 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
VITAE Academia Biomédica Digital, Número 30, Enero - Marzo, 2007 Neurociencia Estudio Electrofisiológico de la relación temporal entre la onda Theta y la actividad neuronal unitaria en el hipocampo de ratas Electrophysiological study of the temporal relationships between theta waves and neuronal unitary activity in rat hippocampus 1 Rafael Salas, 2Francisco Hernández 1rasalas@cantv.net
Neurofisiología
Cátedra de Fisiología, Escuela de
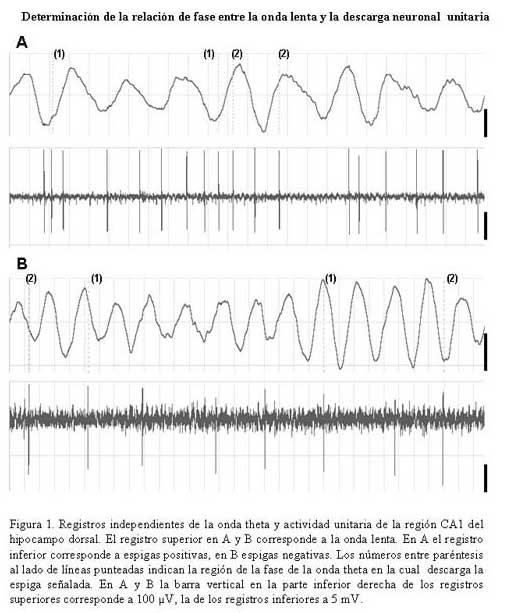
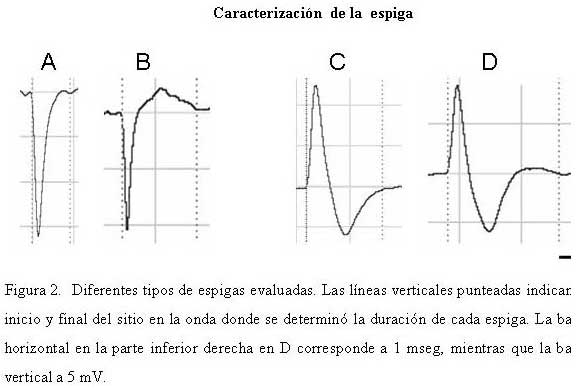
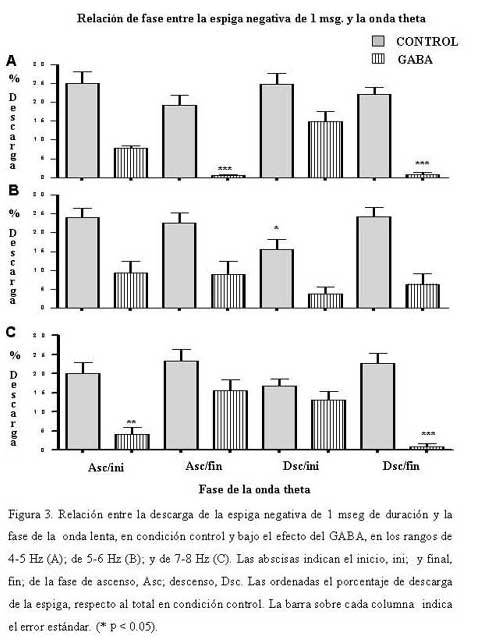
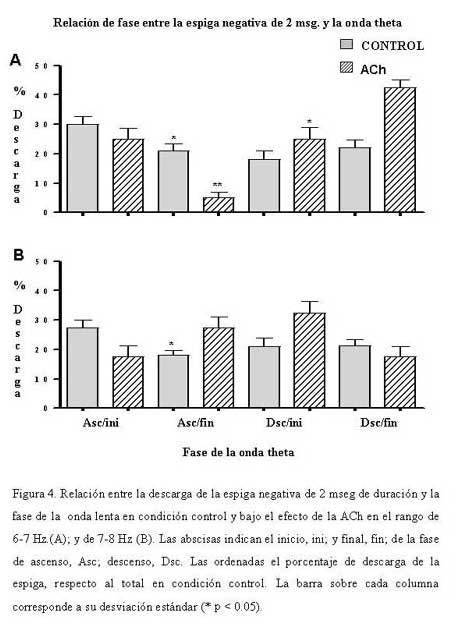
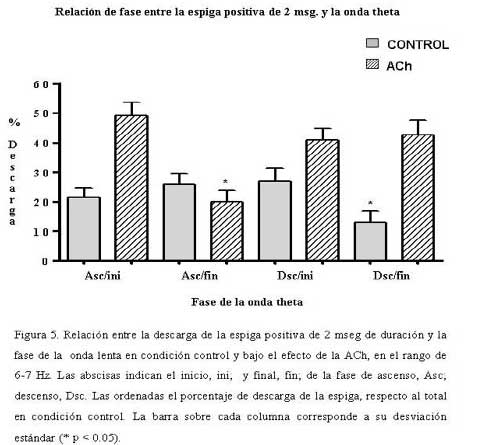
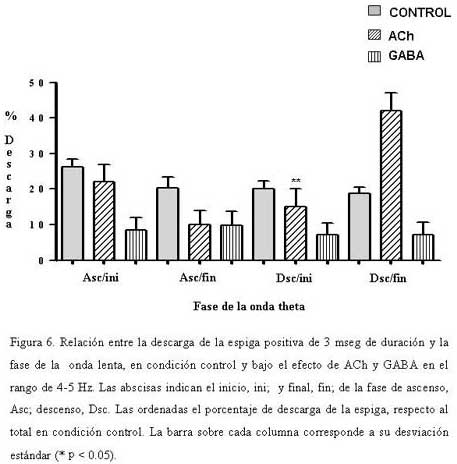
Bioanálisis Fecha de recepción: 03/06/2007 Code Number: va07004 En el hipocampo la probabilidad de descarga tanto de las células piramidales como de las interneuronas parece estar modulada por la fase del ritmo theta. En el presente trabajo se estudiaron, los cambios en la relación temporal entre la descarga neuronal unitaria, y la fase del ritmo theta, inducidos por acetilcolina (ACh), Ácido Gamma-Amino-Butírico (GABA) en la región CA1 del hipocampo, en ratas anestesiadas con uretano. La actividad neuronal se agrupó en función del sentido de la descarga de la espiga y su duración, en espigas negativas y espigas positivas de 1 mseg, 2 mseg, y 3 mseg. La ACh incrementó la actividad en un grupo de células, mientras que en otro la redujo. Ambos cambios de actividad se acompañaron por una modificación en su relación de fase con la onda theta, en forma específica para cada espiga y a una frecuencia y fase particular. El incremento de la actividad gabaérgica, si bien, modificó la relación de fase de la espiga negativa de 1 mseg, parece tener un efecto mucho más importante sobre la descarga neuronal. Los cambios inducidos tanto por la ACh como por el GABA sobre la descarga neuronal y su relación de fase con la onda theta, parecen ser específicos para cada espiga e independientes de la frecuencia del ritmo theta. Palabras Claves: hipocampo, ritmo theta, actividad unitaria, Ach, GABA Abstract Firing of pyramidal and interneuronal cells in hippocampus is modulated by theta rhythm. We decided to study the temporal relation between unit firing and theta rhythm in hippocampus CA1 induced by acetylcholine (ACh) and gamma-aminobutyric acid (GABA) in rats under urethane anesthesia. Neuronal discharges were classified according to the deflection of the spike as positive or negative and their duration in 1 msec, 2 msec and 3 msec respectively. Ach incremented the firing in some cells and decremented firing in other ones. These changes were correlated with a determined phase of theta rhythm, being specific for each type and frequency of spikes. GABA induced a generalized increment in neuronal firing and a modification in the phase relation of the negative spike form of 1 msec in duration. Ach and GABA effects on neuronal firings and its relation with theta rhythm seems to be specific for each characterized wave spike form, with no relation with theta frequency. Key Word: hippocampus, theta rhythm, unitary activity, Ach, GABA Estudio Electrofisiológico de la relación temporal entre la onda Theta y la actividad neuronal unitaria en el hipocampo de ratas Introducción La capacidad de ciertos grupos de neuronas de producir un patrón de descarga, con cierta periodicidad, representa una manera de determinar la excitabilidad organizada y secuencial de redes