 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
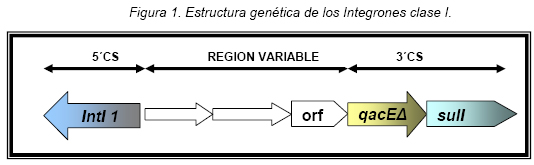
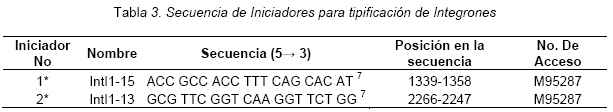
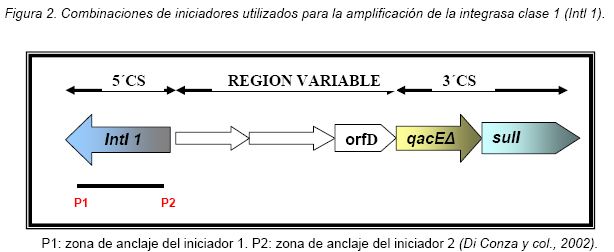
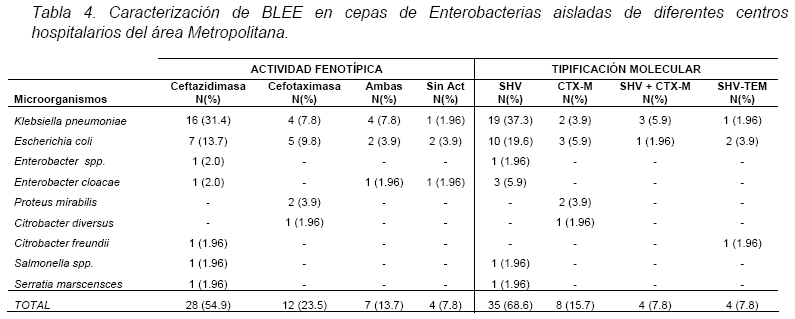
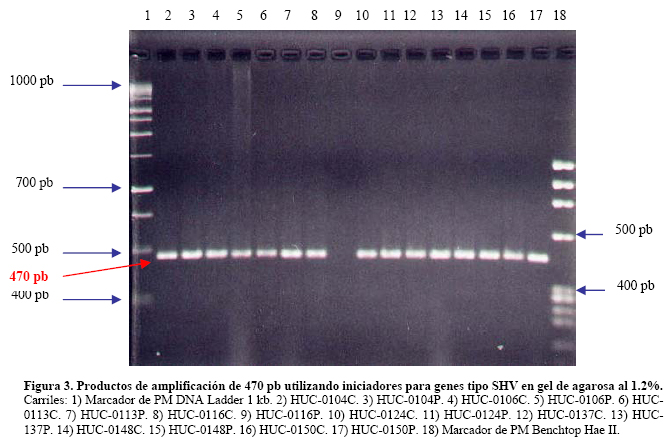
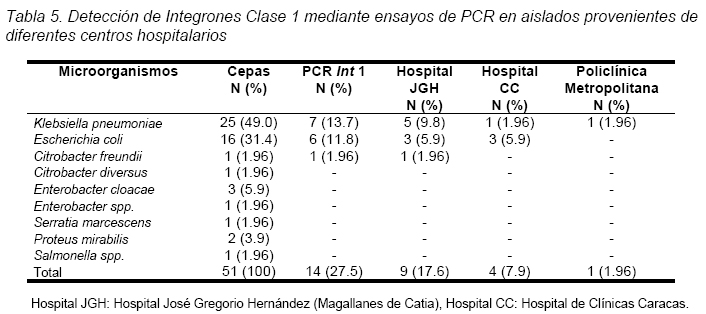
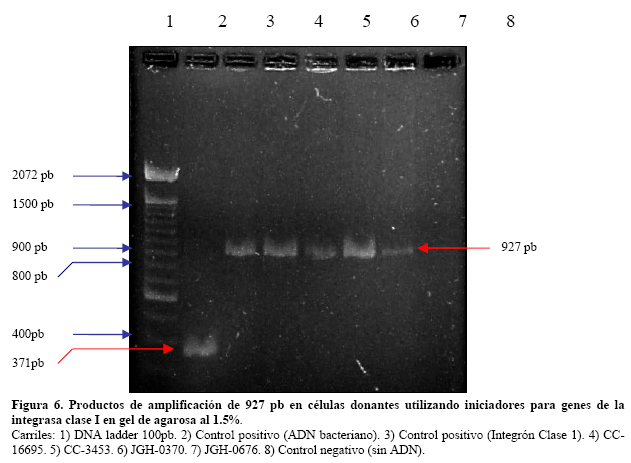
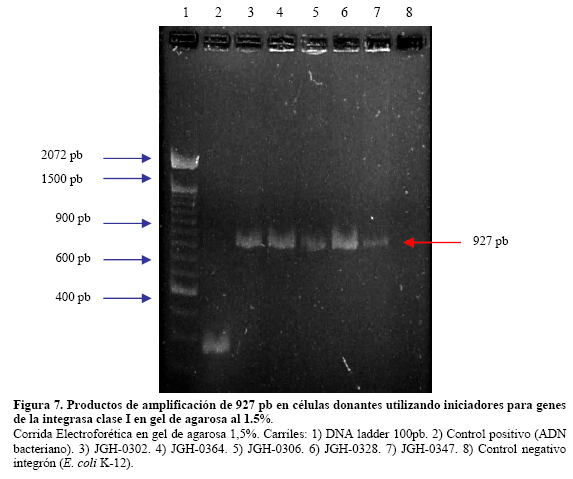
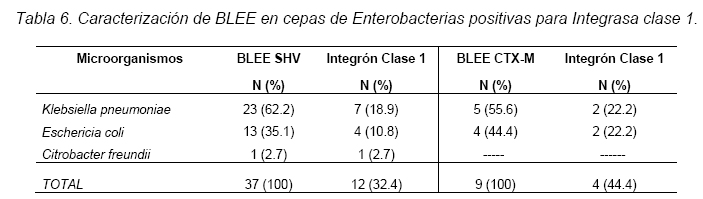
VITAE Academia Biomédica Digital, Número 25, Octubre - Diciembre 2005 Detección de integrones clase I en cepas de enterobacterias productoras de β-lactamasas de espectro expandido tipo SHV y CTX-M grupo-2 Torres, L.1,6; Benítez, M.2; Domínguez, M.2; Torres, O.2; Gagliotta, V.2; Calvo, A.3; Rodríguez, N.4; Ardila, J.5; Pedroza, R.6 1) Cátedra de Microbiología. Escuela de Bioanálisis. Facultad de Medicina. 2) Internado Rotatorio. Escuela de Bioanálisis. Facultad de Medicina. 3) Sección de Bacteriología. Policlínica Metropolitana. 4) Sección de Bacteriología Hospital “José Gregorio Hernández”. 5) Sección de Bacteriología Hospital de Clínica Caracas. 6) Sección de Diagnóstico Molecular de Agentes Infecciosos. Instituto de Medicina Experimental. Facultad de Medicina Experimental. Universidad Central de Venezuela. Caracas-Venezuela Fecha de Recepción 15 julio - Fecha de aceptación 15 septiembre Code Number: va05018 Resumen Las β-lactamasas de espectro expandido (BLEE) son enzimas capaces de hidrolizar el enlace amida cíclico de los oximino-β-lactámicos (cefotaxime, ceftazidime y aztreonam). Son codificadas en megaplásmidos de más de 80 Kda, que generalmente poseen otros genes de resistencia antimicrobiana. Recientemente se han descrito otras estructuras genéticas, denominadas integrones, mediante las cuales diversos genes pueden ser almacenados y diseminados entre especies. Poseen la información genética necesaria para expresar una proteína (integrasa) implicada en la captura y escinsión de genes de resistencia antimicrobiana. En esta investigación se detectaron integrones clase 1 en Enterobacterias productoras de BLEE. Se estudió una muestra de 51 Enterobacterias aisladas de siete centros hospitalarios de Caracas. Se realizó la detección fenotípica y molecular de BLEE. Seguidamente se transfirieron plásmidos mediante conjugación en medio sólido y se aislaron plásmidos de cepas donantes y transconjugantes. Por último, se detectaron integrones clase 1 mediante PCR, utilizando iniciadores para la integrasa clase 1. El 68.6% de las cepas presentan BLEE tipo SHV, un 15.7% tipo CTX-M grupo-2 y 7.8% portan ambos tipos de BLEE. De 36 cepas conjugadas, un 81% transfirió plásmidos portadores de genes que codifican BLEE. El análisis de los aislamientos plasmídicos, mostró en todas las cepas transconjugantes una banda de 25.000 pb y en un 80% se evidenció una banda plasmídica mayor a 50.000 pb. El 27,5% (14) de cepas donantes y sus respectivas transconjugantes son portadoras de integrones clase 1 asociados a plásmidos conjugativos. El 32.4% de las cepas con integrones producen BLEE tipo SHV mientras que el 44.4% BLEE CTX-M. Abstract The extended-spectrum-β-lactamases (ESβLs) are enzimes that can hidrolize the amide ciclic bond of the oxyminos-β-lactamics (cefotaxime, ceftazidime and aztreonam). These are encoded in megaplasmids of more than 80 kDa, that generally posees others genes of antimicrobial resistance. Recently, there have been described another genetic