 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
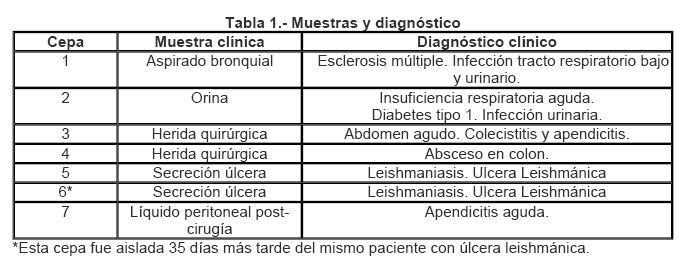
VITAE Academia Biomédica Digital, Número 27, Abril-Junio, 2006 Microbiología Variabilidad en los exoproductos de Pseudomonas aeruginosa aislada de diferentes muestras clínicas Nina Polanco Daza1, Reyna Moronta2, José Antonio O'Daly3 1olancon@camelot.rect.ucv.ve ,
Inmunología Laboratorio de Bacteriología, Escuela de
Bioanálisis,
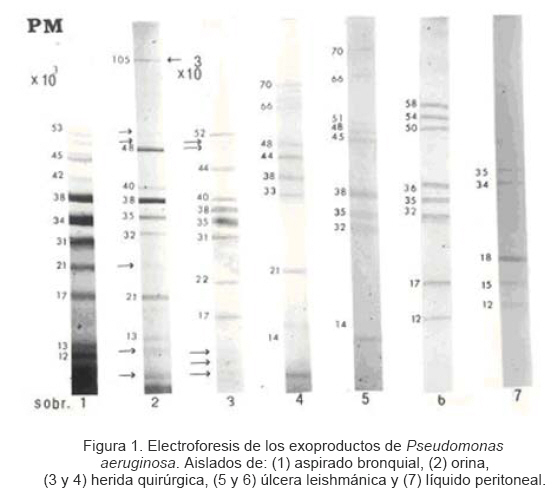
Universidad Central de Venezuela Fecha de recepción: 05/04/2006 Fecha de aceptación: 08/05/2006 NOTA: Toda la información que se brinda en este artículo está destinada al conocimiento general. En ningún caso sustituye el asesoramiento de un médico. Ante cualquier duda que pueda tener en cuanto a su estado de salud, consulte con su médico o especialista. Code Number: va06013 Pseudomonas aeruginosa es un bacilo Gram negativo que puede aislarse de diferentes ambientes especialmente en hospitales. Sintetiza una variedad de factores que contribuyen con su patogenicidad, por tal razón se planteó analizar los exoproductos de P. aeruginosa aisladas de pacientes del Hospital Vargas de Caracas-Venezuela. La electroforesis SDS-PAGE de los exoproductos de las distintas cepas reveló variación tanto en número de bandas proteicas como en pesos moleculares de las mismas. El mayor número lo presentó la cepa aislada de aspirado bronquial y el menor la cepa aislada de líquido peritoneal. Una banda de 35.000D apareció en 86% de las cepas, una banda de 105.000D solamente fue evidente en el aislado de orina y reportamos en este trabajo cuatro bandas diferentes a las descritas por otros autores. Se concluye que P. aeruginosa sintetiza y excreta in vitro diferentes factores que pudieran estar relacionados con el cuadro clínico de la enfermedad. Palabras Claves:Electroforesis, Pseudomonas aeruginosa, muestras clínicas, exoproductos. Abstract Pseudomonas aeruginosa is a Gram negative bacilli isolated from different environments mainly from hospitals being considered a nosocomial pathogen. The pathogenic factors synthesized by this bacteria are rarely investigated, thus we analyzed by SDS PAGE the exoproducts of P. aeruginosa isolates from different clinical conditions in patients from the Hospital Vargas, Caracas, Venezuela. We found variability in the number of protein bands as well as in their molecular weights from different isolates. The highest number of protein bands was found in the bronchial isolate while the lowest number in the isolate from peritoneal fluid. A band of 35,000 D appeared in 86% of the strains, a band of 105,000 D was only evident in the isolated from urinary tract. We conclude that P. aeruginosa synthesizes in vitro differents factors that could be related to the clinical localization of the infection. Key Word: Electrophoresis, Pseudomonas aeruginosa, clinical samples, exoproducts. Introducción Las infecciones causadas por las bacterias Gram negativas se han incrementado en relación con aquellas causadas por las bacterias Gram positivas (1, 2, 3). De particular interés es Pseudomonas aeruginosa, un bacilo Gram negativo que en los últimos treinta años ha aumentado su frecuencia como agente etiológico de diversas infecciones en humanos, generalmente de origen nosocomial, donde se ha identificado como agente causal de septicemias (4), endocarditis infecciosa (5, 6, 7), infecciones del tracto respiratorio inferior (8, 9), meningitis (10, 11, 12) e infecciones oculares (13, 14, 15), entre otras patologías. P. aeruginosa no solamente tiene un papel importante como agente de infección nosocomial, también puede causar infecciones en individuos de la comunidad, de allí que se ha aislado de esputo de personas con neumonía extrahospitalaria (16, 17, 18), sinusitis e infecciones del tracto urinario (19, 20, 21) y