neuronales. La actividad sincronizada de una población neuronal podría ser la consecuencia de un sistema oscilador, que generaría un patrón de actividad rítmico, a una frecuencia relativamente constante. Uno de estos patrones es la actividad eléctrica rítmica que se registra en varias regiones del hipocampo, constituida por oscilaciones lentas periódicas a una frecuencia de 4 a 12 Hz, el ritmo theta (11) o actividad rítmica lenta (ARL). Esta actividad rítmica theta está determinada por el patrón de descarga de las células del septum medial (5,21). El ritmo theta se ha podido diferenciar en dos tipos sobre la base de su sensibilidad a la atropina; uno sensible a la atropina en el rango de frecuencia de 4–7 Hz, que puede ser registrado en animales anestesiados con uretano. El otro resistente a la atropina, está en el rango de 7–12 Hz, es eliminado selectivamente por el uretano (5,14,15). Se considera que el ritmo theta sensible a atropina es originado por un componente colinérgico de fibras ascendentes del sistema septo-hipocampal. (15,17,32), mientras que el ritmo theta residual, resistente a grandes dosis de atropina, podría deberse a proyecciones GABAérgicas (10,12,19,25,26,29), probablemente originadas en la corteza entorrinal (1,16,30,31). El ritmo theta muestra una clara relación con la conducta de los individuos y la descarga neuronal entre y a través de varias regiones del hipocampo. La probabilidad de descarga tanto de las células piramidales como de las interneuronas varía como una función del ritmo theta (5,8,22,23). Se ha demostrado en ratas despiertas, que las células piramidales y las interneuronas de todas las sub-regiones del hipocampo descargan con mayor probabilidad cerca del pico positivo de la onda theta, mientras que en animales anestesiados con uretano la probabilidad de descarga para las interneuronas en CA1 es cerca del pico negativo de la onda theta y la actividad de las células piramidales se distribuye alrededor del pico positivo (9). Sin embargo el uretano no parece modificar la frecuencia del ritmo theta sensible a atropina (5). Por otra parte, se ha observado que las “células de lugar”, aquellas células que modifican su actividad cuando el animal cambia de lugar, descargan en una fase particular de la onda theta en el rango de frecuencia de 7-12 Hz (18). El estudio de la relación entre el ritmo theta y la descarga unitaria, en ratas entrenadas, mostró que la mayoría de células piramidales e interneuronas tienen preferencia por descargar en la fase negativa de la onda theta (7). El ritmo theta parece coordinar la descarga neuronal. Ella actuaría como el control operacional del hipocampo, por lo que cualquier cambio en esta actividad oscilatoria, bien por interferir su ritmo y/o con sus corrientes generadoras, debería alterar la relación entre la descarga neuronal unitaria y la fase de la onda theta. En este trabajo se estudiaron los cambios en la relación temporal entre la descarga neuronal unitaria y la fase del ritmo theta, inducidos por la inyección local de acetilcolina (ACh), Ácido Gamma-Amino-Butírico (GABA), por medio de registros electrofisiológicos convencionales en la región CA1 del hipocampo, en ratas anestesiadas con uretano. Materiales y métodos Los sujetos experimentales fueron 14 ratas machos de la cepa Sprague Dawley de peso entre 280-300 gramos, mantenidas bajo un ciclo luz:oscuridad de 12:12 horas y alimentados ad libitum. Los animales fueron anestesiados con una solución de uretano (Ethyl Carbamate), a una dosis