structures, called integrons, through which different genes can be stored and disseminated between strains. They posees the genetic information requiered to express a protein (integrase), implied in the capture and storing of antimicrobial resistance genes. In this investigation we detected class 1 integrons in Enterobacterias that produce ESβLs. We studied a sample of 51 Enterobacterias isolated from seven hospitals of Caracas-Venezuela. The fenotipic and molecular detection of ESβLs was realized, in the following, plasmids were transfered by conjugation in a solid media and isolated from donor and transconjugant strains. Finally class 1 integrons were detected by PCR, employing primers for the class 1 integrase. The 68.6% of the strains present ESβLs type SHV, 15.7% type CTX-M-group 2, and 7.8% carry both types of ESβLs. From 36 conjugated strains, 81% transfered plasmids carriers of genes that coded ESβLs. The analysis of the plasmidic isolates, showed in all the transconjugant strains, a band of 25.000 pb and in a 80% a plasmidic band larger than 50.000 pb was observed. The 27.5% (14) of donor strains and their respective transconjugants are carriers of class 1 integrons asociated to conjugative plasmids. The 32.4% of the strains with integrons produce ESβLs type SHV, instead the 44.4% produce ESβLs type CTX-M. Introducción La rápida aparición y diseminación de microorganismos resistentes a los agentes antimicrobianos, constituye un problema de salud pública a nivel mundial. La presión selectiva ejercida mediante el uso excesivo de antibióticos, junto a los diversos mecanismos de transferencia genética que portan las bacterias, contribuyen cuantiosamente con esta situación. En la actualidad, los β-lactámicos siguen constituyendo una de las mejores opciones terapéuticas antimicrobianas. Sin embargo, la efectividad de los mismos ha sido contrarrestada por la aparición de las β-lactamasas. Los genes que codifican estas enzimas pueden residir en el cromosoma bacteriano o en plásmidos8. Estas últimas de mayor relevancia ya que pueden ser transferidas a diversos géneros bacterianos y diseminarse en el ambiente hospitalario y en la comunidad. Las β-lactamasas de espectro expandido (BLEE) son enzimas que presentan un espectro incrementado de actividad hidrolítica, contra las cefalosporinas de tercera y cuarta generación y sobre el aztreonam. Son derivadas a partir de mutaciones puntuales de las β-lactamasas de espectro ampliado (BLEA) TEM-1, TEM-2 y SHV-117 , aunque existen otras familias de BLEE, como las tipos CTX-M y PER que tienen orígenes diferentes y una escasa relación estructural con las TEM y SHV. La movilización y estabilización de genes de resistencia a antibióticos ha sido conducida por los mecanismos de transferencia horizontal de la información genética, principalmente a través de la conjugación1, 2, 6 . La transferencia de ADN mediante conjugación, esta mediada por plásmidos conjugativos, que contienen toda la información genética necesaria para promover su propia transferencia1 . Plásmidos transmisibles y/o transposones permiten el intercambio de genes que codifican para determinantes de resistencia a múltiples antibióticos, entre especies poco relacionadas, representando el medio más común para adquirir determinantes de resistencia a antibióticos. En años recientes se han reportado la existencia de otra clase de unidades genéticas denominadas