 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
VITAE Academia Biomédica Digital, Número 30, Enero - Marzo, 2007 Parasitología Ultraestructura de Blastocystis hominis y su enquistamiento en cultivo polixénico. Blastocystis hominis ultrastructure and encystment in polixenic culture. 1Carmen Guzmán de Rondón, 2Héctor Arrechedera, 3Eva Pérez de Suarez 1guzmanca@ucv.ve
Parasitología
*Laboratorio de Amibiasis, Cátedra de
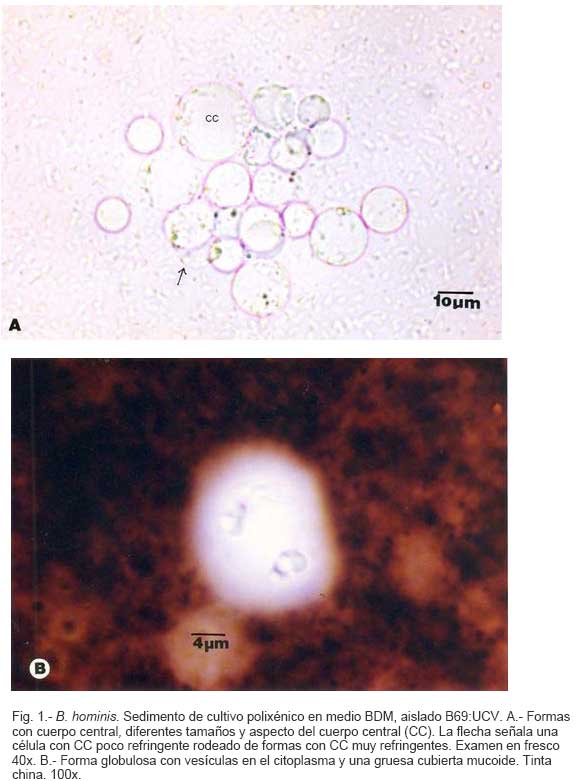
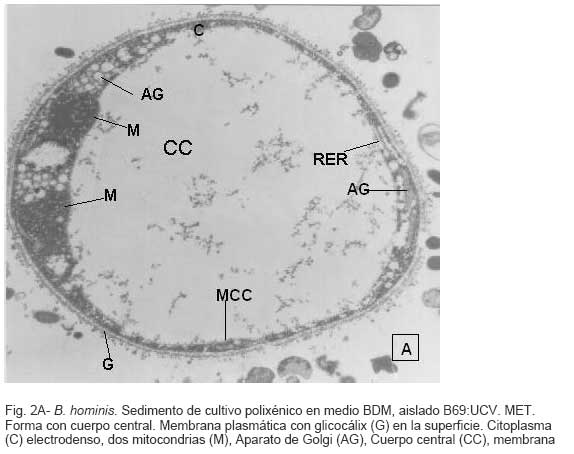
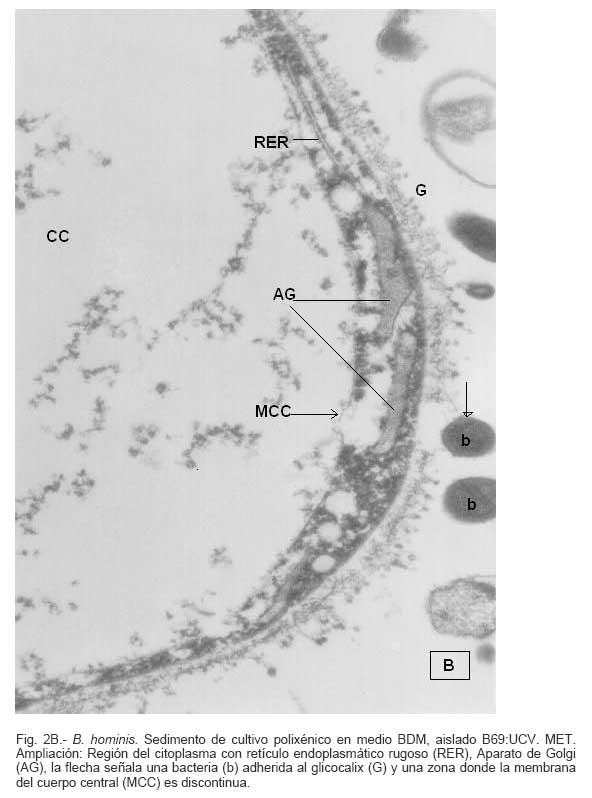
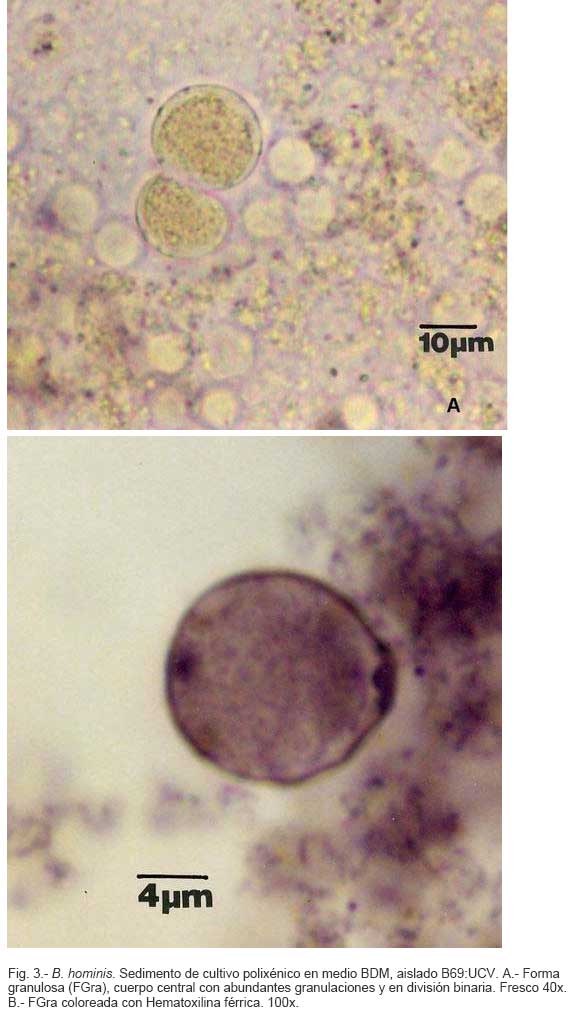
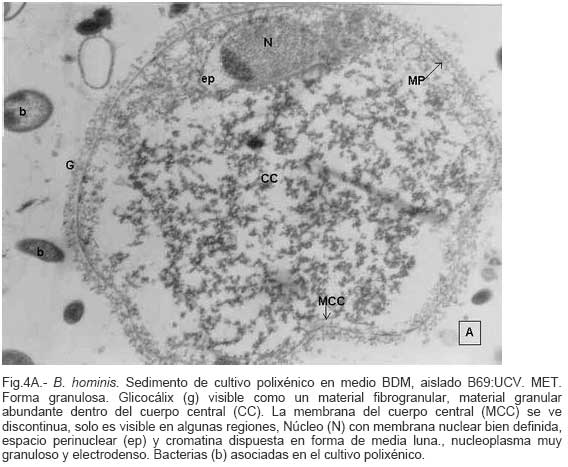
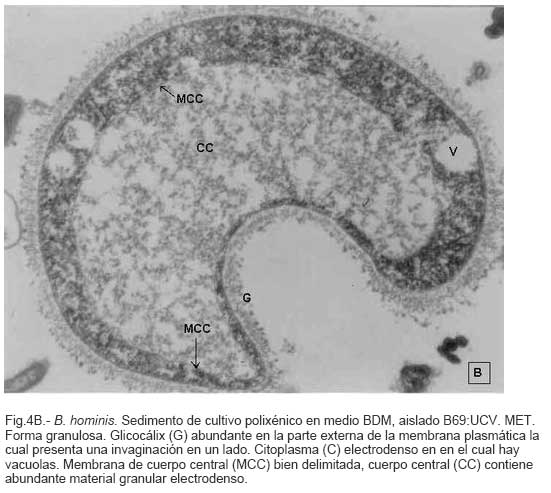
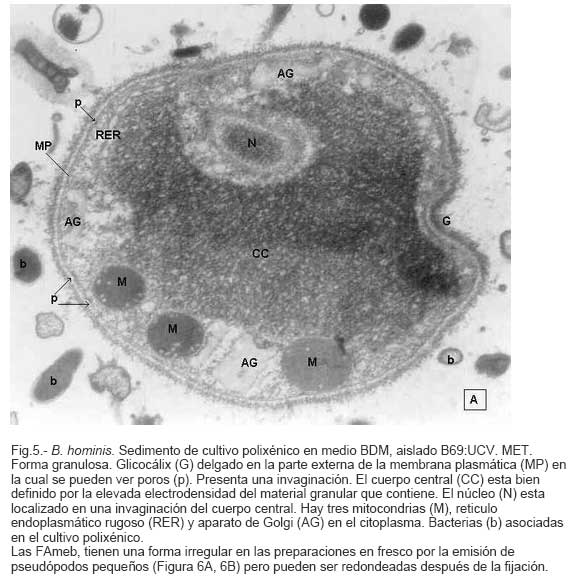
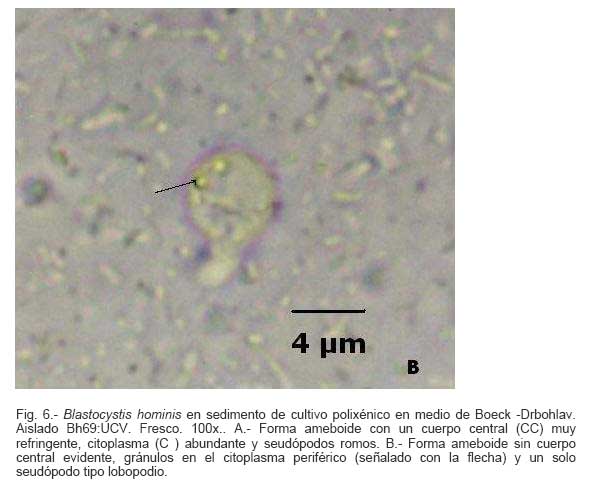
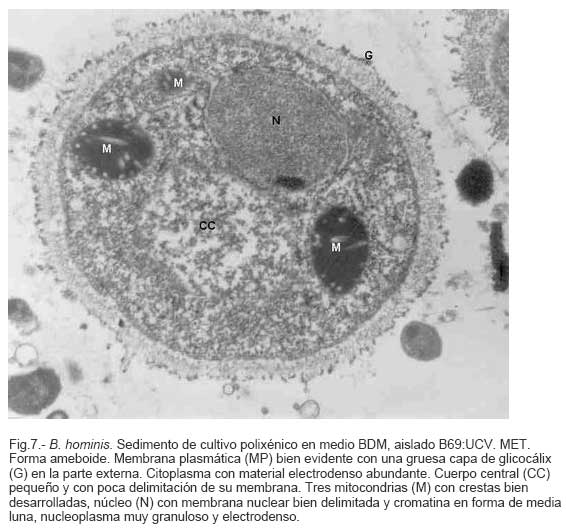
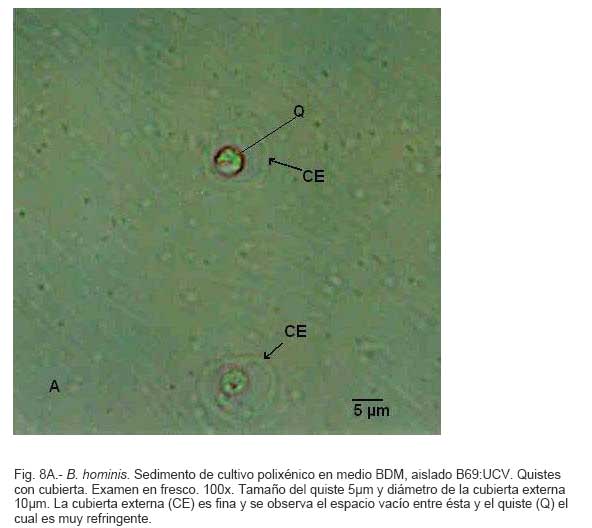
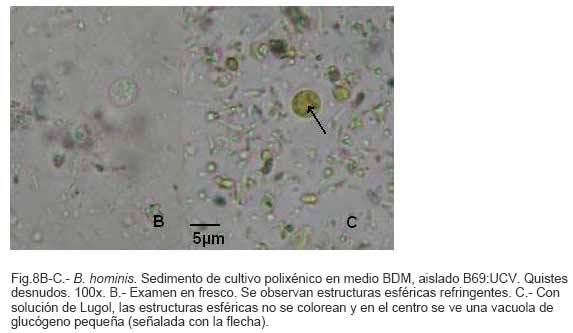
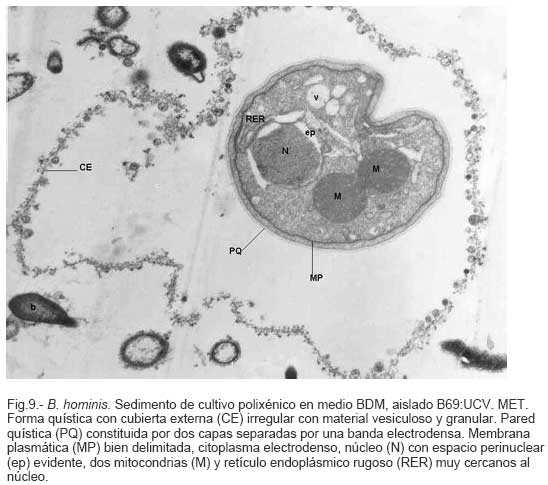
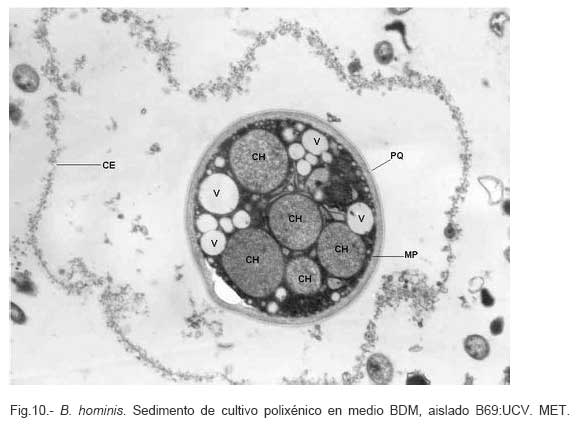
Parasitología, Escuela de Bioanálisis Fecha de recepción: 11/05/2007 Code Number: va07006 Blastocystis hominis (Bh) es un organismo, cuya taxonomía, ciclo biológico y forma de transmisión ha generado polémica. Han sido descritas una variedad de formas por microscopía óptica (MO) y microscopía electrónica de transmisión (MET) y las características de las formas reportadas como quistes en MET no son uniformes. El conocimiento de todas las variantes morfológicas es indispensable para la identificación y para los estudios biológicos. La caracterización morfológica de B. hominis fue realizada en el aislado Bh 69: UCV, obtenido a partir del cultivo original de las heces de un paciente con sintomatología intestinal y mantenido en el laboratorio de Amibiasis en cultivo polixénico en medio Boeck-Drbohlav (mod) (BDM). Las formas vegetativas del parásito, forma con cuerpo central (FCC), forma granulosa (FGra), Forma ameboide (FAmeb), y los quistes que aparecieron de manera espontánea en el cultivo, son descritas al microscopio de luz (400x y 1000x) y MET según las técnicas convencionales. Característicamente todas las formas vegetativas están rodeadas por una cubierta mucoide y presentan un cuerpo central, que ocupa la mayor parte de la célula y delimita un delgado anillo citoplasmático, en el cual los organelos celulares son observados como inclusiones granulares refringentes. Aparecen dos tipos de quistes, los desnudos de forma redonda u oval, muy refringentes, con 5-6 µm de diámetro y otros con una cubierta externa muy delgada, separada por un espacio de 2-3 µm. Mediante MET en el citoplasma de todas las formas, puede describirse el núcleo con cromatina dispuesta en forma de media luna, mitocondrias redondeadas con pocas crestas, numerosas vacuolas digestivas, aparato de Golgi, RER y gran cantidad de ribosomas y el glicocálix sobre la membrana plasmática. La pared quística presenta tres zonas definidas y la cubierta externa está constituida por material fibrogranular y vesiculoso. Algunos quistes presentaron estructuras esféricas cuya morfología no corresponde a la de mitocondrias ni núcleos y que podrían