 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
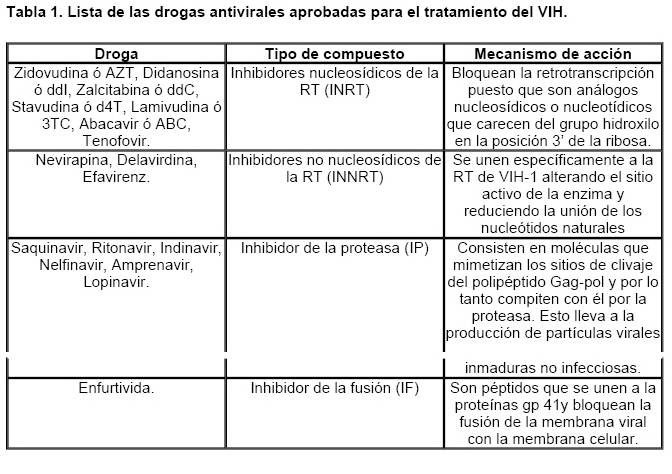
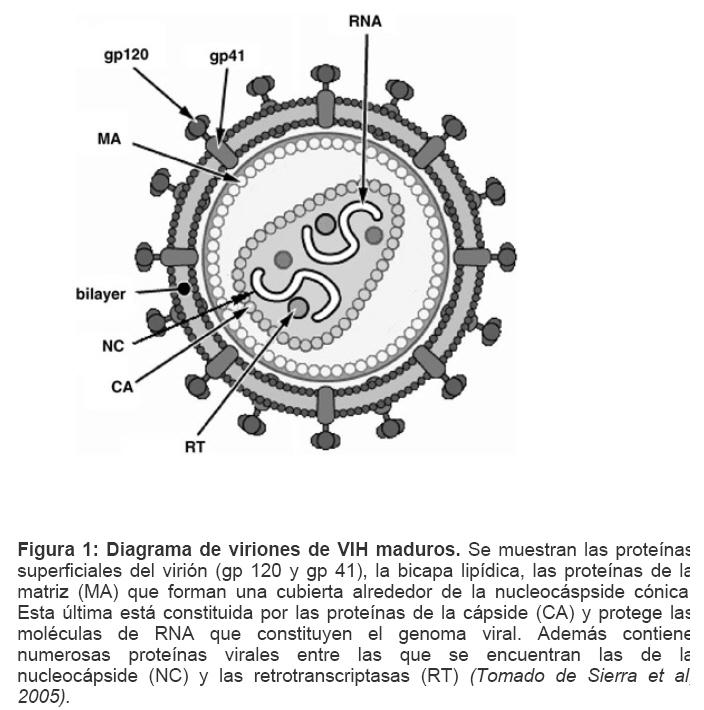
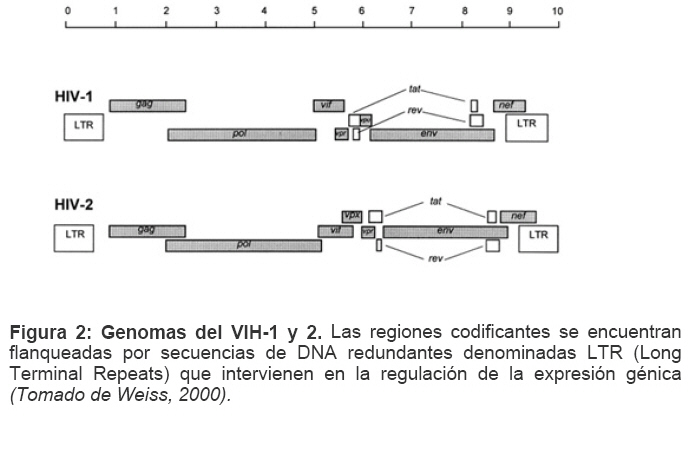
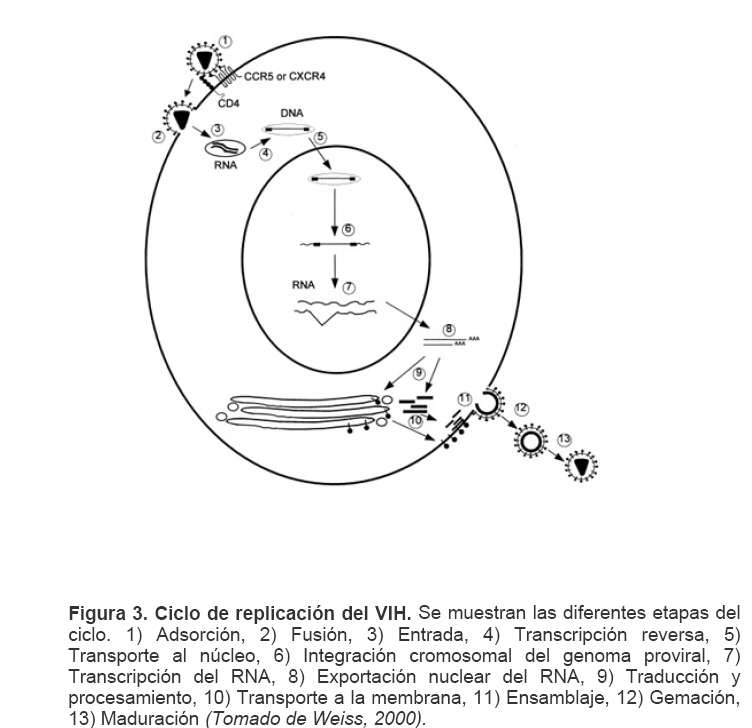
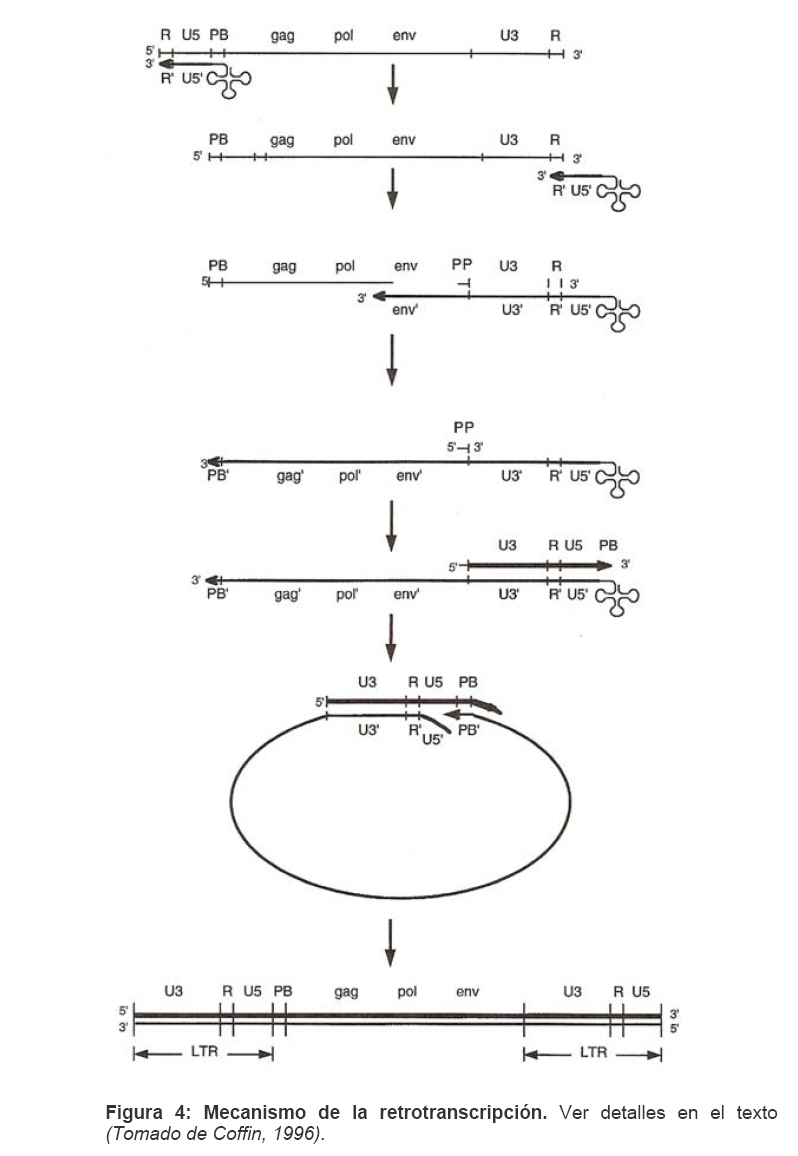
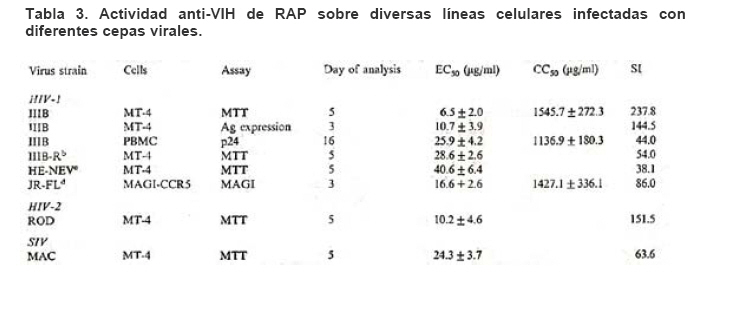
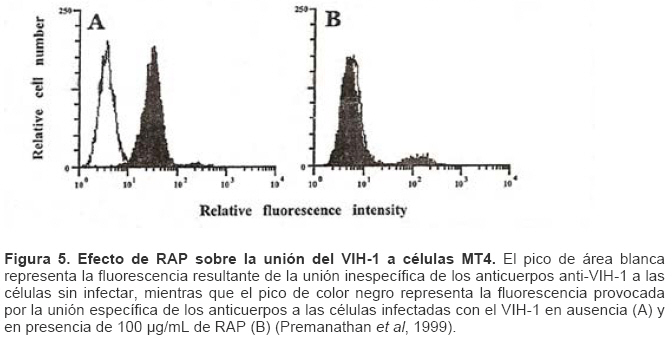
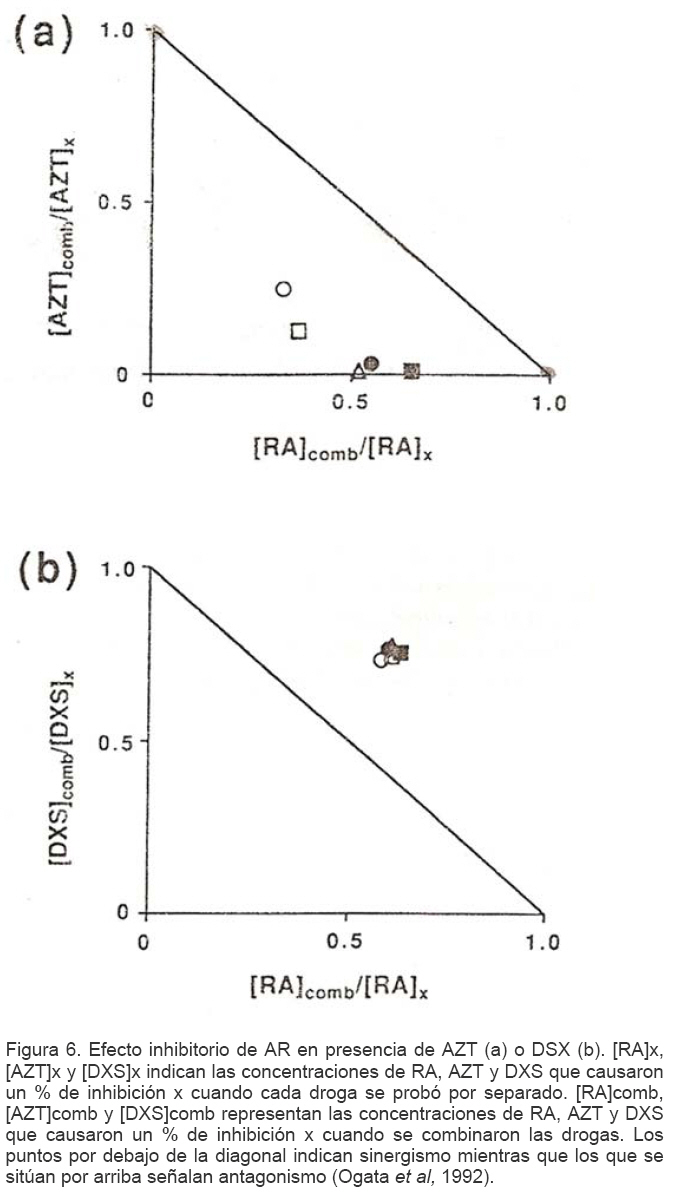
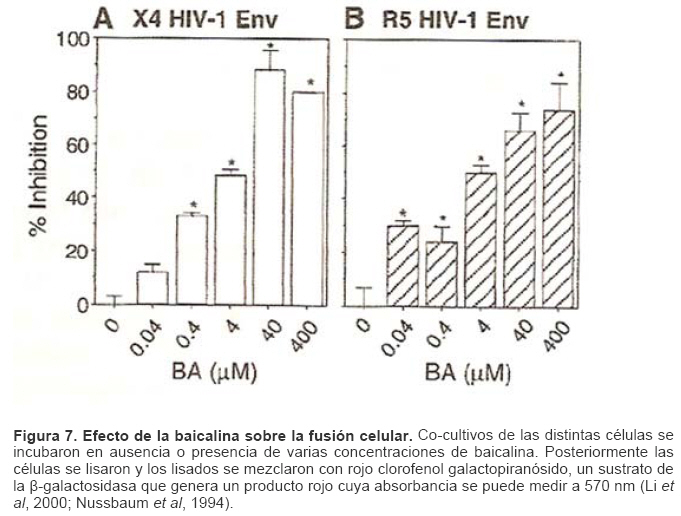
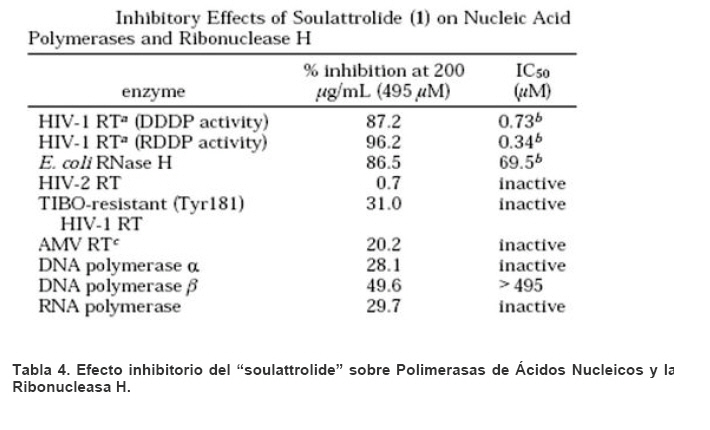
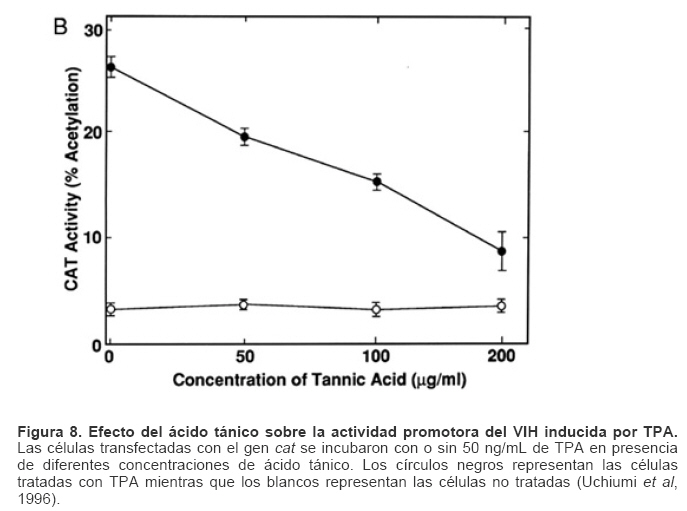
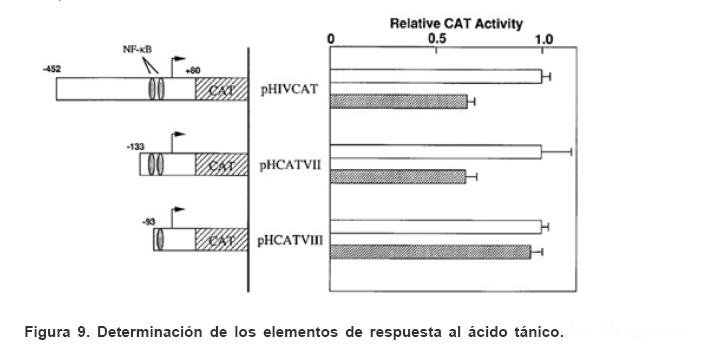
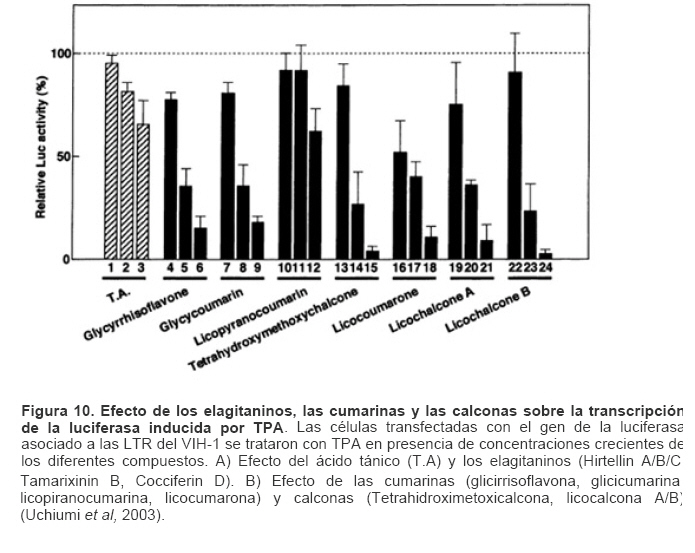
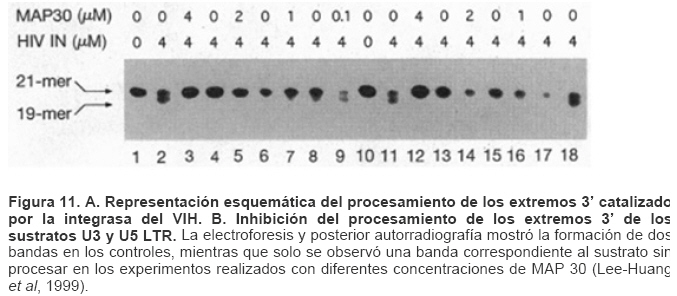
Mecanismos de acción de compuestos antivirales aislados en plantas: el Virus de la Inmunodeficiencia Humana como modelo. Mechanism of action of antiviral compounds isolated from plants: HIV virus as a model. Manuel Okatami Perera Lecoin okacomi@gmail.com Fecha de recepción: 15/07/2007 Code Number: va07012 El SIDA causa más de 3 millones de muertes anualmente a nivel mundial, siendo el VIH el agente etiológico de la enfermedad. Actualmente no existe una vacuna efectiva contra el VIH, por lo que las estrategias adoptadas para enfrentar la epidemia están enfocadas hacia la prevención de nuevas infecciones y el desarrollo de fármacos dirigidos contra el virus. Estas drogas han permitido prolongar la esperanza de vida de los pacientes con SIDA, pero su alto costo, escasa disponibilidad y efectos adversos, han motivado el hallazgo de nuevos compuestos de origen natural con actividad anti-VIH que no resulten nocivos, sean accesibles y de bajo costo. La búsqueda se ha concentrado en plantas debido a su amplio registro etnomedicinal, por lo que la presente revisión bibliográfica pretende describir y analizar los mecanismos de acción de las sustancias anti-VIH obtenidas de plantas. Estos agentes actúan a diferentes niveles de la replicación viral y sobre distintos blancos, permitiendo vislumbrar la posibilidad de generar nuevos tratamientos que permitan combatir eficazmente la enfermedad. Palabras Claves:SIDA; IF; INRT; INNRT. Abstract AIDS is responsible for more than 3 million deaths annually worldwide and the HIV is the etiologic agent of the disease. At the moment, there isn’t an efficient vaccine against HIV, thus the strategies adopted to face the epidemy have been focused on the prevention of new infections and to the development of medicaments against the HIV. These drugs have been able to increase the life expectancy of AIDS patients, but their high cost, scarce availability and adverse effects have led to find new compounds with anti-HIV activity which would be harmful, accessible and not expensive. Research has focused in plants because of their extensive ethnomedicinal reports. Therefore, the present review pretends to analyze and describe the mechanisms of action of anti-HIV substances obtained from plants. These agents have different targets and act at different levels of viral replication, suggesting the possibility of new treatments which could deal efficiently with the disease. Key Word: AIDS; FI; NRTI; NNRTI. Introducción El SIDA (Síndrome de la Inmunodeficiencia Adquirida) es hoy en día una de las enfermedades infecciosas que causa mayor número de muertes anualmente. Para el año 2005, 40.3 millones de personas padecían de SIDA y 3.1 millones de individuos fallecieron por enfermedades relacionadas con este síndrome, de los cuales 2.400.000 eran originarios del áfrica subsahariana. Esta es la región más afectada, con más de 25 millones de infectados y el mayor número de nuevos casos (3.2 millones en 2005). (Zarocostas, 2005). El agente causal del SIDA es el virus de la inmunodeficiencia humana (VIH), el cual fue descubierto en 1983 por un