 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Tsinghua Science and Technology, December 2001, 6(5), pp. 432-437 TINY, a DREB-Like Transcription Factor Binding to DRE Element CAO Zhifang Department of Biological Sciences and Biotechnology,Tsinghua University, Beijing
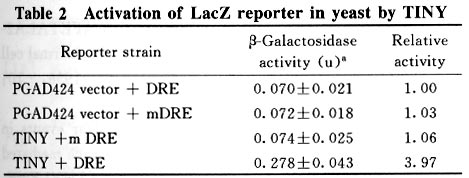
100084, China * Supported by the National Key Basic Research Science Foundation of China (G1999011703); the National Natural Science Foundation of China (No.39770167), and the Natural Science Foundation of Tsinghua University. Received: 2001-01-16; revised: 2001-06-04 Code Number: ts01093 Abstract: TINY isolated through a transposon-mutagenesis experiment designed to recover dominant gain-of-function alleles in Arabidopsis encodes a protein containing a putative DNA-binding domain which is conserved in AP2/EREBP transcription factors of plants. AP2/EREBP transcription factors play a variety of regulatory roles in several developmental processes and in response to biotic and environmental stresses. Using the yeast one-hybrid analysis system, we demonstrated that TINY could function as a DREB-like transcription factor specifically binding to the dehydration-responsive element (DRE) cis-acting element. Key words: TINY, transcription factor; dehydration-responsive element (DRE) Introduction Recently, AP2/EREBP transcription factors, a novel type of transcription factors, have been isolated from a variety of higher plants. AP2/EREBP-related genes form a large family which consists of many members found in several plant species, such as Arabidopsis thaliana, tobacco, tomato, rice, maize, castor bean, etc[1,2]. All the proteins of the AP2/EREBP family contain one (or two) conserved DNA-binding domain(s) composed of 57-70 amino acid residues, which are named the AP2/EREBP domain. All AP2/EREBP proteins possess typical structural characteristics of transcription factors, such as a DNA-binding domain (AP2/EREBP domain), nuclear localization signals (basic amino acid sequences), and putative transcription activation domains (acidic regions, serine-rich regions, etc.). The members of the AP2/EREBP family can be divided into at least two groups: AP2 and EREBP groups, according to the number of AP2/EREBP domains. The AP2 group includes AP2 and ANT found in ~iArabidopsis,~i Glossy 15 and ids1 found in maize. The genes of this group encode the proteins that contain two AP2 domains[3-7] which may function as transcriptional regulators and thus play important roles in plant developmental processes. The genes classified in the EREBP group encode proteins possessing only one AP2/EREBP domain. Many genes, such as EREBP1-4, Pit4-6, DREB1A-C (CBF1-3), DREB2A-B, AtERF1-5, and AtEBP, belong to this group[8-16]. Some of the members have been found to function as transcription factors (Fig. 1). The members of this group can be further divided into three subgroups based on their binding to different cis-acting elements. As shown in Fig.1, the members in the first subgroup (EREBP subgroup) include EREBP1-4 of tobacco, Pit4-6 of tomato, AtERF1-5 and AtERP of Arabidopsis, etc. These transcription factors have been experimentally shown to recognize and bind to the GCC box. The core sequence of the GCC box is AGCCGCC, which is the ethylene-responsive element identified from the genes encoding the b-1, 3-glucanase, PR-1 and PR-5d proteins of tobacco. It has been shown that the GCC box is present in the 5‘ untranscribed regions of almost all the basic pathogenesis-related genes. The second subgroup (DREBP subgroup) mainly consists of DREB1A-C (CBF1-3) and DREB2A-B of Arabidopsis. DREB1A-C and DREB2A-B specifically recognize and bind to the dehydration-responsive element (DRE) cis-element or DRE-like cis-elements involved in plant responses to drought, high-salt, and cold stresses[11]. DRE or DRE-like cis-elements frequently exist in promoters of plant genes (such as rd29A, rd17, kin1, etc.) induced by dehydration, high-salt, and cold stresses. The core sequence of DRE or DRE-like cis-elements is A/GCCGAC. The third subgroup (RAV subgroup) includes two members: RAV1 and RAV2[18] . Their N-terminal regions are homologous to the AP2/EREBP DNA-binding domain. It has been shown that the proteins encoded by RAV1 and RAV2 can bind to the CAACA motif in vitro[18]. The TINY gene, encoding a putative protein containing 218 amino acid residues, was isolated through a transposon-mutagenesis experiment designed to recover dominant gain-of-function alleles in Arabidopsis[19]. Enhancement of the expression of the TINY gene suppressed cell proliferation and exhibited pleiotropic