integrones. Estos, son elementos pertenecientes a la familia de transposones Tn21 y Tn3 1 , capaces de albergar genes de resistencia a antibióticos. Los integrones están constituidos por dos regiones de ADN muy conservadas, situadas en sus extremos. En la región conservada 5´ (5´CS) existen 3 elementos necesarios para la captura y expresión de genes exógenos (cassettes): uno que codifica una integrasa (int I), el otro es el lugar de recombinación sitio-específico (attI), y un promotor (Pant) para la expresión de los genes cassettes integrados. En la región conservada 3´ (3´CS) se hallan insertados el gen qacEA1 y el gen sulI 11 . Entre estas dos zonas se pueden insertar uno o varios tipos de genes (casettes génicos) de resistencia, en una zona denominada región variable 15 (Figura 1). De acuerdo con la secuencia de la integrasa se han clasificado los diferentes tipos de integrones. Se conocen nueve clases, los clase 1, 2 y 3 contienen genes de resistencia antimicrobiana. Algunos genes de β-lactamasas pueden estar codificados en dichos cassettes génicos 9, 19. Los integrones no son móviles por si mismos, pero con frecuencia se hallan asociados a transposones que a su vez se encuentran en plásmidos conjugativos, por lo que su movilidad horizontal esta asegurada 21 . Los integrones aparecen como estructuras genéticas que albergan genes de resistencia denominados cassettes génicos, permitiendo de esta manera el reconocimiento, la captación, intercambio y almacenamiento de nuevos mecanismos de resistencia, como es el caso de las β-lactamasas. En Venezuela son escasas las investigaciones que asocian la presencia de integrones clase 1 en bacterias multirresistentes aisladas de ambientes hospitalarios. Además, no existen en nuestro país reportes de la presencia de BLEE asociadas a este tipo de integrones. Es por ello la importancia de la identificación de integrones, especialmente en cepas de origen nosocomial, en las cuales la multirresistencia puede estar asociada a estas estructuras. En esta investigación se demostró la presencia de integrones clase I, asociados a plásmidos conjugativos in-vitro, en cepas de Enterobacterias y sus respectivas transconjugantes productoras de BLEE de las familias SHV y CTX-M. Metodología Muestra La población estudiada está conformada por 51 cepas de Enterobacterias productoras de BLEE, recolectadas entre los años 2002-2004. La distribución de las mismas fue la siguiente: K. pneumoniae (25), E. coli (16), P. mirabilis (2), Enterobacter spp. (1), E. cloacae (3), C. diversus (1), C. freundii (1), Salmonella spp. (1) y S. marcescens (1). Fueron aisladas de diversas muestras y provenientes de diferentes servicios de hospitalización, de los siguientes centros hospitalarios del área Metropolitana de Caracas: Hospital ¨José Gregorio Hernández¨ (23), Policlínica Metropolitana (6), Maternidad ¨Concepción Palacios¨ (5), Hospital Militar ¨Dr. Carlos Arvelo¨ (2), Instituto Clínico ¨La Floresta¨ (2), Hospital de Clínicas Caracas (12), Hospital de Niños ¨J. M. de los Ríos” (1). Ensayos de susceptibilidad antimicrobiana y detección fenotípica de β-lactamasas de Espectro Expandido (BLEE) Se llevo a cabo la metodología de Kirby-Bauer recomendada por la NCCLS 2004. Se determinó el perfil de resistencia a los siguientes antibióticos: amikacina (Ak), gentamicina (Ge), ciprofloxacina (Cip), trimetropin/sulfametoxazol (Sxt), piperacilina/tazobactam (Tzp), amoxicilina/clavulánico (Amc), cefoxitin (Fox), aztreonam (Atm), ceftazidime (Caz), cefotaxime (Ctx), cefepime (Fep), meropenem (Mem), imipenem (Imp). La detección fenotípica de BLEE, se realizó mediante el método de doble difusión en disco10 y por el recomendado por la NCCLS 2004. Los controles de calidad utilizados fueron: E. coli ATCC 25922-ATCC 35218 y Pseudomonas aeruginosa ATCC 27853, Klebsiella pneumoniae ATCC-700603. Detección molecular de BLEE mediante ensayos de Reacción en Cadena de la Polimerasa (PCR) Para realizar el PCR para detección de BLEE se trabajó según el esquema de Chanawong y col. (2000), utilizando células bacterianas completas. Los iniciadores utilizados para la detección de BLEE son mostrados en la Tabla 1. El protocolo utilizado para la reacción de PCR fue el siguiente: 4 min a 95 ºC, 30 ciclos de 1 min a 94ºC (desnaturalización), 1 min y 30 seg a 55ºC (alineamiento), 1 min a 72ºC (polimerización), y una extensión final de 10 min a 75ºC. Los reactivos y volúmenes utilizados están enumerados en la Tabla 2. Los productos de amplificación fueron visualizados mediante electroforesis en gel de agarosa al 1,2%. Se utilizó el marcador de PM ladder 100 pb. El control positivo para BLEE tipo SHV: K. pneumoniae M-1740. Para BLEE tipo TEM: E. coli M-3195. Para BLEE tipo CTX-M: K. pneumoniae M-1803. El control negativo: E. coli ATCC 25922. Todas las cepas controles positivos fueron donadas por el Dr. Marcelo Galas del Instituto de Enfermedades Infecciosas-ANLIS. Buenos Aires-Argentina. La caracterización de BLEE tipo SHV se llevó a cabo mediante ensayos de PCR-RFLP (reacción en cadena de la polimerasa y análisis de polimorfismos mediante el uso de enzimas de restricción). Una vez obtenido el producto de amplificación del gen codificante para β-lactamasas tipo SHV, se procedió a cortar el ADN con la enzima de restricción Nhe I. Esta enzima corta una secuencia especifica de nucleótidos generada en la posición 238 del gen, producto del cambio mutacional G----A, esta mutación es característica de varias BLEE derivadas de SHV-1 (SHV-2, SHV-5, SHV-12). De esta manera se logró determinar que el producto de amplificación correspondía a una BLEE derivada de SHV-1. Transferencia plasmídica de genes que codifican para BLEE y de Integrones clase I La transferencia de plásmidos que codifican para BLEE y que portan genes estructurales correspondientes a Integrones clase I, se llevó a cabo mediante ensayos de Conjugación Bacteriana en medio sólido. Las cepas donantes corresponden a un numero de 36 cepas de Enterobacterias que poseían los marcadores fenotípicos adecuados para dicho ensayo, es decir, presentaban sensibilidad aRifampicina y/o Ácido nalidíxico. La cepa receptora de plásmidos conjugativos fue la Escherichia coli K-12 NCTC50170, RC-711 rif (donada por el CVCM). La selección de cepas transconjugantes fue realizada en placas de medio agar Luria-Bertani (agar LB) suplementadas con rifampicina (40 µg/mL) y ampicilina (30 µg/mL) y una segunda placa de agar LB suplementadas con rifampicina (40 µg/mL) y ceftazidime ó cefotaxime (10 µg/mL). Aislamiento plasmídico Para el aislamiento de plásmidos se utilizó el método de lisis alcalina, empleando polietilenglicol y cloruro de litio4 . En el caso de cepas transconjugantes se utilizó la técnica a mediana escala y para las cepas donantes a pequeña escala, ambos cultivos con 24 horas de crecimiento en agitación a 