s) (data not shown). 3 Discussion In the previous study, we mentioned that the presence of TFE led to inactivation of CIAP when the TFE concentration in the subtrate system exceeded 30% and thus we concluded that inactivation of CIAP in TFE solutions is due to unfolding of CIAP by TFE[21]. In the present study, we repeated the previous experiments under the current testing condition and got the similar results. But further, the results of this study suggest that the CIAP activity increases when incubated in TFE solutions (0-50%) . So it should be reasonable that we now conclude that CIAP is activated at high concentrations of TFE. We also noticed that the CIAP activity was activated the most for the native enzyme added to the substrate system containing TFE concentrations lower than 15%. Further increasing TFE concentration inhibited the enzyme activity. The possible explanation for this phenomenon is that there are different effects of TFE on the CIAP activity. First, TFE directly affects the enzyme conformation, improving the enzyme activity. However, TFE also affects the interaction between enzyme and substrate, improving the enzyme activity for TFE concentrations in the substrate system lower than 15%, and inhibiting the protein activity at higher TFE concentrations. Thus, when the enzyme incubated in the buffer containing different TFE concentrations for 12 hours at 25 °C was added to the substrate system containing the same TFE concentrations, the final activity was the result of the two effects and the second one is more important. The cosolvent TFE, which has been used extensively to study protein structures, is known to induce and stabilize the a-helix, a secondary structure element in proteins, in regions that have an intrinsic propersity to form a-helix in aqueous solutions[22-26]. This study showed that TFE can significantly change the CIAP secondary structure, especially increasing the a-helix content, but the CIAP tertiary structure does not significantly change. The results suggest that the activation of CIAP is due to its secondary structural change, especially the increased a-helix content. To our knowledge, this is the first instance that increasing the a-helix content of a protein increased the protein activity. We have compared the time evolution of the activation with the change of the protein secondary structure. The acquisition of the secondary structure is completed within the dead time of 15 sand is thus extremely rapid compared to the activity change, which occurs on the time scale of hours. That the conformation changes do not coincide with activation suggests that in the very fast phase, the protein molecular structure is formed and followed by slower slight rearrangements of the side-chain packing in subsequent stages which can not be measured by conventional methods. It is well known that enzyme activity is dependent on conformational integrity. The effects of structural changes on the activity of enzymes have been extensively studied in the literature. The complete recovery of enzyme activity is usually thought to indicate the return to the native state such as folding of an enzyme in vitro. There are, however, some reported cases of CIAP with different structural forms exhibiting the same activity. Studies of CIAP renaturation have also shown that total recovery of the native-like structure is not necessary for the reconstitution of the active site, indicating that the active site domain can be reconstituted independently of other structural domains[13,14]. Our results also show that the higher catalytic activity of CIAP can be attained with much different secondary structural forms. Alkaline phosphatase is presented in large amounts in the liver, intestine, placenta, bone, and kidney of mammals[1]. Large amounts of alkaline phosphatase are found within the intralumenal contents of the calf intestine exhibiting a specific activity 20-times higher than that of the enzyme of the mucosa[27]. Although it is impossible that our results have direct physiological significance, this phenomenon, the activation of alkaline phosphatase by TFE which changes the secondary structure of the enzyme, might simulate the effect of how alkaline phosphatase activity is modulated in mammal cells by regulating the secondary structure of the protein through different physiological conditions. Studies of the structure-function relationship are always important in investigations of enzymes. The analysis of the effect of cosolvents on the structure and functions of proteins can lead to a better understanding of the structural, conformational and functional aspects of proteins and enzymes. Recent reports have shown that during the denaturation of a number of enzymes by GdnHCl, urea or heat, inactivation occurring before noticeable conformational changes of the enzyme molecule as a whole can be detected[28 - 31]. Therefore, it was suggested by Tsou that the enzyme active site is usually situated in a limited region of the enzyme molecule that is more flexible than the enzyme molecule as a whole and is therefore more easily perturbed than the rest of the molecule[32,33]. Recently, there were reports that chicken liver dihydrofolate reductase[34], adenylate kinase[35], and lipocalin-type prostaglandin D synthase[36] can be activated by denaturants (urea and guanidine hydrochloride) over a wide range of concentrations. Such effects suggest that the activation of enzymes may be due to the increasing conformational flexibility at active sites. But the detailed molecular mechanisms of activation are still unsolved. Further, structural analysis by multidimensional NMR or crystallography in the presence of TFE would be expected to clarify details of this mechanism. References
Copyright 2001 - Tsinghua Science and Technology The following images related to this document are available:Photo images[ts01092f5.jpg] [ts01092f2.jpg] [ts01092f1.jpg] [ts01092f6.jpg] [ts01092f3.jpg] [ts01092f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}