effects, e.g. , reducing the height and elongation of hypocotyl and fertility. The function of the TINY protein remains unclear. The deduced amino acid sequence of TINY contains a conserved AP2/EREBP domain composed of 57 amino acids and located between amino acids 35 and 92 of the TINY protein. In addition, the putative protein of TINY has two other features of transcription factors: the N-terminus end (between amino acids 23 and 28) with basic amino acid sequences that can be considered as a nuclear localization signal (NLS) and the C-terminus (amino acid 140 - 218), a strongly acidic region, may serve as the transcription activation domain. Comparison of amino acid sequences of AP2/EREBP domains shows that TINY has high similarity to the DREB subgroup, especially to DREB1A - C , which strongly suggests that TINY might also be a DNA-binding protein belonging to the AP2/EREBP family. Further experimental evidence is needed. This study demonstrates that TINY can specifically bind to DRE cis-element in yeast using one-hybrid system. The results revealed that TINY could function as a DREB-like transcription factor. 1 Materials and Methods (1) The TINY gene was kindly provided by Dr. George Coupland (Department of Molecular Genetics, John Innes Centre, Colney, Norwich NR4 7UJ, United Kingdom). (2) To examine the transactivation activity of DREB protein, 3-tandem-copies of a 75 bp fragment (the wild type) containing a DRE cis-element from the promoter of the rd29A gene and 3-tandem-copies of a mutated 75 bp fragment (only DRE element TACCGACAT was replaced with TATTTTCAT) were inserted into EcoRI-MluI sites of MCS in the pHISi-1 expression vector and EcoRI-SalI sites of MCS in the pLacZi expression vector (MATCHMAKER one-hybrid system, Clontech), respectively[11]. Two recombinant plasmids were transformed into the yeast YM4271 strain for transactivation experiments. The recombinant yeast with two reporter genes (HIS and URA) grew in a selective medium plate with SD/His- Ura- and its growth was completely inhibited by 15 mmol/L 3AT (3-aminotriazole, a competitive inhibitor of the yeast HIS3 protein). (3) The full-length coding region of the TINY gene was prepared with polymerase chain reaction (PCR). The primer sets used for the amplification of the TINYgene were 5‘-AAAAGAATTCATGATAGCTTCAGAGAGTAC -3‘ and 5‘-AAAAGTCGACGACTTAATAATTA-TACAGTCCT-3‘. The PCR fragment was cloned to the EcoRI-SalI site of pBluescript II SK+ vector for sequencing. The verified full-length coding region was fused to the EcoRI-SalI site of the pGAP424 vector (MATCHMAKER one-hybrid system, Clontech). Transformation was carried out using 1 mg of pGAD424 plasmid recombined with TINY and by LiAc-polyethylene glycol according to the protocol of the one-hybrid system (Clontech). Transactivation experiments were carried out on a selective medium plate with SD/His- Ura- Leu- and 0 mmol/L,15 mmol/L, 30 mmol/L, 45 mmol/L, and 60 mmol/L 3AT. Quantitative b-galactosidase assays were performed as described by Guarente[20]. 2 Results This study used the yeast one-hybrid system to demonstrate the specific binding of the TINY protein to the DRE cis-element, which shares a similar AP2/EREBP domain with DREB proteins. Table 1 and Fig.2 show the transactivation results for TINY proteins in yeast cells. When pGAD424 plasmids containing TINY cDNA were transformed to a yeast reporter carrying wild-type DRE, the yeast cells grew normally in the presence of 15 mmol/L to 60 mmol/L 3AT. However, the yeast cells carrying the mutated DRE element (mDRE) did not show similar 3AT-resistance. TINY, as a DREBP protein, could not bind to the mDRE in which the CCGA residues were replaced by TTTT[11]. The results in Tables 1 and 2 and Fig.2 suggest that TINY specifically binds to the wild-type DRE sequence (CCGAC). 3 Discussion Recently, it has been reported that AP2 domains are present in numerous gene products with unknown functions in various plants. This strongly suggests that the AP2 domain is an important and evolutionarily conserved element, which is essential for these proteins to function[1,2]. Ohme-Takagi and his colleagues showed that the DNA binding preference of AtERF is consistent with those with only the minimum DNA binding domains[15,21], which suggests that the regions flanking the DNA binding domain are not critical for mediating the protein-DNA interaction in vitro[15,21,22]. Thus, the AP2 domain is essential and sufficient to confer the DNA binding activity of the EREBP-like group proteins. The solution structure of the AP2 domain of AtERF1 in complex with the GCC box has been recently determined[23]. The