 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
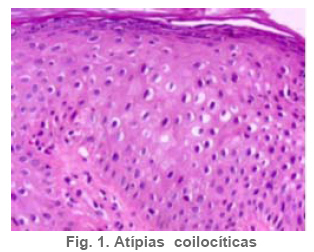
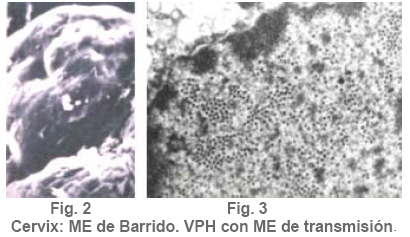
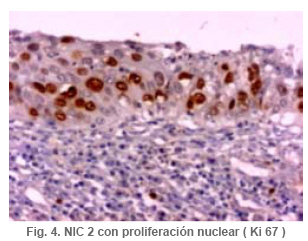
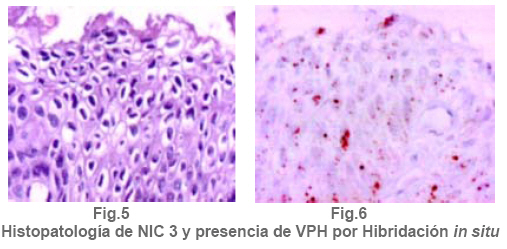
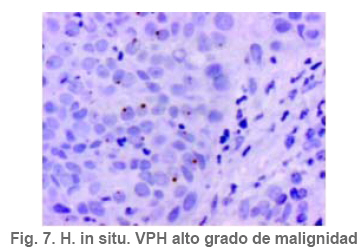
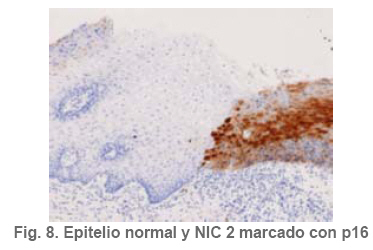
Anatomía Patológica Actualización sobre la historia del virus del Papiloma Humano en Venezuela y su relación con el cáncer cervical Jorge García Tamayo Anatomopatólogo Laboratorio de Patología Molecular NOVAPATH, Maracaibo, Venezuela. Fecha de recepción: 21/04/2006 Fecha de aceptación: 23/05/2006 NOTA: Toda la información que se brinda en este artículo está destinada al conocimiento general. En ningún caso sustituye el asesoramiento de un médico. Ante cualquier duda que pueda tener en cuanto a su estado de salud, consulte con su médico o especialista. Copyright 2006 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela Code Number: va06009 Palabras Claves: vph, papiloma humano, cáncer cervical Introducción En octubre del año 1971, se dio en Maracaibo, Venezuela, el VIII Congreso Latinoamericano de Patología. Un ginecólogo (Jorge Nágel) y un patólogo (Elio Casale), ambos venezolanos que ejercían en Caracas, demostraron en un trabajo libre la presencia de partículas virales en unas lesiones del cuello uterino que aparecían como manchas rosadas; para ello habían utilizado el microscopio electrónico (ME). José Trinidad Núñez Montiel era un ginecólogo reconocido como colposcopista en Suecia y en Argentina, donde para la época iba a hablar en los congresos sobre “mosaicos” y “recuadrados”, unos términos esotéricos para casi todos los ginecólogos venezolanos (en esos tiempos ni los norteamericanos le daban mucha importancia al colposcopio como arma diagnóstica). JT Núñez Montiel sí, él era capaz de demostrar -utilizando una pinza que abría todo el canal endocervical y usando lugol- dónde estaban los focos de displasia en los sitios más recónditos del cuello uterino. Cuando Nuñez Montiel escuchó lo que decían Nagel y Casale en el VIII Congreso de la SLAP, inmediatamente pensó que podíamos, con la ayuda del colposcopio y de un microscopio electrónico (ME), resolver el problema del origen del cáncer cervical. Ya habíamos examinado muestras del cérvix, mirado las tricomonas y su efecto sobre el epitelio cervical, habíamos tomado muestras de casos de tricomoniasis con el ME y estábamos pensando en comunicar nuestros hallazgos en unos trabajos que serían publicados unos años más tarde (1, 2, 3, 4). Sabíamos que los virus del Papiloma Humano producían las verrugas en la piel y los condilomas en las mucosas, los cuales evidentemente eran verdaderos tumores. Aquellos crecimientos con apariencia de coliflor, los condilomas genitales, eran tumores ginecológicos que se veían en mujeres de escasos recursos, y el cáncer y la pobreza nos parecía que iban juntos. Tan pronto terminó el Congreso de la SLAP del año 1971, nos dimos a la tarea de tomar biopsias y de buscar hasta detectar las partículas del VPH en el cuello uterino. Vimos los virus en exudados, en el núcleo de muchas células y destacamos características ultraestructurales que no se conocían ya que las muestras histológicas tan solo mostraban cambios que eran para aquel tiempo muchas veces interpretados como áreas con sobrecarga glucogénica. Entenderíamos con la sencilla tinción con el reactivo de Schiff- que no era glicógeno lo que modificaba el epitelio exocervical. Era difícil convencer a mis colegas patólogos de que los halos coilocíticos en los cortes histológicos no eran glucógeno. Cuando Núñez Montiel les hablaba de los virus, sencillamente se reían, pues para la época, nadie nos creía. (Ver las atípias coilocíticas en la Figura 1). Dos años después, en el siguiente IX Congreso Latinoamericano de Patología (1973) presenté en un par de