ser células hijas. Las características morfológicas descritas, coinciden con las reportadas previamente para las formas vegetativas y los quistes y sugerimos una posible secuencia en la transformación de las formas vegetativas a formas de resistencia. Palabras Claves: Blastocystis, protista, quistes Abstract Blastocystis hominis (Bh) is an organism, whose taxonomy, biological cycle and mode of transmission is controversial. A variety of forms have been described using ligth and transmission electron microscopy (TEM) and there is not uniformity in reporting cysts characteristics in TEM. The knowledge of the morphology of Blastocystis hominis is indispensable for identification and biological studies. Morphologic characterization was made in B. hominis, isolated Bh69:UCV, obtained from the original culture from a patient's faeces with intestinal symptomatology and maintained in the Amibiasis laboratory in polixenic culture in Boeck-Drbohlav (modified) media (BDM). The vegetative forms of the parasite, central body form (CBF), granular form (GF), amoeboid form (AF), and cysts which spontaneously appeared on the cultures, are described, using light microscope (400x and 1000x) and TEM, according to conventional techniques. All the vegetative forms are characteristically surrounded by a mucoid cover, and have a central body that occupies most of cell and delimits a thin cytoplasmic ring, in which the cellular organelles are observed like refractile inclusions. There are two types of cysts, naked cyst are spherical to ovoid, refractile, 5-6 µm of diameter and cyst with external cover, separated by a space of 2-3 µm. By TEM, in the cytoplasm of all the forms, a nucleus with chromatin in crescent-shaped form, spherical mitochondria with few crests, digestive vacuoles, Golgi, RER and great amount of ribosomes and glicocálix on the plasmatic membrane were observed. Cyst wall showed three defined zones, with the external cover made of fibrogranular and vesicular material. Some cysts presented spherical structures whose morphology does not correspond with mitochondrias or nuclei and they could be daughters cells. The morphologic characteristics agree with previously reported for the vegetative forms and cysts, and we suggest a possible sequence in vegetative forms transformation to resistance forms Key Word: Blastocystis, protist, cysts Ultraestructura de Blastocystis hominis y su enquistamiento en cultivo polixénico. Introducción Blastocystis hominis es un organismo anteriormente ubicado en el Reino Protista, Subreino Protozoa (Zierdt, 1988), recientemente fue reconocida su relación filogenética con otros protistas Stramenopiles (Silberman et al. 1996, Arisue et al., 2002). Infecta con elevada frecuencia a individuos de todas las edades, con sintomatología gastrointestinal y sin asociación demostrada con otro patógeno, también se observa en personas asintomáticas (García et al.1984; Diaczok and Rival, 1987; Gallager and Vanglarsick, 1987; El Marsry et al., 1988; Guglielmenti et al, 1989; Bratt and Tikasingh, 1990; Jeddy and Farrington, 1991; Pérez