domain is composed of a three-stranded anti-parallel b-sheet and an a-helix packed approximately parallelto the b-sheet, with the b-sheet interacting monomerically with the target DNA. Moreover, a computer modeling experiment demonstrated that similar arrangements of the secondary structural elements was observed in other AP2 domains[22,23]. The precise amino acid residues within the AP2 domain required for DNA binding have been identified, and the amino acid residues of the AP2 domain of AtERF1 identified to contact the DNA bases are well conserved among the sequences of the DRE and GCC subfamilies. Therefore, it is likely that the DNA-binding sites of the AP2 domain for those two subclasses are the same. The conserved residues in the b-sheet region sequences of these two subclasses can be classified into three categories: those that are responsible for DNA recognition, those that stabilize the protein structure, and those that modulate the specificity. However, the exact amino acid residues of the third region remain unknown. Sequence comparisons and NMR results revealed that the C-terminal in all known AP2 domains contains a conserved core region that forms an amphipathic a-helix. This a-helical structure may mediate protein-protein interactions[2]. A search of the GenBank and expressed sequence tags (EST) data bases demonstrated that numerous plant genes encode proteins possessing an AP2 domain[1,2]. The sequence analyses indicated that they act as transcriptional regulators, but their target genes and binding sequences are unknown, which limits a full understanding of the role of the proposed regulator. Characterization of the amino acid residues of the AP2 domain determining the specificity binding to different cis-acting elements will be helpful for formulating hypotheses about the functions of proteins containing such sequence motifs. Detection of a functional site in a protein is of great interest. In general, a limited number of key residues are responsible for specific protein properties. Typically, in the protein family which consists of several subgroups, the physicochemical properties of amino acid residues determining different functions among the different subclasses are conserved within one subgroup, but differ from other subgroups[24-28]. To understand the molecular mechanisms of binding specificity between DREBP and EREBP subgroups, we compared the common amino acid residues of the b-sheets in each subgroup and between the two subgroups. We found four identifying sites in line with the rule that the properties are conserved in a subgroup: (1) The ninth site is the absolutely conserved proline (P) in EREBP, but this residue is not an aromatic amino acid in DREBP. (2) The fourteenth and nineteenth sites are valine (V) and glutamate (E) in DREBP, and alanine (A) and aspartic acid (D) in EREBP. (3) The twenty-sixth site is the absolutely conserved valine (V) in EREBP, but this residue is not valine in DREBP. We postulated that the four amino acid residues might determine the different binding specificities between the DREBP, and EREBP subgroups. This hypothesis is being tested. Comparison of the amino acid residues of the AP2 domains of TINY, DREBP, and EREBP showed that they share the amino acid residues required for DNA binding, and the four amino acid residues in TINY that were found to be conserved in each group were similar to those in DREBP. These observations suggest that TINY might bind to the DRE element. The results showed that TINY can bind to the DRE element in vivo (yeast expression assays) and can function as a transcription factor. Because the hypothesis has not yet been confirmed, we can not rule out the possibility that it is only by coincidence that TINY can bind to the DRE element. We proposed that although both DREB1 and DREB2 can bind the same cis-acting element (DRE element), they function as trans-acting factors in two separate signal transduction pathways for low-temperature and dehydration conditions, respectively[11]. However, the detailed biological function of TINY is not yet known. Identifying TINY as a transcriptional factor binding to the DRE element will help us discover the action function of TINY in Arabidopsis and the gene expression regulated by the DRE cis-acting element. Acknowledgements We are grateful to Dr. George Coupland for the TINY gene References
Copyright 2001 - Tsinghua Science and Technology The following images related to this document are available:Photo images[ts01093f1.jpg] [ts01093f2.jpg] [ts01093t2.jpg] [ts01093t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}