trabajos libres nuestros resultados. Fue en septiembre en Mérida, en la península mexicana de Yucatán. Al final de la presentación de ambos trabajos, el Dr. Javier Arias Stella que había estado presente se me acercó para decirme que seguramente yo no tenía idea de la importancia de lo que acababa de mostrarles, pero que era algo crucial. El tema se retoma Con las vueltas que da la vida, sería en San Sebastián, País Vasco, a comienzos de la década de los noventa del pasado siglo XX, cuando volveríamos a revivir aquellos días. Don Javier Arias Stella tuvo la deferencia de comentar este hecho en una hermosa charla sobre nuestra especialidad dictada en el País Vasco, en unos lluviosos días de noviembre vividos con Saudy en unión de tan buenos amigos, allá en Donostia, frente al mar cantábrico. Su conferencia fue reproducida en 1993 en la Revista Patología de la Sociedad Latinoamericana de Patología (SLAP), en México (5). La presentación de los dos trabajos libres del IX Congreso de la SLAP en Mérida (Yucatán, México, en septiembre de 1973) había sido publicada en las memorias del congreso (Suplemento de la Revista Patología (Mex) mas como es de suponer, su divulgación fue muy pobre (6, 7). La historia formal sobre el VPH y las publicaciones alrededor de este virus y su relación con el cáncer, para todos, se iniciarían con los cambios citológicos descritos por Meisels y Fortín (8), por Puroloa y Savia (9) en 1976 y por el germano ZurHausen unos años más tarde (10). Una publicación relacionada con nuestros hallazgos, habría de producirse un par de años después en 1978, en una revista médica venezolana que para aquella época llevaba un buen record de publicación ininterrumpida (11). El trabajo resumió nuestras experiencias durante los años de trabajo en Maracaibo y quiso ser un homenaje al increíble personaje que había sido JT Nuñez Montiel. Por ello se iniciaba con este epígrafe: “Esta investigación sobre la papilomatosis cérvicovaginal es el producto de las inquietudes y el tesón de José Trinidad Núñez Montiel, brillante ginecólogo zuliano, hoy ya desaparecido. Él sintió muy de cerca el abatimiento y la decepción al ver como sus colegas dentro y fuera de la Universidad del Zulia, no valoraron sus esfuerzos para hacer investigación sobre el cáncer del cuello uterino. Este trabajo es el fruto de sus ideas y espero pueda servir como homenaje póstumo a su memoria”. Unos años más tarde, con Francisco (Pancho) Rincón Morales -quien nos decía con orgullo sentirse discípulo de las ideas y de la obra de JT Nuñez Montiel-, nos interesamos en el tema de las lesiones vulvares provocadas por el VPH y volvimos a hacerles, durante los años vividos en la Universidad Central de Venezuela, estudios con el ME de transmisión e hicimos ME de barrido y de esas observaciones surgieron varias publicaciones (12, 13, 14). Ver ultraestructura de barrido y de transmisión del VPH en las Figuras 2 y 3. El carcinoma cervical seguía punteando nuestras estadísticas de mortalidad por cáncer y, entretanto, el ejercicio ilegal de la Medicina por mucha gente que leía muchas muestras citológicas sin supervisión de patólogos, nos llevaron a múltiples discusiones y controversias con el MSAS y con los citotecnólogos. Nuestra posición como gremio fue respaldada varias veces por la Federación Médica Venezolana. Lejos de mejorar el despistaje citológico, las desavenencias recrudecieron en la medida que comencé a publicar por escrito mis reflexiones. Algunas en la prensa de Caracas y de Maracaibo y quizás especialmente por las verdades que decía un libro titulado “Reflexiones de un Anatomopatólogo” (15), una publicación auspiciada por la Directiva de la Sociedad Venezolana de Anatomía Patológica en el año 1991. En “Más reflexiones sobre la Patología y el País” -1998- (16), se incluían artículos de prensa sobre el tema en