equipo de investigadores del Instituto Pasteur en París, aunque no fue sino hasta 1984 que el hallazgo tomó relevancia tras la confirmación por parte de investigadores estadounidenses de que un retrovirus era el responsable de la enfermedad (Hirsch y Curran, 1996). Éste virus causa una disminución progresiva de los linfocitos T CD4 +, ya que estos constituyen el principal blanco del virus. Debido a que estas células forman parte importante del mecanismo de defensa del organismo, ello favorece la aparición de infecciones oportunistas que resultan letales (Sierra et al, 2005). El VIH puede ser dividido en dos tipos, VIH-1 y VIH-2, en base a las propiedades serológicas y al análisis de las secuencias del genoma viral (Luciw, 1996). El VIH-1 se subdivide en nueve subtipos o clados (A a I) y es el virus que muestra la mayor prevalencia a escala mundial, siendopredominante en Asia, África, Europa y América. El VIH-2 se subdivide en 5 subtipos (A a E), exhibe un periodo de latencia superior al del VIH-1 y es la forma predominante en algunos países de África occidental, aunque se ha expandido a Europa (particularmente a Portugal) y a la India (Luciw, 1996. Weiss, 2000). El análisis de la secuencia nucleotídica relaciona los genomas del VIH con virus de la inmunodeficiencia aislados en simios (SIV), por lo que se cree que el VIH se introdujo en las poblaciones humanas por zoonosis (Weiss, 2000). Debido a que el SIDA representa un problema de salud a escala mundial, muchas investigaciones se han dirigido al estudio de la biología del VIH con la finalidad de determinar posibles blancos terapéuticos o desarrollar medidas preventivas efectivas contra la enfermedad. La elaboración de una vacuna contra el VIH ha sido infructuosa debido a la alta tasa de variabilidad genética del virus, que provoca la aparición continua de formas virales mutantes que logran evadir eficazmente el sistema inmune del hospedador (Esparza, 2005). Esto ha llevado al diseño de estrategias alternativas para combatir la pandemia. Una de ellas consiste en lanzar campañas de educación destinadas a prevenir nuevas infecciones. Según el Programa de las Naciones Unidas para el SIDA, una campaña efectiva de información podría reducir a menos de 2 millones el número de nuevas infecciones anuales, pero esto requeriría una inversión de 15 a 20 billones de dólares anuales en los países en vías de desarrollo, una suma insostenible dada su situación económica (Esparza, 2005). Por otro lado, se han desarrollado fármacos con el objeto de combatir la infección. En los últimos años han aparecido en el mercado medicamentos (Tabla 1) que si bien no erradican por completo la infección, sí permiten aumentar considerablemente la esperanza de vida de los pacientes que los utilizan. Estos medicamentos se clasifican según su mecanismo de acción en inhibidores de la proteasa, inhibidores de la fusión e inhibidores de la retrotranscriptasa. Éstos últimos se pueden separar a su vez en nucleosídicos y no nucleosídicos (Tabla 1). No obstante, menos del 7% de la población infectada con VIH tiene acceso a estos tratamientos debido a sus altos costos y escasa disponibilidad (Esparza, 2005). Asimismo, el uso de estos medicamentos puede causar efectos adversos como diarreas, dolores de cabeza, anemia, anorexia, dolores abdominales, erupciones, entre otros (Mullin et al, 1999). Debido a que el 80% de las personas que viven en países en vías de desarrollo dependen casi exclusivamente de la medicina tradicional para remediar sus problemas de salud, ha resultado particularmente interesante la búsqueda de terapias anti-VIH basadas en compuestos naturales antivirales presentes en organismos vivos (Cos et al, 2004). Este tipo de investigaciones