de Suárez et al., 1997, Stark et al, 2007). Es polimórfico y hasta hace pocos años se pensaba que B. hominis solo presentaba formas vegetativas, las cuales eran nombradas de acuerdo a su aspecto morfológico, como forma con cuerpo central (FCC), forma granulosa (FGra) y forma ameboide (FAmeb) (Zierdt et al., 1967; Boreham and Stenzel, 1993). Pérez de Suárez y Guzmán de Rondón en 1994, describieron una variante morfológica que presenta abundantes vesículas lipídicas en el citoplasma y la denominaron forma globulosa (FGlob). Zierdt et al., en 1967, proponen que la forma granular podría ser el estado quístico, pero es Melhorn, quien en 1988, describió por primera vez la ultraestructura de los quistes de B. hominis en heces y fue corroborado en estudios posteriores (Stenzel and Boreham, 1991, Zaman 1995, Chen, 1999) y se ha reportado el enquistamiento in vitro (Suresh et al., 1993, Guzmán de Rondón y col, 1996, Init et al. 2003). Existen reportes sobre infección experimental en ratas por vía intracecal con formas de cultivo y por vía oral con quistes (Suresh et al., 1993, Yoshikawa et al. 2004). El ciclo evolutivo de B. hominis ha sido objeto de especulaciones y se han propuesto diferentes hipótesis al respecto (Zierdt, 1973, Boreham and Stenzel 1993, Singh et al. 1995, Tan, K. 2004). Sin embargo ninguna ha sido comprobada, ni hay detalles de la secuencia de eventos que ocurren en su evolución dentro del intestino humano. Está demostrado que puede transmitirse de un hospedador a otro por medio de quistes (Init et al. 2003) y las diferentes variantes morfológicas vistas en las muestras biológicas y en cultivos, sugieren un desarrollo del organismo pasando por diferentes fases hasta ahora no definidas. El quiste pareciera jugar un importante papel en la resistencia a las drogas ya que el enquistamiento fue inducido in vitro al contacto con el metronidazol (Init et al. 2003). El objetivo de este trabajo fue estudiar las variantes morfológicas de un aislado de Blastocystis hominis en un cultivo polixénico, haciendo énfasis en los quistes producidos espontáneamente en dicho cultivo. Materiales y métodos El aislado Bh69:UCV de B. hominis, proveniente de un paciente con sintomatología intestinal, fue obtenido en el Laboratorio de Amibiasis a partir del aislado original y mantenido en cultivo polixénico con la flora intestinal asociada, en el medio de Boeck-Drbohlav modificado (BDM), el cual es un medio bifásico, con una fase sólida constituida por huevo y sangre humana desfibrinada, que se esteriliza mediante tindalización a 80ºC, una fase líquida constituida por un Ringer, al momento de la siembra se agrega almidón de arroz estéril y se incuba a 37ºC (Pérez de Suárez, E.1986, Pérez de Suárez y Guzmán de Rondón, 1994). El cultivo fue examinado a las 24, 48 y 72 horas mediante examen microscópico del sedimento entre lámina y laminilla y el mantenimiento del aislado fue por resiembras sucesivas a un medio de cultivo fresco. Para la observación se utilizó microscopio Olympus calibrado, con ocular micrométrico para medición e identificación