otitis (22). Al igual que en la mayoría de los patógenos bacterianos, la virulencia de P. aeruginosa es multifactorial y es el producto de la interacción de muchas variables que involucran tanto a la bacteria como al hospedador. Estas interacciones múltiples contribuyen sin lugar a dudas al amplio espectro que presenta la enfermedad causada por este bacilo en los pacientes infectados. La patogenicidad o virulencia de esta bacteria se ha asociado con numerosos factores, algunos son parte estructural de la célula y otros son sintetizados por ella y excretados al tejido circundante donde se desarrolla la infección (23). Los factores de virulencia descritos son: 1- Exotoxina A, la cual es muy letal, ya que ejerce un papel inmunosupresor sobre los linfocitos B y T humanos (23, 24, 25); 2- Exoenzima S, también es una enzima muy virulenta que media la fijación bacteriana a las células del hospedador y actúa como inmunosupresora (23, 26); 3- Proteasas que producen necrosis en piel, pulmón, córnea, además de la inactivación proteolítica del interferón gamma humano y del factor de necrosis tumoral (TNF) (27, 28); 4- Elastasa, capaz de producir hemorragia (27, 29); 5fosfolipasa C, hemolisina termolábil que induce la liberación de mediadores inflamatorios (30, 31, 32); 6- Pili, que aumenta la virulencia al mediar la adherencia a las células epidérmicas tanto normales como aquellas provenientes de piel quemada (17, 33); 7- Exomucopolisacárido (Slime), que tiene la capacidad de alterar marcadamente la motilidad, endocitosis y formación del fagosoma de los granulocitos neutrófilos humanos normales (34, 35). Estos factores no siempre se han podido identificar, en todos los aislados de Pseudomonas aeruginosa, esto sugiere la existencia de variabilidad antigénica en este bacilo, por lo cual se planteó investigar la variabilidad fenotípica en los productos excretados in vitro en cepas de P. aeruginosa aisladas de pacientes con diferentes cuadros clínicos. Materiales y Métodos Electroforesis Aislamiento e identificación de Pseudomonas aeruginosa. El material de estudio está constituido por siete cepas de Pseudomonas aeruginosa, aisladas de pacientes de ambos sexos con edades comprendidas entre 17 y 60 años, que fueron hospitalizados en el Hospital Vargas de Caracas, Venezuela, y quienes presentaban diferentes patologías ocasionadas por esta bacteria (Tabla 1). Las muestras se obtuvieron según las normas del Código de Bioética y Bioseguridad, capítulos 2 y 3 (36) y fueron procesadas según la metodología de Thompson y Millar, 2003 (37). Las cepas fueron aisladas e identificadas en el Laboratorio de Bacteriología de la Cátedra de Microbiología en la Escuela de Bioanálisis, Facultad de Medicina de la Universidad Central de Venezuela, siguiendo la metodología de Kiska y Gilligan, 2003 (38). Exoproductos de las cepas de Pseudomonas aeruginosa. Las cepas previamente identificadas como Pseudomonas aeruginosa se sembraron por aislamiento en agar Mac conkey (Difco) y se incubaron a 35ºC de 18 a 24 horas. Posteriormente, se recolectaron las colonias necesarias para hacer una suspensión cuya turbidez fue comparable con el patrón Nº 1 de Mac Farland (39). Un ml de cada suspensión se agregó a 150 ml. de medio de cultivo Eagle (Gibco) libre de proteínas para evitar enmascarar los exoproductos propios de la bacteria, se incubaron a 35ºC con agitación constante, por el período de 18 a 24 horas, tiempo después del cual se recolectaron las bacterias mediante centrifugación a 27.138 x G por 10 minutos a 4ºC en una centrifuga Sorvall, rotor G16 y el sobrenadante obtenido se filtró en una membrana Millipore de 0,22 µm de diámetro de poro. A continuación, se dializó contra agua destilada, durante un lapso de 48 horas a 4ºC. Transcurrido este tiempo, las muestras se liofilizaron y se resuspendieron en 2 