37°C. El aislamiento plasmídico fue visualizado mediante electroforesis en gel de agarosa al 0,6%. Se utilizó el marcador de peso molecular del bacteriófago Lambda Hind III. Detección de Integrones clase I mediante ensayos de Reacción en Cadena de la Polimerasa (PCR) Se utilizaron iniciadores (Tabla 3) que reconocen secuencias específicas de la integrasa clase 1 (Figura 2), la detección de la integrasa se llevó a cabo a partir de las células bacterianas de las Enterobacterias y sus correspondientes transconjugantes y del ADN plasmídico aislado, según el protocolo de Di Conza y col. (2002)7. Los volúmenes de reactivos fueron similares a los usados en el PCR de identificación de BLEE. El protocolo utilizado para la reacción de PCR fue el siguiente: 6 min a 94 ºC, 30 ciclos de 45 seg a 95ºC (desnaturalización), 45 seg a 55ºC (alineamiento), un minuto a 72ºC (polimerización), y una extensión final de 10 min a 72ºC. El control positivo utilizado fue Lisado celular obtenido de la cepa HUC-0117, la cual presenta un integrón clase 1 14 y el negativo fue un lisado celular preparado a partir de la cepas de Escherichia coli K-12 ó Escherichia coli ATCC 25922, las cuales no presenta ningún integrón. Resultados Detección fenotípica y molecular de BLEE Mediante el análisis de los niveles de resistencia (CIMs) para las cefalosporinas de espectro expandido, se evidenció que el 54.9% de las cepas productoras de BLEE muestran una mayor actividad de tipo ceftazidimasa. Mientras que solo un 23.5% de las cepas estudiadas mostró una fuerte actividad hidrolítica para el cefotaxime y valores de resistencia o susceptibilidad intermedia para el cefepime. Igualmente hay que señalar que 7 (13.7%) cepas poseen un patrón fenotípico característico de ambos tipos de BLEE, es decir, actividad marcada frente a ceftazidime y cefotaxime (Tabla 4). Los ensayos de detección molecular mediante PCR revelaron que el mayor porcentaje de BLEE circulantes en estas cepas corresponde a la familia SHV (presuntamente SHV-5) con un 60.78% (Figura 3, Tabla 4). Mientras que un 15.68% de las cepas amplificaron con los iniciadores específicos para genes de la familia CTX-M del grupo 2 (Figura 4, Tabla 4). Así mismo se encontró que 4 (7,8%) cepas presentan genes que codifican para las BLEE tipo SHV y CTX-M concomitantemente. Por otra parte, hubo resultados de amplificación positiva para genes de la familia TEM, pero siempre en asociación con genes de BLEE tipo SHV (Tabla 4). Ensayos de transferencia plasmídica de genes que codifican para BLEE y de Integrones clase I. De las 51 cepas de Enterobacterias se conjugaron 36 (69%), debido a que estas poseían los marcadores fenotípicos adecuados para el ensayo. De estas 36 Enterobacterias se obtuvieron 29 cepas transconjugantes, es decir 81% de las cepas transfirieron material plasmídico a la cepa receptora (E. coli K-12). A las 21 cepas restantes, se les realizó aislamiento plasmídico de manera directa, y se evidencio mediante PCR la presencia de genes de BLEE tipo SHV y CTX-M, de la misma manera se determinó la presencia de integrones clase 1 sobre ADN plasmídico. En todas las transconjugantes obtenidas mediante ensayos de conjugación, se detecto fenotípicamente la producción de BLEE, de la misma manera se identifico el tipo de BLEE por PCR, en las transconjugantes y en todos los casos se identifico el mismo tipo de BLEE que portaba la Enterobacteria donante. Hay que