de 1 gr./Kgr de peso, vía intraperitoneal. El animal profundamente anestesiado se colocó en un aparato estereotáxico luego se procedió a realizar una craneotomía de acuerdo a las coordenadas obtenidas del atlas para el cerebro de ratas (20), usando como punto de referencia el Bregma (ver mas adelante coordenadas). Con ayuda de un micromanipulador se colocó un microelectrodo de metal (ø 10 μm) adosado a una microcánula de inyección de acero (ø 100 μm) de forma tal, que la punta del microelectrodo se encontraba de 2 a 3 μm por delante del orificio de salida de la microcánula, en la región CA1 en el hipocampo dorsal de acuerdo a las siguientes coordenadas: AP = 3,5 – 4,3; L = 2,5; DV = 1,8 – 2. Sistema de Registro y Adquisición: Los microelectrodos de Tungsteno (W.P.I.®) de 1 MΩ de resistencia, fueron conectados a un pre-amplificador (Mentor® N-950). La señal obtenida en forma independiente de cada hipocampo, fue amplificada por medio de un amplificador (Tektronix®, AM502), y registrada en forma continua a través de una tarjeta genérica de sonido de dos canales con una velocidad de muestreo de 48 Ks/s conectada a una computadora Pentium que disponía de un programa editor de señales de audio ( Syntrillium CoolEdit Pro 1.5). Las señales tanto del ritmo theta como de las descargas neuronales fueron digitalizadas y grabadas directamente en el disco duro para su análisis posterior. Típicamente se registró de 1 a 3 minutos de actividad basal, luego se aplicó la droga por vía de la cánula intracraneal y se mantuvo el registro hasta que la actividad neuronal retornaba a su condición basal. El sitio de registro electrofisiológico y de aplicación de la droga fue corroborado mediante lesión electrolítica al final del experimento, que luego se evidenció en cortes histológicos observados bajo microscopio de luz. Inyección de las Drogas: Todas las drogas fueron preparadas en solución fisiológica 0,9 % NaCl, manteniéndose la isosmolaridad de las soluciones finales.: solución de sulfato de Atropina (Sigma); 0,5 mgrs/mL; solución de Acido Gamma-Amino-Butírico (Sigma), GABA; 0,5 grs/mL; solución de Cloruro de Acetilcolina (Sigma), ACh; 1 mgr/mL. Previo a la colocación de los microelectrodos en la zona de registro en el hipocampo, la cánula metálica y el tubo plástico que la conectaba a una inyectadora (Hamilton®) de 1 μL, fueron llenados y purgados con la solución de la droga a inyectar. Se determinó que la inyección de 0,1 mL/8-10 seg era una velocidad adecuada para garantizar la estabilidad del registro. Sin embargo la velocidad de difusión de la droga no se cuantificó. Los efectos de la droga se evaluaron en segmentos de registro obtenidos luego de terminada su aplicación. Procesamiento de la señal: Separación de la onda lenta y actividad unitaria (espigas) El programa para análisis de sonido dispone de una serie de filtros digitales y permite hacer análisis de frecuencia mediante la aplicación de la Transformada Rápida de Fourier (FFT). La señal adquirida fue inicialmente filtrada con un esquema pasa-banda de 4-10 Hz, separando de esta manera la onda lenta. Se procedió luego a formar 6 grupos por rango de frecuencia de 1 Hz de ancho de banda de 4-5, 5-6, 6-7, 7-8, 8-9 y de 9-10 Hz. Para este procedimiento se tomó en cuenta la frecuencia que presentaba la mayor amplitud de onda determinada por el análisis de frecuencia para cada rango. La actividad unitaria se aisló del mismo segmento de registro aplicándole un filtro pasa banda de 0.1 a 5 KHz. Caracterización de la actividad