parasitológica, y con cámara digital adaptada para el registro fotográfico. Los sedimentos de cultivos con diferentes tiempos de incubación, desde 24 hasta 72 horas fueron fijados en Karnosky y este material fue procesado según las técnicas convencionales de microscopía electrónica de transmisión (MET) para su posterior estudio ultraestructural. Se lavó con buffer fosfato y postfijó con tetróxido de Osmio. Las células fijadas fueron deshidratadas en una serie ascendente de acetona y luego incluídas en Araldita. Los cortes fueron contrastados con acetato de Uranilo y observados en microscopio electrónico Hitachi modelo H-300, del Laboratorio de Biología Celular del Instituto de Medicina Tropical, de la Universidad Central de Venezuela. ResultadosEl examen microscópico del sedimento del cultivo polixénico de B. hominis en el medio BDM, a las 24, 48 y 72 horas de incubación reveló el marcado polimorfismo previamente descrito para este organismo, observándose las formas con cuerpo central (FCC), forma granulosa (FGra), forma ameboide (FAmeb) y los quistes, además de una marcada variación en el tamaño de 4-30 µm. Con aumentos 400x y 1000x (Figura 1A), las FCC presentan forma redondeada u ovalada, 4-30 µm de diámetro, un organelo membranoso de gran tamaño denominado cuerpo central que puede ocupar entre 40- 75% del tamaño celular y su aspecto puede ser muy refringente o traslúcido. El citoplasma es escaso, delimitado externamente por la membrana citoplasmática e internamente por la membrana del cuerpo central, queda reducido a un delgado espacio en el cual se ven las inclusiones citoplasmáticas. Sobre la membrana citoplasmática está presente una cubierta mucoide, observada en todas las variantes morfológicas y demostrable en las preparaciones con tinta china (Figura 1B). Al microscopio electrónico (Figura 2A, Figura 2B) esta cubierta externa se observa como un material fibrogranular alrededor de la membrana citoplasmática (glicocálix). El cuerpo central (CC) tiene una membrana (MCC) aparentemente discontinua en algunas zonas, y puede o no contener material electrodenso como gránulos finos. En el citoplasma pueden verse núcleos (N) con un espacio perinuclear (ep), 1-3 mitocondrias (M), Retículo Endoplásmico Rugoso (RER), Retículo Endoplásmico Liso (REL), Aparato de Golgi (AG) y ribosomas libres (R). El núcleo presenta la membrana nuclear (MN) con poros (p), característica de los eucariontes. Las mitocondrias varían en número, forma y tamaño, dentro de una misma célula y en una misma cepa. Están localizadas cerca del núcleo. Presentan forma circular, tubular o vesiculosa, con elevada electrodensidad y crestas abundantes o escasas que se ven como espacios claros. El retículo endoplásmico, presenta una estructura sacular simple o múltiple, de electrodensidad moderada y puede o no tener ribosomas adheridos. El aparato de Golgi está constituido por estructuras vesiculares múltiples, localizado muy cercano al núcleo y las mitocondrias. En el citoplasma hay vesículas libres con membrana sencilla, inclusiones electrodensas y ribosomas libres (Figura 2B). Las