permitiría elaborar tratamientos accesibles y de bajo costo que combatan eficazmente el SIDA en los países que más lo requieren. Ello ha motivado la búsqueda de dichas sustancias principalmente en plantas, debido a que existe un registro extenso de su uso etnomedicinal (Cos et al, 2004). Asimismo, numerosas investigaciones han demostrado que una amplia gama de agentes virales es susceptible al tratamiento con extractos de plantas. En efecto, ciertos compuestos naturales como los flavonoides, terpenoides, lignanos, alcaloides, y saponinas, entre otros, exhiben una actividad antiviral contra virus como el de la influenza, los de la hepatitis B y C, el del herpes simple, los poliovirus de tipo I y los rotavirus (Jassim y Naji, 2003). Las investigaciones con el VIH han suministrado resultados alentadores con el descubrimiento de ciertos flavonoides, cumarinas, polisacáridos no sulfatados, triterpenos y proteínas que exhiben actividad anti-VIH actuando sobre diferentes etapas del ciclo de replicación. Debido a la importancia que tiene el encontrar tratamientos efectivos contra el SIDA, el objetivo del presente trabajo es realizar una revisión bibliográfica que permita describir y analizar los mecanismos de acción de algunos compuestos naturales aislados en plantas con propiedades anti-VIH, para su potencial uso como agentes antiretrovirales. Mecanismos de replicación del VIH Antes de discutir los trabajos relacionados con las propiedades anti-VIH de algunos compuestos naturales, es importante comprender los pasos del ciclo de multiplicación del virus, ya que ello permitirá entender cómo y donde actúan estas sustancias. El VIH pertenece al género lentivirus de la familia Retroviridae. Los viriones maduros miden entre 100 y 120 nm de diámetro y presentan una membrana lipídica que rodea una nucleocápside de forma cónica en la que se encuentra el genoma viral y ciertas proteínas virales (Figura 1): la retrotranscriptasa, la proteasa, la integrasa y las proteínas Vif, Vpr, Nef, Vpu (en el caso del VIH de tipo 1) y Vpx (en el caso del VIH-2) (Luciw, 1996. Sierra et al, 2005). También se encuentran ciertos factores celulares, en particular a nivel de la membrana del virión que es adquirida cuando la partícula viral gema a través de la membrana plasmática de la célula durante le última fase de la infección (Sierra et al, 2005). El genoma está constituido por dos moléculas idénticas de RNA de simple cadena de 9.2 kb, con polaridad positiva y que sirven de templado para la retrotranscriptasa viral. Esta enzima transforma el RNA viral de simple cadena en DNA de doble cadena que luego se incorpora al genoma de hospedador. La forma viral integrada y latente se denomina provirus. El genoma proviral presenta una región codificante central en la que se encuentran los genes virales, algunos de los cuales se encuentran solapados (Figura 2) (Luciw, 1996). Flanqueando a la región codificante se encuentran secuencias de DNA redundantes (idénticas tanto en el extremo 3’ como en el extremo 5’) denominadas LTRs (del inglés “Long Terminal Repeats”). Las LTRs son regiones reguladoras de la expresión génica y se encuentran divididas en tres dominios con distintas funciones (U3, R y U5) (Coffin, 1996). El dominio U3 contiene elementos promotores de la transcripción y secuencias reconocidas por otros factores celulares que modulan la transcripción al influir en la tasa de iniciación. El dominio R presenta el elemento de respuesta de activación en trans (Tar, del inglés “trans-activation response element”). Tar es una secuencia nucleotídica que se encuentra tanto en el RNA como el DNA viral formando unbucle que es reconocido por la proteína viral Tat. Ésta controla la expresión de los genes virales (Luciw, 