cuestión, publicados en El Nacional, Panorama, El Diario de Caracas, etc. En ellos se iba repitiendo la historia de aquello a lo que acostumbraba a denominar Pancho “la cifra ignominiosa”. Nuestra cruel estadística que muestra como con el tiempo, luego de más de 50 años de hacer despistaje de cáncer por citología, lejos de haber mejorado, ahora la curva del carcinoma cervical en Venezuela va en ascenso. Esto nos ha parecido siempre una muestra de subdesarrollo o quizás de la entropía tropical, ese desorden e irresponsabilidad que ha caracterizado desde siempre a quienes en posiciones de mando deberían haber adoptado -con seriedad y organización- medidas para que el cáncer del cuello uterino estuviese desapareciendo de nuestra tierra. Con el argumento de “la idiosincrasia”, o mas bien por nuestra particular manera de ver la cosas superficialmente, sin ir al meollo del asunto, el tiempo ha transcurrido, sigue pasando y lejos de mejorar las condiciones de las pacientes -que son la mayoría del pueblo que no tiene recursosse deteriora cada día más. Los políticos usualmente se ocupan de llenarse los bolsillos o hablan mucho y hacen muy poco y por eso es que pareciera más sensato pensar que paradójicamente, el culpable de todo puede que sea “el excremento negro del diablo”, como denominara Pérez Alfonso al petróleo que ha hecho tan rica esta nación y tan pobres a sus ciudadanos. No obstante, se pueden buscar estadísticas, siempre muy atrasadas y cada vez más maquilladas, sobre las cifras del cáncer cervical, sin poder ocultar que no existe un Registro Nacional de Cáncer confiable. (Ver en la Figura 4 marcada con el anticuerpo Ki67, la proliferación nuclear en una lesión NIC2). La mayoría de los carcinomas del cuello uterino se inician como alteraciones intraepiteliales, por lo que frecuentemente existen lesiones sincrónicas y metacrónicas en el epitelio cervical displásico y neoplásico, que son provocadas por la expansión clonal de células epiteliales transformadas. Ahora se sabe que en la transformación de las células del cáncer cervical siempre han mediado eventos genéticos previos y se conoce que ellos están asociados a la presencia del VPH en la génesis de esta neoplasia, lo cual ha sido señalado desde hace muchos años y es en la actualidad parte consustancial del proceso de carcinogénesis. Con el correr de los años, se han descrito más de 200 tipos de VPH y estos han sido clasificados en 16 grupos (17). Los VPH de tipo genital se clasifican de acuerdo con el potencial para provocar cambios malignos en el epitelio cervical en 3 tipos. 1- De alto riesgo: (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68, 73 y 82), 2- de riesgo intermedio: (26, 53 y 66) y 3- de bajo riesgo: (6, 11, 40, 42, 43, 44, 54, 61, 70, 72, 81 y CP6108) (18). Entre los virus de alto riesgo, los más conocidos son los 16 y 18 que se observan en un 50% y 20% de los casos de carcinoma epidermoide respectivamente (19). No obstante, tan solo una minoría de los casos infectados con los VPH de alto riesgo se transformará en cáncer. En las figuras 5 y 6 se observa la histopatología de una lesión cervical NIC3 y la presencia de VPH detectada por hibridación in situ. Descripción del VPH El VPH pertenece a la familia Papovaviridae, que está formada por virus que tienen una cápside con 72 capsómeros con el genoma del virus. Los capsómeros están compuestos por dos proteínas estructurales: L1 que forma el 80% de la partícula viral y L2 que es la proteína menor de los cápsidos, de 43- 53 kD. El genoma es una molécula de doble cadena de ADN organizada con 8 a 10 marcos de lectura abierta (ORFs) la cual es semejante para todos los VPH. El genoma está dividido en tres regiones: la región larga de control (LCR) que no tiene potencial para codificar, la región de las proteínas tempranas (E1-E8) y la región de las proteínas