unitaria Se utilizaron tres criterios independientes para caracterizar la actividad unitaria. En primer lugar, por el sentido de deflexión inicial de la espiga; en espigas de descarga negativa y espigas de descarga positiva. Además cada espiga se discriminó por su amplitud y su duración se determinó visualmente en forma directa sobre el monitor mediante el uso de cursores de tiempo, fijando un nivel de referencia en el inicio de la primera deflexión de la espiga, y el final cuando la espiga alcanzaba nuevamente el nivel basal. Localización y cuantificación de la descarga neuronal unitaria en relación con la fase de la onda lenta. En todos los experimentos se evaluaron segmentos de registros de igual duración, tanto en condición control; como después de haber aplicado la droga. Se cuantificó la cantidad de espigas por regiones de cada fase de la onda theta y se determinó el porcentaje respecto al total de espigas del segmento del registro control. Las espigas aisladas se compararon con el curso temporal de la onda lenta en cada rango. Para determinar la coincidencia de la espiga con una porción de la fase de ascenso o de descenso, se tomó como referencia una línea que dividía la onda lenta en dos partes, de tal manera que podíamos subdividir, a la fase ascenso en una porción que llamamos inicial (Asc/ini) localizada por debajo de la línea, y una parte superior o final (Asc/fin) por encima de la línea. La parte superior de la fase de descenso, la llamamos inicial (Dsc/ini) y su parte inferior, final (Dsc/fin). Visualmente se determinó para cada espiga, con cual parte o porción de cada fase de la onda lenta coincidía la actividad neuronal unitaria. No se tomaron en cuenta aquellas espigas que coincidían con el valle o el pico de la onda, ya que no se pudo determinar sí correspondían a la fase de ascenso o de descenso de la onda theta. Análisis estadístico. Los datos fueron expresados antes y después de haber aplicar la droga, como porcentajes de descarga de cada espiga, respecto al total en el segmento considerado control. La media y la desviación estándar de los porcentajes fueron cuantificados y graficados utilizando el programa GraphPad Prism 4. Los datos obtenidos se compararon aplicando la prueba estadística “t de Student” para muestras no pareadas. Se consideraron estadísticamente diferentes cuando el nivel de significancia (p) fue menor de 0,05. En todos los rangos de frecuencia de la onda theta, se realizaron dos tipos de prueba tanto en condición control, como bajo el efecto de la droga; en una se comparó la media de los porcentajes de descarga en la fase de ascenso, con los de la fase de descenso; en la otra la descarga de la espiga en la porción inicial de la fase se comparó con la descarga en su porción final. Resultados Determinación de la relación de fase entre la onda lenta y la descarga neuronal unitaria: En todos los registros obtenidos de la región CA1 de cada hipocampo, se aisló tanto la onda lenta como las espigas correspondientes para cada rango de frecuencia. La Figura 1 muestra a modo de ejemplo, dos pares de segmentos de registros simultáneos, de ondas lentas de 4-5 Hz, y las espigas correspondientes, una cuya primera deflexión de la espiga es positiva (A) y la otra predominantemente negativa (B). La proyección de las espigas (líneas punteadas) sobre la onda lenta permitió determinar la porción de la fase de la onda theta con la cual coincidía la espiga. Tanto en A como en B, los números entre paréntesis señalan la porción de la fase