destacar, que se observó la adquisición de resistencia por parte de las transconjugantes al cefoxitin en un 7%. Aislamientos plasmídicos El 100% de los aislamientos plasmídicos obtenidos en cepas transconjugantes, presentan una banda con un peso molecular en el rango de 25000 pb. Igualmente se observa que en más del 80% de las cepas existe una banda con un peso molecular superior a los 50000 pb. Por otra parte se evidencia que un 44.8% de las cepas poseen dos bandas con un peso molecular más bajo, de aproximadamente 3500 pb y 2500 pb respectivamente (Figura 5). En los aislamientos plasmídicos de las cepas donantes se observa una banda con un peso molecular de aproximadamente 25000 pb, similar a la observada en las cepas transconjugantes. Menos del 5% de las cepas donantes posee una banda por debajo de los 2000 pb, que no se observa en la electroforesis de las transconjugantes. En el resto de las cepas el patrón de aislamiento plasmídico es similar tanto en cepas donantes como en sus respectivas transconjugantes (Figura 5). Detección de Integrones clase I Se realizó la detección de Integrones clase I sobre las 51 cepas de Enterobacterias y las 29 transconjugantes obtenidas. Un total de 14 (27.5%) cepas de Enterobacterias amplificaron con iniciadores específicos para el gen de la integrasa clase 1, este porcentaje incluye, K. pneumoniae (7), E. coli (6) y C. freundii (1) (Tabla 5). De las 14 cepas que resultaron positivas para integrones clase 1, nueve (64.3%) cepas provienen del Hospital ¨Dr. José Gregorio Hernández¨, cuatro (28.6%) cepas del Hospital de Clínicas Caracas y una (7.2%) cepa de la Policlínica Metropolitana. A continuación, se observan productos de amplificación con células completas en el rango de 900 pb, la misma se correlaciona con el tamaño estimado teóricamente, mediante la búsqueda de la secuencia de la integrasa clase I en el Gen Bank, que es de 927 pb (Figura 6). En la corrida anterior, se muestra en el carril 1 el marcador de peso molecular, en el carril 2 el producto de amplificación de 371 pb del control positivo (E. coli K-12) de ADN bacteriano. En el carril 3, se observa el producto de amplificación de 927 pb del control positivo (E. coli HUC-0117) de integrones clase 114 . En los carriles 4 y 5 se observa la amplificación de cepas de Escherichia coli, CC-16695 y CC-3453, respectivamente. En los carriles 6 y 7 se observan dos cepas de Klebsiella pneumoniae, ambas provenientes del Hospital ¨Dr. José Gregorio Hernández¨. En la Figura 7, se muestra en el carril 3, el producto de amplificación de una cepa de Citrobacter freundii proveniente del Hospital ¨Dr. José Gregorio Hernández¨, en el carril 4 se muestra otra cepa de E. coli y por último, en los carriles siguientes (5 al 7) cepas de K. pneumoniae, todas provenientes del mismo centro hospitalario. De las 14 cepas de Enterobacterias, en las cuales hubo amplificación para genes de la integrasa clase 1, sólo nueve de ellas poseían cepas transconjugantes y a este grupo se les realizó PCR, para evidenciar la presencia de integrones. La amplificación para genes de la integrasa clase I, fue positiva para las nueves cepas transconjugantes obtenidas por conjugación (Figura 8). Los datos obtenidos indican que de las 14 cepas portadoras de integrones clase I, el 32.4% de las mismas producen BLEE de tipo SHV y un 44.4% producen enzimas de la familia CTX-M (Tabla 6). Discusión Los patrones de resistencia