ml. de PBS pH 7.2, se les determinó la concentración de proteínas mediante el método de Lowry y Col., 1951 (40) modificado por Herbert y col, 1983 (41) y se conservaron a -20ºC hasta su uso. Electroforesis Los productos excretados por Pseudomonas aeruginosa se separaron mediante electroforesis en gel SDS-poliacrilamida al 10%, siguiendo el método de Laemmli, 1970 (42). En cada pozo del gel se colocaron 25 µl de cada muestra que contenía 2 mg. de proteínas en buffer de disociación. La electroforesis se realizó en un aparato Miniprotean III (Bio-Rad) en buffer de corrida durante un período aproximado de 3 horas a 100 voltios y a temperatura ambiente. Posteriormente, las proteínas se fijaron con Etanol 30%-AcOH 7%, se colorearon con Comassie brillant blue al 0,05% y se sometieron a decoloración hasta evidenciar las bandas. Como control de pesos moleculares (Bio-Rad) se utilizaron: Albúmina bovina (66.000 D), Ovoalbumina (45.000 D), Anhidrasa carbónica (31.000 D) y Lisozima (14.400 D). Resultados La Tabla 1 presenta las muestras clínicas y el diagnóstico clínico de los pacientes infectados por Pseudomonas aeruginosa. La figura 1 muestra la electroforesis en gel de SDS-poliacrilamida de los exoproductos de Pseudomonas aeruginosa aislada de pacientes con diferentes infecciones causadas por esta bacteria (Tabla 1). La electroforesis se realizó en diferentes espacios de tiempo y con diferente gel, usando los mismos patrones de pesos moleculares. Los pesos moleculares de las diversas bandas proteicas fueron calculadas con la curva de regresión lineal (Quantity One Gel Documentation System-Bio-Rad) mediante los controles de peso molecular ya mencionados. La columna 1 representa los exoproductos de una cepa aislada de aspirado bronquial; en ella se evidencian 13 bandas con pesos moleculares que oscilan entre 54.000 y 12.000 D, distribuidas de la siguiente manera: cinco bandas entre 45.000 y 54.000 D, cuatro bandas entre 42.000 y 31.000 D (31.000, 34.000, 38.000 y 42.000 D) y cuatro bandas entre 21.000 y 12.000 D (12.000, 13.000 D, 17.000 y 21.000). Las bandas con pesos moleculares de 31.000, 34.000 y 38.000 D aparecen más pronunciadas. Las bandas de 53.000 y 42.000 D no aparecen en los otros productos de estas cepas de Pseudomonas aeruginosa. La banda de 38.000 D, aparece en la cepa aislada de orina (columna 2), en la de herida quirúrgica (columna 3 y 4), secreción de úlcera leishmánica (columna 5); están ausentes en la cepa aislada 35 días más tarde del mismo paciente con úlcera leishmánica (columna 6) y en la cepa aislada de líquido peritoneal. La banda de 35.000 D aparece en todos los exoproductos menos en la cepa aislada de una herida quirúrgica por acceso en cuello (columna 4). La banda de 21.000 D también aparece en la cepa asilada de orina y de herida quirúrgica (columna 2 y 4) y la banda de 17.000 D solamente se evidencia en la columna 1, 3 y 6 (Fig. 1). La columna 2 corresponde a los exoproductos de una cepa aislada de una muestra de orina de un paciente masculino. Se evidencian 13 bandas con pesos moleculares que oscilan entre 105.000 y 7.000 D, las cuales están distribuidas de la siguiente manera: una banda de 105.000 D, tres bandas de 48.000, 49.000 y 50.000 D, cuatro bandas 32.000, 35.000, 38.000 y 40.000 D, una banda débilmente entre 32.000 y 21.000 D, una banda de 21.000 y 13.000 D y dos bandas una de 12.000 y otra de 7.000 D aproximadamente. En esta columna destaca la banda de 105.000 D, la cual no aparece en ninguno de los exoproductos del resto de las cepas bacterianas. La columna 3 presenta los exoproductos de una cepa aislada de herida quirúrgica abdominal. En ella se observan principalmente ocho bandas con pesos moleculares que oscilan entre 52.000 y 17.000 D: (52, 44, 40, 38, 35, 31, 22 y 17) x 1000 D y otras bandas menos pronunciadas de 50.000 y 48.000 