relacionada con la espiga: (1) porción inicial (ini), (2) porción final (fin). Caracterización de la espiga: En función del sentido de la descarga, la actividad neuronal se agrupó en dos poblaciones, una de espigas negativas y otra de espigas positivas. Luego se procedió formar tres subgrupos de espigas, en función de su duración de 1 mseg; 2 mseg; 3 mseg. La Figura 2 muestra el registro ampliado de la forma de onda de dos espigas negativas (A-B) y positivas (C-D), así como el inicio y el final de la onda donde se determinó la duración de cada espiga. Efecto de la ACh, y el GABA sobre la Onda Theta: El efecto de la ACh sobre la onda theta se pudo evaluar en 16 experimentos. El análisis de frecuencia permitió evidenciar un incremento significativo (p < 0.05) de la frecuencia de la onda theta en los siguientes rangos de 5-6 Hz de 5,35±0,06 a 5.6±0,05 Hz, mientras que de 7-8 Hz, la frecuencia paso de 7,4±0,05 a 7,6±0.06 Hz. Sin embargo la ACh no modificó la actividad theta en los otros rangos evaluados. El efecto del GABA se cuantificó en 12 experimentos, los cambios producidos sobre la onda theta fueron diferentes a los inducidos por la ACh. EL GABA incrementó significativamente (p < 0.05) la frecuencia de la onda theta en los siguientes rangos de 4 -5 Hz la frecuencia pasó de 4,3±0,08 a 4,7±0,07 Hz; de 5 – 6 Hz se incrementó de 5.38± a 5.5± Hz; en el rango de 6 –7 Hz de 6.36± a 6.5± Hz, mientras que de 7 – 8 Hz de 7.3± a 7.5± sin afectar las frecuencias de 8 a 10 Hz. Efecto de ACh y GABA sobre la relación de fase entre la onda lenta y la descarga neuronal unitaria: Espiga Negativa de 1 mseg.: Se evaluaron 7 casos de neuronas que descargaban este tipo de espiga. En condiciones control la frecuencia promedio de descarga fue de 2 esp./seg y su relación de fase con la onda lenta mostró diferencias únicamente en la fase de descenso del rango de 5-6 Hz (Fig. 3B). En este rango la espiga descargó significativamente menos en la porción inicial (Dsc/ini) de la fase de descenso, en comparación con su porción final (Dsc/fin). La aplicación local de ACh incrementó ligeramente la frecuencia de descarga de esta espiga, y provocó que descargase en igual proporción en ambas fases de la onda theta en el rango de 5-6 Hz. El efecto del GABA se pudo evaluar en 6 células que descargaban espigas de este tipo, la frecuencia de descarga se redujo en promedio a 0.61 esp/seg (* p < 0.05). Por otra parte modificó la relación entre la descarga de la espiga y la fase de ascenso y descenso de la onda theta. En el rango de 4-5 Hz (Fig 3A), la espiga descargó significativamente (* p < 0.05) menos tanto en la porción final de la fase de ascenso (Asc/fin) como de descenso (Dsc/fin) de la onda theta en relación con su porción inicial (Asc/ini y Dsc/ini). Además modificó la relación de fase en el rango de 5-6 Hz (Fig. 3B), en este caso la espiga descargó en igual proporción en ambas fases de la onda theta. Adicional a estos cambios el GABA provocó que la espiga descargase significativamente (* p < 0.05) en menor proporción en la porción inicial de la fase de ascenso (Asc/ini) en relación con la porción final (Asc/fin), mientras que descargó significativamente (* p < 0.05) menos en la porción final de la fase de descenso (Dsc/fin) respecto a su porción inicial (Dsc/ini) en el rango de 7-8 Hz de la onda theta (Fig. 3C). Espiga Negativa de 2 mseg. En 7 células que descargaban este tipo de espiga, la frecuencia en condición control fue de 1.4 esp/seg, y su relación de fase con la onda theta en