FGra al microscopio de luz (Figura 3A, 3B) son similares a las FCC pero tienen el cuerpo central lleno de gránulos. Al microscopio electrónico tienen material electrodenso agrupado en cúmulos o distribuidos uniformemente en todo el cuerpo central (Figura 4A, Figura 4B, Figura 5A). Estas formas al igual que las FCC pueden verse deformadas con invaginaciones que también se reflejan en el cuerpo central. En algunas células el cuerpo central presenta mayor electrodensidad (Figura 5A). En el citoplasma se ve el núcleo con su nucleolo característico en forma de media luna, las mitocondrias, ribosomas, RER, y vacuolas. Presenta un glicocálix de grosor variable sobre la membrana citoplasmática (Figura 4A, Figura 4B, Figura 5A). Las FAmeb, tienen una forma irregular en las preparaciones en fresco por la emisión de pseudópodos pequeños (Figura 6A, B) pero pueden ser redondeadas después de la fijación. El cuerpo central generalmente es más pequeño que en las otras formas (Pérez de Suárez y col, 1994, Guzmán de Rondón, 1995) y sobre la membrana plasmática también esta presente la cubierta mucoide. Al microscopio electrónico (Figura 7) la membrana plasmática está bien delimitada y el glicocálix es una capa gruesa de material fibrogranular dispuesto de manera radiada. La membrana del cuerpo central no está bien definida en algunas células, pero puede diferenciarse el material interno del resto del contenido citoplasmático. El citoplasma contiene abundante material fibrogranular grueso y hay un aumento de electrodensidad en la parte externa de la membrana plasmática. El núcleo presenta las características ya descritas, pero puede observarse ligeramente de mayor tamaño. Las mitocondrias pueden presentar abundantes crestas (Figura 7). Los quistes pueden presentar una cubierta externa muy delgada (Figura 8A), otros no la tienen y pueden denominarse quistes desnudos (Figura 8B,C). Los quistes con cubierta presentan una delicada capa externa separada de la pared quística por un espacio claro, el cual es casi imperceptible en el examen al microscopio de luz. El diámetro de la cubierta externa es de 10 µm, mientras que el quiste verdadero solo alcanza 4-5 µm de diámetro. Este tiene forma redondeada u oval, en el sedimento se ven muy refringentes, verdosos y se les distingue muy bien la pared gruesa, algunas veces estructuras internas. Pueden presentar una retracción de la pared quística y verse de manera similar al aspecto que adquieren los quistes de otros protozoarios intestinales (Figura 8A). Los quistes desnudos, tienen iguales características que los anteriores, pero carecen de la cubierta externa (Figura 8B, C). En las preparaciones teñidas con lugol se destacan en el interior cierta cantidad de estructuras esféricas prominentes, entre las cuales pudiera encontrarse el núcleo, pero con esta coloración es difícil identificarlo. Además pueden verse pequeñas vacuolas de glucógeno (Figura 8C). La cubierta externa de los quistes, ultraestructuralmente se observa constituida por material fibrogranular y dentro está el quiste propiamente dicho, cuya pared