D y tres bandas entre 12.000 y 7.000 D. La columna 4 muestra los exoproductos de una cepa aislada de secreción de herida quirúrgica por acceso en cuello. En ella se observan diez bandas con pesos moleculares entre 70.000 y 7.000 D. Se encuentran dos bandas con 70.000 y 66.000 D, entre éstas aparece una débilmente coloreada de aproximadamente 68.000 D. Estas bandas de 70.000 y 66.000 D también aparece en la cepa aislada de una secreción de úlcera leishmánica (columna 5). Se observan cuatro bandas más de 48.000, 44.000, 38.000 y 33.000 D, una con 21.000 D, otra de aproximadamente 15.000 D y una de 7.000 D. Hay ausencia de proteínas con peso molecular entre 66.000 y 48.000 D y 33.000 y 21.000 D. La columna 5 muestra los exoproductos de una cepa aislada de secreción de una úlcera leishmánica, en ella se observan nueve bandas entre pesos moleculares de 70.000 y 13.000 D, distribuidas de la siguiente manera: (70, 66, 51, 48, 45, 38, 35, 32 y 13) x 1000 D. Hay ausencia de bandas en el rango de peso molecular entre 66.000 y 51.000 D, 45.000 y 38.000 D y entre 32.000 y 13.000 D. La columna 6 representa los exoproductos del segundo aislado de Pseudomonas aeruginosa realizado 35 días más tarde de la úlcera leishmánica. En ella se observan ocho bandas con pesos moleculares entre 58.000 y 12.000 D, estas son: (58, 54, 50, 36, 35, 32, 17 y 12) x 1000 D. Es notoria la ausencia de bandas mayor de 58.000 D, así como entre 50.000 y 36.000 D y entre 32.000 y 17.000 D. La columna 7 muestra los exoproductos de una cepa aislada de líquido peritoneal posterior a cirugía por apendicectomía. Se observa que solamente presentó cinco bandas protéicas con pesos moleculares entre 35.000 D y 12.000 D, estos son: (35, 34, 18, 15 y 12) x 1000 D. Se nota la ausencia de bandas con peso molecular mayor a 35.000 D y entre 34.000 y 18.000 D. Discusión Las Pseudomonas aeruginosa pueden excretar una variedad de factores, muchos de los cuales han sido asociados con su virulencia, esta característica aunada a los patrones de resistencia a los antimicrobianos que posee, inciden directamente en la evolución de la enfermedad ocasionada por esta bacteria. En el presente trabajo se demuestra a través de electroforesis en gel de poliacrilamida, que las cepas analizadas elaboran y liberan factores proteicos diferentes, tanto en el número de bandas como en sus pesos moleculares. La cepa aislada de aspirado bronquial produjo el mayor número de bandas, mientras que la cepa aislada del líquido peritoneal produjo el menor número de ellas (Fig. 1). Existen reportes que indican que en las muestras del tracto respiratorio bajo se identifican la exoenzima S, la exotoxina A y proteasas en proporciones de 85, 79 y 65%, respectivamente (43), en nuestro caso y a juzgar por los PM de éstas enzimas (entre 66.000 y 71.500 D para la exotoxina A y 105.000 D para la exoenzima S) no se evidenciaron bandas con estos pesos moleculares ya que la mayoría se ubicaron entre 12.000 y 54.000 D. El único exoproducto con una banda proteica de peso molecular similar a la de la exoenzima S (105.000 D) fue el producido por la cepa aislada de orina (Fig.1.1). Aunque hace dos décadas la exoenzima S no era muy común en los aislados clínicos (46, 47), pero en la última decada se ha identificado con alta frecuencia en cepas aisladas de orina (Hamood y col, 1996 (43), Rumbaugh y col., 1999 (44) y Rumbaugh y col., 1999 (45), su mayor prevalencia en la actualidad podria estar relacionada a la frecuencia de P. aeruginosa como agente infeccioso de diversos cuadros clínicos, incluyendo las del tracto urinario sobre todo en pacientes hospitalizados. Si bien no evidenciamos la exotoxina A en los exopoductos de la cepa aislada de aspirado traqueal, si parece estar presente en las cepas aisladas de la herida de absceso en cuello y de una úlcera leishmánica (Fig. 1. columna 4 y 5). Tomando en consideración que la exotoxina A se aísla con mayor frecuencia en muestras de heridas (43), se esperaría encontrarla también en las cepas aisladas de herida de colón y de la úlcera leishmánica 35 días más tarde después del aislado inicial. Sin embargo, su ausencia (Fig.1 columna 3 y 6) podría interpretarse por la variación fenotípica que pudiera ocurrir en la bacteria en una infección crónica, debido a la presión inmunológica inducida por el hospedador y/o por la presión de los tratamientos antimicrobianos suministrados al paciente (49). Es notorio que en casi todos los aislados (Fig. 1. Columna 1, 2, 3, 5 y 6) aparecen bandas que por sus pesos moleculares de 49.000, 50.000 y 52.000 D pudieran ser proteasas, ya que en esta bacteria se han reportado varias proteasas con pesos moleculares dentro del rango 48.000 y 54.000 D (50, 51). Aunque las proteasas son factores comunes en la mayoría de los aislados de Pseudomonas aeruginosa en este trabajo no se pudo evidenciar en los aislados de la herida quirúrgica en cuello (Fig. 1 columna 4) ni en el de líquido peritoneal (Fig. 1 columna 7). Existe una gran variedad de proteínas de pesos moleculares menores a los descritos anteriormente, algunos de ellos se encuentran entre 34.000 y 38.000 D, los cuales están presentes en todos los aislados y podrían interpretarse como adhesinas, cuyo papel es en la adherencia no mediada por Pili de Pseudomonas aeruginosa a la matriz celular (52) y esta función explicaría su presencia en la mayoría de los aislados de las muestras clínicas. Otras, que se encuentran en el rango de 12, 13, 21.000 están en los exoproductos de la cepa aisladas de aspirado bronquial, orina y absceso en cuello. Es notoria en los exoproductos de las cepas 2, 3 y 4 la presencia de una proteína de 7.000 D, la cual por su peso molecular podría tratarse de flagelinas (53). Estos resultados demuestran que las cepas de Pseudomonas aeruginosa aisladas de diferentes cuadros clínicos presentan variabilidad fenotípica manifestada en sus productos extracelulares. Esta variabilidad parece depender del cuadro clínico del paciente y por ende de la muestra clínica de donde fueron aisladas. En consideración de esto, si tal variabilidad ocurriese en la infección in vivo, el conflicto entre el hospedador y la bacteria disminuiría, ya que se eliminarían los efectos patógenos del (los) factor (es) no excretados al medio circundante. Por ejemplo, se ha demostrado que aquellos pacientes infectados con cepas no productoras de exotoxina A tienen mejor pronóstico en la enfermedad (24, 54, 55). En conclusión estos resultados demuestran que las cepas de diferentes tejidos en los diversos cuadros clínicos en humanos presentan variabilidad fenotípica in vitro. Esta variación no parece ser codificada geneticamente (44, 45) ni tampoco depende del medio de cultivo, ya que todas las cepas se cultivaron en las mismas condiciones. De acuerdo con esto, Woods y col, 1986 (56), encontraron variabilidad en los productos excretados por Pseudomonas aeruginosa pero en la concentración de las enzimas dependiente de la muestra clínica aisladas de los pacientes estudiados. Resalta el número de bandas proteicas evidenciadas en los exoproductos de aspirado bronquial, orina y herida quirúrgica no están descritas por otros investigadores, cuyo papel patógeno habría que definir dada la importancia de esta bacteria como agente infeccioso. Agradecimientos: Los autores agradecen su colaboración a la Unidad de Microbiología y Enfermedades Infecciosas del Hospital Vargas de Caracas, Venezuela. Referencias Bibliográficas
Copyright 2006 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela The following images related to this document are available:Photo images[va06013f1.jpg] [va06013t1.jpg] |
| |||||||||
{kind=link}
{kind=link}