tardías (L1 y L2). Las proteínas E1 y L2 son necesarias para la replicación extracromosómica del virus y existen para poder completar el ciclo vital del virus. Una de las características del cáncer cervical es la pérdida de la expresión de la proteína viral E2. Se cree que en las células basales del epitelio neoplásico, esta proteína, E2, pierde su expresión y esto produce una pequeña cantidad de E2-ORF, el cual como producto de fusión es capaz de represar la replicación del ADN y su expresión y por lo tanto mantendría el virus en fase latente en las células basales (20,21). Las proteínas E1 y E2 se van a unir a secuencias específicas del ADN, de tal modo que E1 es la proteína iniciadora de la replicación, para lo cual las secuencias únicas de E1 en adenina-timina van a estar flanqueadas por dos o tres sitios de unión con la proteína E2, con alta afinidad para las secuencias de adenina-timina. E2 regula la trascripción y, como se dijo, es capaz de reprimir algunos promotores que controla la expresión de los genes E6 y E7 y de esta manera es un regulador de la proliferación celular y de la capacidad de transformación de las células infectadas. La proteína E4 se expresa en estadios tardíos de la infección cuando se están ensamblando los viriones, en tanto que la proteína E5 en un marco de lectura abierta está demorada en las células neoplásicas y ella no es necesaria para mantener la transformación maligna de las células huésped. En la etapa no productiva episomal, las células basales del epitelio llegan a formar hasta 200 copias con gran amplificación del genoma. Posteriormente, se transcribirá el ARNm y se van a codificar las proteínas L1 y L2 de la cápside, cosa que se produce únicamente en las capas superficiales del epitelio, allí se pueden comenzar a ver con el microscopio electrónico las partículas virales. La infección persiste en las células basales porque ellas no sufren cambios, no hay coilocitosis, no hay lisis celular, no son productivas. Las proteínas E6 y E7 son, sin duda alguna, las más importantes en sus propiedades oncogénicas. Ellas mantendrán el ambiente celular para que el genoma viral pueda mantenerse extracromosómico. Hoy en día se conoce bien la capacidad de E6 y E7 para inmortalizar y transformar las células del cuello uterino. Los modelos de E6-p53 y de E7-Rb han servido para demostrar como el VPH provoca los cambios preneoplásicos (22,23). El blanco de E6 es la proteína supresora de tumores conocida como p53, pues E6 promueve su degradación a través de la ubiquitina. De manera que las células que expresan E6 tienen bajos niveles de p53, lo cual favorece la acumulación de mutaciones en los cromosomas. La mayor parte de las alteraciones cromosómicas en el cáncer cervical, se han encontrado en los brazos de los cromosomas 3p, 4p, 5, 6, 11q y 17p; éstos cambios conllevan un fenómeno que es crucial en el proceso de transformación maligna: la pérdida de la heterocigosis (LOH)( 24 - 27 ). En los epitelios planoestratificados, los estudios del fenómeno LOH en la carcinogénesis temprana señalan que el cromosoma más frecuentemente implicado es 3p, pero si se desglosan las alteraciones cromosómicas en las distintas fases del cáncer cervical, podríamos señalar que en NIC se ve LOH en 3p, 5p y 5q, 6p y 6q, 11q, 13q y 17p; en el carcinoma invasor se ve LOH en 3p, 6p y 6q, 11q, 17p y 18q; en las metástasis ganglionares se observa LOH en 3p, 6p, 11q, 17p, 18q y en el cromosoma X. Todos estos eventos se inician con el fenómeno denominado inestabilidad cromosómica, por ello, es importante señalar que al menos en la mitad de los casos de NIC de bajo grado, es posible detectar alteraciones de genes supresores en 6p, las cuales se ponen en evidencia en el 90% de los casos de NIC de alto grado (28). Algunos de estos resultados han sido revisados por nosotros recientemente (29). Es un hecho comprobado que son las oncoproteínas E6 y E7 de los VPH de alto riesgo las que más se relacionan con p53 y con el gen del retinoblastoma (Rb), los cuales llevan a la hiperproliferación, inmortalización e inestabilidad genética del epitelio cervical e, igualmente, conducen a la malignización asociada a los tipos de VPH de alto riesgo. E6 puede funcionar como factor de transcripción celular por la homología de su estructura con presencia de “dedos de zinc”, dominios éstos que pueden unirse al ADN para activar la transcripción (30). El proceso de apoptosis es interferido a través de p53 y de otros mecanismos en los que influye E6 para evitar la muerte celular programada. Esto se logra a través del bloqueo del ciclo celular por el inhibidor de la cinasa dependiente de ciclina p21 y Bax, que es una proteína proapoptótica presente en los epitelios indiferenciados y que puede ser degradada por E6 a través de la ubiquitina. En la Figura 7 se demuestra el inmunomarcaje en los núcleos de un cáncer cervical, con hibridación in situ, utilizando una sonda para virus de alto grado de malignidad. Ya hemos señalado que el sitio de replicación del VPH es el epitelio bien diferenciado y se considera que también influye E6 para evitar el proceso de diferenciación terminal del epitelio escamoso. Se ha demostrado que E6 del VPH-16 es capaz de interactuar con proteínas relacionadas con el calcio dentro del retículo endoplasmático. Algunas proteínas citoplasmáticas que poseen dominios PDZ se unen al extremo C-terminal de las proteínas de E6 de VPH de alto riesgo e interaccionan con proteínas que se relacionan con el control de la uniones intercelulares y con la proliferación celular, por lo que se presume que influyen en los contactos intercelulares y en el control de la polaridad y la división celular (31). En cuanto a E7, su principal papel consiste en inalterar el control del ciclo celular. Esta proteína es, principalmente, nuclear; su expresión está asociada al incremento de la síntesis del ADN celular y de la proliferación celular. De la interacción de E7 con las proteínas de la familia Rb se produce la inhibición del factor de transcripción E2F, por tanto, E7 forma un complejo con Rb y su degradación se promueve a través del sistema de ubiquitinización (32). Por otra parte, además de degradar a Rb, otros inhibidores del ciclo celular como p21 y p27, son controlados por E7. Precisamente el efecto de la inactivación en los genes del epitelio cervical se ha investigado a través de la expresión de p16, una proteína que reacciona con más énfasis al producirse la inactivación de Rb por la proteína E7 del VPH. Hoy en día se considera que p16, la Ciclina D1 y Ki 67 pueden ser marcadores biológicos complementarios que ayudan a examinar la relación de VPH con la capacidad de invasión inicial del CC. Al utilizar la inmunohistoquímica y la hibridación in situ para VPH, investigamos esta situación en casos de cáncer cervical. De ello, algunos resultados previos han sido remitidos y presentados en eventos internacionales; en trabajos de investigación en curso, ( Blasco Olaetxea, E, Guzmán Bistoni C, Molina J, Méndez V, y col. ) al utilizar anticuerpos, lectinas y la detección de grupos sanguíneos en las membranas de los queratinocitos del cervix, estamos examinando el papel que pueden jugar las glicoproteínas de membrana y p16 en el comportamiento biológico del epitelio cervical. La presencia de p16 en una lesión de NIC contrasta con el epitelio cervical normal en la Figura 8. Referencias bibliográficas
Copyright 2006 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela The following images related to this document are available:Photo images[va06009f7.jpg] [va06009f8.jpg] [va06009f2-3.jpg] [va06009f5-6.jpg] [va06009f4.jpg] [va06009f1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}