los rangos de 6-7 Hz y de 7-8 Hz (Fig. 4), mostró diferencias significativas (* p < 0.05), la espiga descargó en menor proporción en la porción final de la fase de ascenso (Asc/fin) en relación con su porción inicial (Asc/ini). La ACh incrementó la frecuencia de descarga de esta espiga a 2.5 esp/seg, y modificó su relación de fase con la theta. En el rango de 6-7 Hz (Fig. 4A), la célula redujo su descarga en la porción final de fase de ascenso (Asc/fin) en relación con su porción inicial (Asc/ini), mientras que la incrementó en la porción final de la fase de descenso (Dsc/fin) en comparación con su porción inicial (Dsc/ini). Sin embargo, en el rango de 7-8 Hz, la ACh provocó que la espiga descargase en igual proporción en ambas fases de la onda theta (Fig. 4B). El GABA redujo significativamente la frecuencia de descarga de las neuronas que generaban este tipo de espiga a 0.14 esp/seg (* p < 0.05) por lo cual, los cambios en la relación entre la descarga neuronal y la fase de la onda lenta no se pudieron cuantificar. Espiga Positiva de 2 mseg. Se evaluaron 6 neuronas que descargaban espigas positivas de 2 mseg de duración. En condición control la frecuencia promedio de descarga fue de 1.6 esp./seg. El análisis de relación de fase mostró que descargaba en menor proporción en la fase de descenso en casi todos los rangos de la onda lenta. Además en el rango de 6-7 Hz (Fig. 5), descargó a su vez en menor proporción en la porción final de esta fase (Dsc/fin), en comparación con su porción inicial (Dsc/ini). La aplicación local de ACh incrementó su frecuencia de descarga a 3.5 esp/seg. La relación de fase con la onda lenta mostró un comportamiento opuesto al observado en condición control. El análisis de la descarga de la espiga dentro de una misma fase, mostró que en el rango de 6-7 Hz de onda theta, la espiga descargó significativamente menos en la porción final de la fase de ascenso (Asc/fin) respecto a su porción inicial (Fig. 5). El efecto del GABA se pudo evaluar en 5 células en las cuales se registró este tipo de espigas. Su frecuencia de descarga se redujo en forma significativa a 0.02 esp/seg (* p < 0.05). Espiga Positiva de 3 mseg. En 12 neuronas que descargaban este tipo de espigas, su frecuencia de descarga en condición control fue de 2.1 esp./seg. Por otra parte el análisis de su relación con la onda theta mostró que la espiga descargaba en igual proporción tanto en la porción inicial como en la final de cada fase. El efecto de la ACh se pudo evaluar en 6 células. Su frecuencia de descarga se redujo a 1.4 esp/seg., y descargó significativamente en mayor proporción en la porción final de la fase de descenso (Dsc/fin) de la onda lenta en comparación con su porción inicial (Dsc/ini) el rango de 4-5 Hz. En 6 neuronas en las cuales se evaluó el efecto del GABA, la descarga de esta espiga se redujo a 1.22 esp/seg, sin modificar su relación de fase con la onda theta (Fig. 6). Discusión La ACh y el GABA incrementaron la actividad de la onda theta en el rango 4-7 Hz, que se corresponde con el rango de frecuencia de la onda theta atropina sensible que no es afectada por el uretano, y concuerda en parte con resultados reportados (13,17), en los que se demuestra por medio de estudios de correlación entre registros electrofisiológicos y muestreo de ACh por microdiálisis en el hipocampo de ratas anestesiadas con uretano, que incrementos en amplitud y frecuencia de la onda theta, se correlacionan positivamente con la liberación de ACh. Este aumento en la frecuencia