presenta de afuera hacia dentro una capa externa, fina y filamentosa, una banda intermedia electrón opaca y una capa interna homogénea, en contacto con la membrana celular (Figura 9). Hay quistes con organelos citoplasmáticos similares a las formas vegetativas, con el núcleo rodeado del espacio perinuclear y la cromatina dispuesta en la periferia, las mitocondrias redondeadas con pocas crestas, AG, RER y gran cantidad de ribosomas libres (Figura 9). Otros quistes presentan gran cantidad de gránulos electrodensos, vesículas electrontransparentes de forma circular y con diferente tamaño y cinco estructuras esféricas con membrana propia, cuya morfología no corresponde a núcleos ni mitocondrias (Figura 10). Forma quística. Cubierta externa (CE) irregular constituida por material fibrogranular. Pared quística (PQ), gruesa con tres zonas bien definidas. La membrana plasmática no se identifica claramente por la elevada cantidad de gránulos electrodensos que hay en el citoplasma. El quiste presenta vesículas (V) vacías y cinco estructuras esféricas con membrana propia semejantes a células hijas (CH). Discusión Las características ultraestructurales de las FCC, FGra y FAmeb de B. hominis encontradas en el presente estudio coinciden con los reportes de Zierdt et al. (1967), Zierdt, (1973, 1988, 1991); Dunn et al., (1989); Stenzel and Boreham, (1991); Boreham and Stenzel, (1993); Matsumoto et al., (1987), corresponden a células activas, que presentan membrana celular, en el citoplasma se observan organelos como el núcleo, mitocondrias, RER, AG, ribosomas, vacuolas y se reproducen por división binaria. En el cultivo polixénico en medio BDM, ocurrió el enquistamiento espontáneo de B. hominis aislado Bh69:UCV entre las 24 y 72 horas de cultivo y los quistes presentaron la morfología previamente reportada (Melhorn, 1988, Stenzel and Boreham, 1991; Boreham and Stenzel, 1993, Zaman et al. 1995, 1997). La observación de los quistes en este medio de cultivo nos permitió identificar posteriormente estas formas quísticas en las heces de los pacientes. En la forma quística aquí descrita, la pared tiene características ultraestructurales similares a la de otros protozoarios intestinales, aunque presenta algunas diferencias, como la presencia de una cubierta externa fibrilar. Esta pared cumpliría la función de resistencia, y confiere protección a la membrana plasmática y a todos los organelos presentes en el citoplasma, coincidiendo con las descripciones hechas por Melhorn, (1988), Stenzel and Boreham, (1991), Boreham and Stenzel, (1993), Zaman et al., (1997). Con la descripción de esta forma, queda establecida la posibilidad de una vía de transmisión similar a la de otros protozoarios intestinales parásitos del hombre. Esto ha sido confirmado por Suresh et al. (1993), quien indujo el enquistamiento in vitro y logró infectar ratas Wistar por vía oral, con estructuras que morfológicamente correspondían con los quistes descritos por Stenzel and Boreham, (1991), mientras que no lograron la infección por esta misma vía usando FCC. La transmisión a través de los