1996. Coffin, 1996). La transcripción se inicia en el límite entre la región U3/R y los transcritos que se producen tienen las modificaciones características de los RNAm eucarióticos: un extremo 5’ poliadenilado y una caperuza de 7-metil-guanosina en el extremo 3’ (Cann, 2001). Los genes virales y la función de sus respectivos productos aparecen señalados en la Tabla 2. La infección con el VIH comienza cuando los viriones se unen a las células mediante la interacción entre la glicoproteína gp120, presente en la cubierta viral, y el receptor celular CD4 situado en la membrana plasmática de macrófagos y linfocitos T colaboradores, aunque se han encontrado receptores alternativos como la galactosil ceramida en células CD4- (Luciw, 1996. Sierra et al, 2005). La unión de la gp120 al receptor CD4 causa un cambio conformacional en la glicoproteína exponiendo una región que seune a los co-receptores celulares CCR5 y CXCR4. Éstos determinan el tropismo de las cepas de VIH: las que utilizan el co-receptor CXCR4 infectan los linfocitos T (T trópicas), mientras que las que interaccionan con CCR5 infectan a macrófagos y monocitos (M-trópicas) (Princen y Schols, 2005). Después de la unión a los co-receptores, la membrana viral se fusiona con la membrana celular y la nucleocápside es liberada al citoplasma (Figura 3) (Luciw, 1996). Una vez en el citoplasma, ocurre la decapsidación de la nucleocápside viral mediante la acción de factores celulares y de las proteínas virales MA, Nef y Vif (Sierra et al, 2005). El RNA viral que se libera se retrotranscribe generando DNA de doble cadena. Este proceso requiere de un cebador constituido por una molécula de tRNAlys que se encuentra ubicada cerca del extremo 5’ del RNA viral unida a una secuencia denominada PBS (Primer Binding Site) (Luciw, 1996). La primera cadena de DNA se extiende hacia el extremo 5’ del RNA viral generando un pequeño fragmento que es complementario al dominio R del genoma viral (Cann, 2001). La actividad RNAsa H de la RT degrada el complejo RNA/DNA y libera el DNA recién sintetizado que se une al domino R del extremo 3’ del RNA viral (Figura 4). Esta es la primera transferencia de hebra y viene seguida por la síntesis integral de la primera cadena de DNA. La segunda hebra de DNA requiere de la presencia de dos cebadores situados en la cadena de DNA que sirve de templado: dos motivos polipurínicos (PPT) situados en el límite del dominio U3 y al final del gen pol (cPPT). La síntesis de DNA que se inicia en el límite de U3 genera un fragmento que se une al extremo 3’ del DNA molde por complementariedad y que se sigue extendiendo hacia el extremo 5’ del mismo por la acción de la RT (Figura 4). Simultáneamente, se lleva a cabo la síntesis de DNA desde cPPT hacia el extremo 5’ del DNA molde, de manera tal que se generan dos fragmentos de DNA con una secuencia repetida de alrededor de 100 nucleótidos. Esta secuencia es desplazada, escindida y los dos fragmentos de DNA son ligados por la acción de enzimas celulares, generando el DNA linear de doble cadena (Luciw, 1996. Cann, 2001). La retrotranscripción es el principal proceso responsable de la diversidad antigénica del virus ya que la retrotranscriptasa carece de la actividad exonucleasa 3’-5’, presente en las DNA polimerasas celulares, por lo que la tasa de mutaciones es muy superior a la que se registra en células eucariotas. Se estima que el genoma viral experimenta de una a tres mutaciones por ciclo de replicación (Luciw, 1996). Una vez sintetizado el DNA viral, éste se dirige al núcleo mediante la acción de Vpr, una proteína que ancla el DNA viral a la membrana nuclear permitiendo que penetre al núcleo a través de los poros nucleares (Sierra et al, 2005). El DNA es