para las diversas cefalosporinas de espectro extendido son similares en las cepas donantes y transconjugantes en estudio, se observa un mayor perfil hidrolítico para el ceftazidime y aztreonam, mientras que los valores de resistencia son más bajos para el cefotaxime, este comportamiento evidencia que hay una mayor proporción de BLEE con actividad ceftazidimasa (SHV y/o TEM) 22. También se observaron cepas con una mayor actividad hidrolítica para el para cefotaxime y menores niveles de resistencia para el ceftazidime, por lo cual fenotípicamente se tratan de cepas con actividad cefotaximasa, pudiéndose tratar de cepas productoras de BLEE tipo CTX-M, Por otra parte, se evidenciaron cepas que mostraban una fuerte actividad hidrolítica para ambos sustratos, pudiéndose tratar de cepas que de manera concomitante expresan diversos tipos de BLEE. Los ensayos de PCR, evidenciaron que 68.6% de las cepas son portadoras de genes SHV que presentan la mutación G------A en la posición 238 del gen. Entre las principales BLEE SHV que tienen esta mutación se encuentran: SHV-2, SHV-5, SHV-9, SHV-11, SHV-15 y SHV-34. Se encontró un 15.7% de cepas que codifican BLEE tipo CTX-M grupo 2, todas estas cepas presentan fenotípicamente una fuerte actividad hidrolítica contra cefotaxime y valores de resistencia o susceptibilidad intermedia para cefepime 12, 18, 20. El análisis de los aislamientos plasmídicos sugiere que desde el punto de vista epidemiológico, en nuestros centros hospitalarios probablemente circulan y se transfieren plásmidos con características similares. De la misma manera se puede inferir que los genes que codifican para BLEE están circulando en estos plásmidos de alto peso molecular, situación reflejada a nivel mundial 5 . Estos plásmidos de alto peso molecular se pueden estar diseminando de manera activa entre diferentes géneros y especies si las medidas de control y barrera no son óptimas, en este estudio se pudo observar una alta transmisibilidad plasmídica entre diferentes Enterobacterias mediante ensayos de conjugación. Con respecto a la adquisición de resistencia al cefoxitin de las cepas transconjugantes, a partir de cepas de K. pneumoniae y E. coli, microorganismos que normalmente no son productores de β-lactamasas tipo AmpC., evidencia la probable presencia de plásmidos conjugativos que portan genes para β-lactamasas que hidrolizan a las cefamicinas (enzimas tipo AmpC) en el ambiente hospitalario, dificultando cada vez más la detección de estos mecanismos de resistencia, a la vez que se limitan las opciones terapéuticas. Los resultados de PCR, utilizando iniciadores específicos para integrasa clase 1 señalan que un 27,5% de las cepas analizadas, son portadoras de integrones clase 1. Estos integrones se han encontrado principalmente en especies pertenecientes a la familia Enterobacteriaceae, y la mayoría de los determinantes de resistencia encontrados en la región variable de estos elementos codifican para la resistencia a aminoglicósidos. La frecuencia de integrones clase 1 en las Enterobacterias de un ambiente hospitalario, varía de acuerdo al hospital y al período en que son analizados los aislados clínicos 11 . Los aislados clínicos utilizados en este trabajo pertenecen a diversos géneros, sin embargo, la mayoría de los aislados positivos corresponden a Klebsiella pneumoniae, seguido de Escherichia coli y sólo una cepa de Citrobacter freundii presentó estos