quistes obtenidos de heces también ha sido comprobada, por infecciones experimentales en ratas (Moe et al, 1997, Yoshikawa et. al, 2004). Estos resultados determinan una diferencia biológica entre las FCC y las FQ y revela la existencia de formas de resistencia que corresponderían a un organismo del reino protista. Respecto a la formación del quiste, Boreham and Stenzel, (1991), presentan observaciones de estructuras semejantes a quistes que pudieran originarse de las FCC y plantean que la etapa anterior al quiste presenta una capa fibrilar. De acuerdo a nuestras observaciones sobre la estructura de las formas vegetativas (FCC, FGra, FAmeb) pareciera ser que los quistes se originan a partir de cualquiera de ellas. No se sabe con certeza cual es el proceso, pero pareciera que B. hominis sintetiza los componentes de la pared y los deposita justo entre la membrana plasmática y la cubierta externa o glicocálix, luego esta última se va separando a medida que el quiste madura y finalmente puede ser eliminada para dejar el quiste desnudo. Por esta razón pueden ser observados quistes con o sin cubierta externa. Las características tanto de la pared como de la cubierta externa coinciden con los reportes de Stenzel y Boreham (1991), Boreham y Stenzel (1993), Zaman et al. (1995). Llama la atención en nuestro estudio la variación en el grosor de la capa fibrilar de las diferentes variantes morfológicas y la separación que se produce después de la formación del quiste, demostrada en la secuencia de imágenes presentada. Esto nos sugiere que esta cubierta la produce B. hominis como un material de excreción por lo cual es necesario realizar estudios con métodos de marcaje citoquímico y así dilucidar su origen y composición. La presencia de la cubierta externa ayuda en la identificación de los quistes de B. hominis en el caso de que solo existan estas formas en las heces del portadory contrario a lo que afirma Zaman et al. (1997), su ausencia sí lo dificulta. Sería necesario investigar cual es el proceso mediante el cual los quistes al ser ingeridos siguen su desarrollo en el intestino, bien sea en la superficie de la mucosa o en las profundidades de ella. Suresh et al., (1994), reportan algunos cambios ultraestructurales en el medio de enquistamiento y como aporte para la comprensión del proceso, plantean que la forma ameboide es el estado intermedio entre la FCC y la FQ, destacan la presencia de RER y abundantes ribosomas, lo cual fue observado por nosotros en este estudio y es una evidencia morfológica de la síntesis de proteínas. Según lo descrito en el enquistamiento de Acanthamoeba spp. y de Entamoeba histolytica, se evidencia el empaquetamiento de ribonucleoproteinas que son identificados morfológicamente como cuerpos cromatoides. En nuestros resultados observamos FQ con 5 estructuras internas, que no se corresponden ni con núcleos ni con mitocondrias y se parecen más a células hijas encerradas dentro del quiste (Figura 10). Si nos ubicamos en B. hominis como protozoario, lo mas parecido a estas estructuras, serian células hijas. Suresh