incorporado al genoma del hospedador mediante la acción de la integrasa viral que realiza cortes escalados en los genomas celular y viral, uniéndolos y rellenando los espacios dejados por las bases no pareadas (Cann, 2001). Las primeras rondas de transcripción llevadas a cabo por la RNA polimerasa II celular sobre el DNA viral integrado, generan varios transcritos que migran al citoplasma mediante una vía independiente de Rev. Durante esta fase de replicación temprana se producen múltiples cortes y empalmes de los RNAm y sólo las proteínas regulatorias Tat, Nef y Rev se expresan. Al alcanzar niveles suficientes, Tat se une a Tar y controla la transcripción de los genes virales. Cuando Rev alcanza niveles adecuados, se une a una estructura de los transcritos denominada elemento de respuesta a Rev (RRE) facilitando su paso a través de la membrana nuclear y protegiéndolos del “splicing”, de manera que se generan RNAm sin cortes o con un único corte que producen las otras proteínas virales y el RNA genómico (Sierra et al, 2005). El gen env, que codifica para las proteínas de unión a CD4 (gp120) y para las proteínas virales transmembranales (gp41), se expresa inicialmente como una poliproteína precursora denominada gp160. Ésta es glicosilada dentro del retículo endoplasmático y el aparato de Golgi, en donde una endoproteasa escinde la proteína para generar gp120 y gp41 (Luciw, 1996). Estas proteínas son transportadas por vesículas celulares hasta la membrana plasmática. Los genes gag-pol se traducen para generar la poliproteínas Gag y Gag-pol que son las precursoras de las 6 proteínas estructurales codificadas por gag y las proteasas, integrasas y retrotranscriptasas codificadas por pol. Estas poliproteínas son procesadas proteolíticamente durante la maduración (Sierra et al, 2005). Después de la traducción, las proteínas Gag y Gag-pol migran hacia la membrana celular y empiezan a ensamblarse dirigidas por Gag. Debido a que las proteínas virales se expresan sobre la superficie celular, se puede producir la fusión entre células infectadas y células sanas que expresen el receptor CD4 generando células gigantes multinucleadas (sincitios). La formación de sincitios constituye uno de los mecanismos de propagación del virus y es parcialmente responsable de la disminución progresiva del número de linfocitos T característico del SIDA (Geleziunas y Greene, 2000). Durante el ensamblaje de las partículas virales se asocian las enzimas virales, el genoma viral, el cebador celular RNAtlys y otros compuestos celulares. Los viriones inmaduros geman a través de la membrana plasmática, lo que activa la proteasa viral que escinde las poliproteínas Gag y Gag-pol, liberando las proteínas estructurales y las enzimas (Sierra et al, 2005). Todas ellas interactúan con las proteínas de la cápside (CA) y de la nucleocápside (NC) para generar los viriones maduros. El proceso de ensamblaje y gemación requiere de la disminución del número de moléculas CD4 celulares para evitar su interacción con las proteínas gp120 recién sintetizadas. En este proceso intervienen las proteínas Env, Nef y Vpu. La primera se une a las moléculas de CD4 que pasan por el retículo endoplasmático durante su maduración, mientras que la segunda acelera la endocitosis y la degradación subsecuente de CD4. La proteína Vpu produce canales iónicos en la membrana plasmática que incrementan la tasa de liberación de virus (Luciw, 1996. Sierra et al, 2005).Una vez conocido en detalle el mecanismo molecular mediante el cual el virus lleva a cabo su multiplicación, muchos investigadores se han enfocado en desarrollar terapias que bloqueen algunos de los pasos antes descritos. En particular, ciertos grupos de investigación han centralizado