elementos. La determinación de la presencia de integrones clase 1, en las cepas transconjugantes permitió establecer que estos elementos se encuentran alojados en plásmidos conjugativos, ya que en la totalidad de las cepas transconjugantes se identificaron integrones Clase I. Entre los aislados clínicos que presentan integrones, el 44.4% son productoras de BLEE tipo CTX-M y un 32.4% son productoras de BLEE tipo SHV. La resistencia a ceftazidime, cefotaxime y aztreonam en Enterobacterias es frecuentemente derivada de la expresión de β-lactamasas de espectro expandido tipo TEM y SHV, las cuales no han sido reportadas en cassettes génicos dentro de un integrón. Sin embargo, el significante incremento en la resistencia a ceftazidime y aztreonam en aislados positivos para integrones provee una evidencia circunstancial de la asociación entre integrones y BLEE, probablemente por un plásmido hospedero común 15 . Hasta el año 2002, no existían reportes previos con descripciones de genes codificantes para BLEE tipo de CTX-M asociados a integrones. En estudios por Di Conza y col. (2002) 7, se reporta un nuevo integrón clase 1 denominado InS21, el cual transporta un gen de BLEE CTX-M-2 en una cepa de Salmonella entérica serovar. Infantis. En esta misma publicación se mencionan evidencias preliminares de la presencia de genes de BLEE CTX-M-2 asociados a integrones clase 1 en un plásmido de alto peso molecular de una cepa de Morganella morganii. En esta investigación se demostró la presencia de integrotes clase I en cepas productoras de BLEE de la familia CTX-M, este hallazgo pudiera indicar que los genes rodantes para estas enzimas se encuentren asociados o localizados en integrones clase 1. Sin embargo, es conveniente realizar estudios de biología molecular más específicos para confirmar dicha asociación. Aunque la expresión de los genes de resistencia no sea igual entre las diversas especies, la presencia de integrones supone una gran ventaja selectiva para las bacterias que portan tales elementos genéticos. Estas estructuras, presentan una elevada transmisibilidad y captación por parte de las bacterias, sobre todo en aquellas de origen nosocomial. Esto puede deberse a que no es necesario que la bacteria acumule grandes piezas de material genético, como son los transposones y/o plásmidos para hacerse resistente a un determinado antibiótico, sino que con pequeños segmentos de ADN en el interior del integrón se alcanza la resistencia 15 . A pesar de que los integrones no tienen movilidad genética, los mismos pueden ser diseminados en el ambiente hospitalario a través de plásmidos conjugativos y transposones, en esta investigación se demostró que estas secuencias genéticas están vinculadas a plásmidos transferibles, esta situación debe ser tomada en cuenta por las diversas comisiones de Enfermedades Infecciosas, a la hora de establecer y desarrollar las diversas medidas de control y barrera en sus respectivas Instituciones Hospitalarias. Referencias Bibliográficas
Copyright 2005 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela The following images related to this document are available:Photo images[va05018t5.jpg] [va05018t3.jpg] [va05018t2.jpg] [va05018f1.jpg] [va05018t6.jpg] [va05018t1.jpg] [va05018f7.jpg] [va05018t4.jpg] [va05018f8.jpg] [va05018f3.jpg] [va05018f2.jpg] [va05018f4.jpg] [va05018f6.jpg] [va05018f5.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}