del ritmo theta, estaría de acuerdo con la idea de la coexistencia de dos tipos de ritmos thetas independientes, diferentes en su origen, frecuencia y naturaleza neuroquímica. Un ritmo theta que bajo el efecto de uretano es sensible a la atropina, y depende de la proyección colinérgica septo-hipocampo y Gabaérgica de la banda diagonal de Broca, mientras que el de alta frecuencia (7-12 Hz) sería sensible a uretano y dependería de otras estructuras cerebrales (2,4,5,14,15,26). El análisis de la relación entre la descarga neuronal unitaria y la fase de la onda lenta, mostró cambios específicos tanto en los rangos en que se modificó el ritmo theta, como aquellos en los que no se alteró. La ACh provocó dos efectos en la descarga neuronal unitaria. En un grupo de células incrementó su actividad mientras que en otro la redujo. Ambos cambios de actividad se acompañaron por una modificación en su relación de fase con la onda theta, en forma específica para cada espiga y a una frecuencia y fase particular. El incremento de la actividad Gabaérgica, si bien modifica la relación de fase de la espiga negativa de 1 mseg, parece tener un efecto mucho más importante sobre la descarga neuronal. Este efecto de GABA sobre la actividad neuronal, confirma en parte resultados reportados (28), en los que demuestran que la aplicación de diazepan un inhibidor postsináptico mediado por GABA, en la región CA1 del hipocampo, reduce la actividad neuronal espontánea. Estos hallazgos sugieren que, al menos en ratas anestesiadas con uretano, la modulación de la frecuencia de la onda theta dentro del rango de 4-7 Hz y la descarga neuronal unitaria, parecen estar mediadas en parte por un mecanismo en el que participa la ACh y el GABA. Por otra parte los cambios observados en la relación entre la descarga neuronal y la fase del ritmo theta, muestran que incrementos en la concentración de uno de estos neurotransmisores modifica en forma específica la probabilidad de la descarga neuronal unitaria, en relación con la fase de la onda theta en ciertas frecuencia. Los cambios en la relación de fase no parecen estar determinados por incrementos en la frecuencia del ritmo theta, y pueden darse de dos maneras: una entre la fase de ascenso y la fase de descenso, y la otra dentro de una misma fase. Estos resultados sugieren que la actividad neuronal unitaria guarda una relación específica con la fase de la onda theta en cada rango de frecuencia. Estas observaciones, están de acuerdo con la coexistencia de dos tipos de codificación de la información a través de las redes neurales. Uno basado en un código de frecuencia, en el que la información estaría implícita en la frecuencia de la descarga neuronal (24). El otro procesamiento de la información estaría fundamentado en un código temporal, en el cual la oscilación theta funcionaría como un sincronizador, que permitiría coordinar selectivamente la descarga neuronal en el hipocampo. (3,7,23,22).Agradecemos al Dr. Antonio Eblen la lectura crítica y sus valiosos comentarios sobre este manuscrito. Este trabajo fue parcialmente financiado por la Coordinación de Investigación de la Facultad de Medicina y el CDCH de la Universidad Central de Venezuela. (# 09-184-2005) Referencias
Copyright 2007 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela The following images related to this document are available:Photo images[va07004f2.jpg] [va07004f4.jpg] [va07004f6.jpg] [va07004f3.jpg] [va07004f1.jpg] [va07004f5.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}