su interés en estudiar como actúan los diferentes compuestos naturales con propiedades anti-VIH aislados de plantas. Compuestos naturales de plantas con actividad anti-VIH En los últimos años se han descubierto en las plantas una amplia gama de moléculas que inhiben la infección por VIH, al actuar sobre diferentes etapas del ciclo de replicación viral, desde fases tempranas hasta fases más tardías. Sustancias que actúan en etapas tempranas del ciclo de replicación viral. Se considera que la adsorción de la partícula viral y la fusión a la célula diana constituyen las etapas tempranas del ciclo de replicación viral. Se han identificado compuestos que son capaces de actuar solamente sobre dichas fases, inhibiendo así la entrada del virus a la célula hospedadora. Los inhibidores de la adsorción: la fracción polisacárida de Rhizophora apiculata. Los manglares del género Rhizophora se encuentran distribuidos a lo largo de las costas orientales de África, así como en el sureste de Asia y en Oceanía. Las hojas y la corteza de estas plantas han sido tradicionalmente utilizadas en esas regiones como antisépticos y para tratar diarreas, fiebres, disentería, lepra y malaria (Duke, 2005). En tal sentido, un grupo de investigadores tomó la iniciativa de estudiar el efecto antiviral de esta planta sobre la infección por VIH. Estudios previos habían demostrado que los polisacáridos sulfatados son inhibidores de la multiplicación in vitro del VIH 1 y 2, siendo la heparina, el pentosan polisulfato y los sulfatos de dextrina y dextrano los de mayor efecto. Estos compuestos inhiben la adsorción viral al interferir en la unión de la proteína gp120 al receptor CD4 (Cos et al, 2004). Así, Premanathan et al. (1999) decidieron investigar si se obtenía el mismo efecto sobre el VIH con las fracciones polisacáridas obtenidas de las hojas de Rhizophora apiculata (RAP). Para ello, los autores realizaron la purificación previa de los polisacáridos de la hoja de la planta y luego elaboraron ensayos de inhibición de la citopatogenicidad y replicación del virus. Para estimar la inhibición de la citopatogenicidad, Premanathan et al. (1999) investigaron el efecto de RAP sobre la viabilidad celular y la formación de sincitios en células infectadas con VIH. La disminución de la viabilidad celular fue determinada a través de la disminución de la actividad de una deshidrogenasa mitocondrial, medida por la acción de esta enzima sobre el sustrato MTT (bromuro de 3-(4,5-dimetiltiazol-2 il)-2,5- difenil-tetrazolio), el cual es oxidado originando un producto cristalino azul que puede solubilizarse en medio ácido y cuya absorbancia se mide a 540 nm (Pauwels et al, 1988). La inhibición de la formación de sincitios fue determinada mediante el método de exclusión de azul de tripano. Este último es un colorante que sólo penetra en las células que han perdido la integridad de su membrana plasmática (Pauwels et al, 1988), así que aquellas que formen sincitios incorporarán el compuesto. Por otro lado, Premanathan et al. (1999) estimaron la inhibición de la replicación mediante la producción de antígenos virales y la Activación Multinuclear del Indicador de la Galactosidasa (en inglés, MAGI). La medición de antígenos virales se realizó mediante el método de inmunofluorescencia indirecta, mientras que la detección específica de la proteína p24 se hizo por el método de ELISA. El ensayo MAGI permite determinar el número de partículas infecciosas de VIH en una muestra particular y se basa en la expresión de la enzima β-galactosidasa en células transfectadas con el gen lacZ (células MAGI). Este se encuentra asociado al promotor viral Tar, por lo que su transcripción sólo se inicia cuando el virus penetra a las células e