et. al, (2002) menciona que en caso de pacientes con infecciones recurrentes por B. hominis, particularmente después del tratamiento, se puede dar un proceso parecido a una esporulación. Por otra parte, si nos planteamos la posibilidad de otra ubicación taxonómica para este organismo, este hallazgo puede dar bases para hacer estudios en esa dirección (Silberman et al, 1996). Según Suresh et. al, (2002), y Zhang et al (2007), B. hominis muestra diversas formas de reproducción dentro del mismo esquema de los protozoarios, división binaria, endodiogenia, plasmotomía, brotamiento, esporulación y otro similar a la esquizogonia dentro de “sacos” en el cuerpo central del parásito. Sin embargo no se han encontrado suficientes evidencias de tales propuestas, lo que hace más interesante nuestro planteamiento sobre la observación de células hijas en el interior de lo que hemos llamado quistes. Las evidencias morfológicas nos permiten sugerir que en el proceso de enquistamiento de B. hominis, las FCC al sufrir transformaciones serían las precursoras de los quistes. Se podría extrapolar, sujeto a comprobación, una posible secuencia en el enquistamiento que se iniciaría con una intensa actividad ribosomal en el citoplasma, paso de material hacia el cuerpo central, condensación de este organelo volviéndose más pequeño con la posterior desaparición de su membrana y finalmente el origen a los quistes. Los componentes de la pared quística serían sintetizados por el organismo y la depositaría por fuera de la membrana citoplasmática, entre ésta y el glicocálix, el cual terminaría separándose de la pared y dando el aspecto de quistes con cubierta, para finalmente ser eliminada y dar origen a los quistes desnudos. En el intestino podría pasar lo mismo originando así la forma de resistencia y estado infectante de B. hominis, capaz de resistir las condiciones adversas del ambiente y permitir la transmisión de un individuo a otro. Es necesario diseñar experimentos para la debida comprobación de este planteamiento. La observación y seguimiento de las diferentes formas de B. hominis en los cultivos nos suministra una información importante que enriquece el conocimiento sobre la biología de este organismo que ha generado tantas controversias. Tiene consecuencias directas en la identificación en las muestras de heces, su correlación con la situación clínica de los hospederos humanos, ciclo evolutivo y estudios de taxonomía. Este trabajo fue financiado por los proyectos de investigación: CDCH 09-12-3366-94, CDCH 09-12-3368-94, ingresos propios del Laboratorio de Amibiasis, Cátedra de Parasitología y recursos de la Escuela de Bioanálisis, Facultad de Medicina, UCV. Referencias
Copyright 2007 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela The following images related to this document are available:Photo images[va07006f9.jpg] [va07006f2b.jpg] [va07006f10.jpg] [va07006f2a.jpg] [va07006f1.jpg] [va07006f5.jpg] [va07006f8b.jpg] [va07006f6.jpg] [va07006f7.jpg] [va07006f4a.jpg] [va07006f3.jpg] [va07006f8a.jpg] [va07006f4b.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}