inicia su replicación, produciendo la proteína Tat que controla la transcripción genética al unirse a Tar. El número de células infectadas se puede calcular contando todas aquellas que se tornan azules en presencia de un sustrato adecuado de la β-galactosidasa (Kimpton y Emerman, 1992). En todos los ensayos anteriores, los autores demostraron que las fracciones RAP inhibían la replicación viral y la citopatogenicidad causada por cepas de VIH-1 y 2 en diversas células de cultivo (Premanathan et al, 1999). En efecto, a concentraciones relativamente bajas, los compuestos ensayados disminuyeron a la mitad el número de células azules en ensayos MAGI (ver en la Tabla 3 la EC50, definida como la concentración efectiva de polisacáridos necesaria para reducir en un 50% la citopatogenicidad inducida por el virus o la replicación viral). Al evaluar la citotoxicidad de los compuestos sobre células no infectadas, se registró un daño celular considerable a concentraciones altas de los polisacáridos (ver la CC50 en la Tabla 3, que representa la concentración de RAP necesaria para disminuir en un 50% la viabilidad de células sanas), generando un índice de selectividad alto (SI = CC50/EC50) que reflejó cuan aptos eran los compuestos para disminuir la replicación viral sin causar daño a las células. En células T MT4 infectadas con diferentes cepas de VIH 1 y 2, los polisacáridos inhibieron efectivamente (índice de selectividad > 100) la citopatogenicidad y la producción de antígenos virales asociadas a las infecciones virales (ver Tabla 3). No obstante, tuvieron un menor efecto inhibitorio sobre cepas resistentes a dos INNRT como la nevirapina (HE-NEV) y la emivirina (IIB-R), sugiriendo la posibilidad de que RAP actúe sobre la RT viral de forma similar a estos inhibidores. La prevención de la formación de sincitios entre células T infectadas con VIH-1 y células T sanas, fue directamente proporcional a la cantidad de RAP, pero la concentración necesaria para obtener una inhibición del 50% fue 4 veces mayor a la de sulfato de dextrano. La EC50 representó la concentración de compuesto necesaria para disminuir en un 50% el número de células MAGI azules y la cantidad de antígenos virales con relación a los controles no tratados con el extracto. En el caso de los ensayos colorimétricos con MTT, la EC50 se definió como la cantidad de RAP requerida para aumentar en un 50% la absorbancia de las células con relación a células no expuestas a RAP. La CC50 fue la concentración de extracto que disminuyó en un 50% la viabilidad celular con respecto a células sanas no expuestas a los compuestos. El índice de selectividad (SI) representa cociente CC50/EC50 (Premanathan et al, 1999). Los ensayos de detección de p24 realizados en células periféricas mononucleares de la sangre (PBMC) mostraron un EC50 relativamente alto en relación con el CC50, obteniendo un SI bastante bajo (44), lo que reflejó una menor eficiencia de RAP a la hora de inhibir la infección en células mononucleares aisladas de pacientes sanos (Premanathan et al, 1999). La EC50 representó la concentración de compuesto necesaria para disminuir en un 50% el número de células MAGI azules y la cantidad de antígenos virales con relación a los controles no tratados con el extracto. En el caso de los ensayos colorimétricos con MTT, la EC50 se definió como la cantidad de RAP requerida para aumentar en un 50% la absorbancia de las células con relación a células no expuestas a RAP. La CC50 fue la concentración de extracto que disminuyó en un 50% la viabilidad celular con respecto a células sanas no expuestas a los compuestos. El índice de selectividad (SI) representa cociente CC50/EC50 (Premanathan et al, 1999) Debido a que los polisacáridos sulfatados con actividad antiviral actúan al nivel de la adsorción del virus, los mismos autores decidieron estudiar si el efecto antiviral de RAP se debía a su interacción sobre la unión de los viriones a células T. Para tal fin, las células MT-4 se infectaron con una cepa de VIH-1 en presencia o ausencia de RAP y se incubaron con un anticuerpo policlonal aislado de un paciente infectado con VIH. Posteriormente se agregó un anticuerpo de conejo dirigido contra las inmunoglobulinas humanas y conjugado a fluoresceína, una molécula que fluorece a determinada longitud de onda. Las células marcadas de este modo se analizaron en un citómetro de flujo (Schols et al, 1989). En las células infectadas tratadas con el extracto, los dos picos de fluorescencia se solaparon (Figura 5), indicando un bloqueo en la unión de los anticuerpos primarios anti-VIH a los viriones. Por lo tanto concluyeron que RAP actuaba inhibiendo la adsorción viral, lo que se vio confirmado al medir la expresión del RNAm viral mediante ensayos cuantitativos de RT-PCR. En este último ensayo, al tratar las células con los polisacáridos durante la adsorción viral se redujo significativamente la cantidad de RNAm viral, mientras que al tratar las células después de la fase de adsorción no se observó efecto. Por otro lado, Premanathan et al. (1999) observaron que RAP previno la formación de sincitios sin afectar la unión de anticuerpos dirigidos contra CD4, CXCR4 y CCR5, indicando que la inhibición de la adsorción podría deberse a la interacción de RAP con la proteína gp120. Los grupos sulfato de muchos polisacáridos son los responsables de este efecto. Sin embargo, no se detectaron grupos sulfatados en las fracciones RAP, representando una categoría novedosa de polisacáridos con propiedades anti-VIH (Premanathan et al, 1999). En la prevención de la adsorción podrían estar interviniendo los grupos carboxilados de los ácidos urónicos detectados en las fracciones RAP. Estos, al igual que los grupos sulfatados, tienen un carácter aniónico y podrían interactuar con los residuos básicos de gp120. Otros inhibidores de la adsorción: el ácido repandusínico de Phyllanthus niruri. Existen otros compuestos que actúan al nivel de la adsorción. Tal es el caso del ácido repandusínico (AR) aislado de Phyllantus niruri. Ésta es una hierba medicinal que se ha empleado como diurético y para el tratamiento de la ictericia. En experimentos previos se reportó que el extracto acuoso de la planta presentaba un efecto antiviral al actuar sobre la DNA polimerasa del virus de la hepatitis B y sobre la RNA polimerasa del virus de la mieloblastosis aviar (Ogata et al, 1992). Aunque se ha encontrado que el AR es un inhibidor de la actividad de la RT del VIH-1, como se describirá más adelante, Ogata et al. (1992) observaron que el compuesto también podía operar sinergísticamente con el AZT (un INRT de actividad anti-VIH conocida), probablemente debido a su capacidad para inhibir la adsorción viral. En efecto, al realizar ensayos de viabilidad celular sobre células T infectadas en presencia de AR y sulfato de dextrano (DXS), se observó un antagonismo entre los dos compuestos, indicando que actuaban del mismo modo (Figura 6). Los ensayos de viabilidad celular se basaron en el mismo principio descrito anteriormente para el MTT, pero el sustrato utilizado fue el 2,3-bis(2-metoxi-4-nitro-5sulfofenil)-2H-tetrazolium-5-carboxanilido (XTT), un compuesto que permite generar un producto cromogénico soluble en presencia de un transferidor de electrones adecuado (Riss y Moravec, 1996). Los inhibidores de la fusión: la baicalina de Scutellaria baicalensis. La baicalina es un flavonoide aislado de las raíces de Scutellaria baicalensis, una planta empleada en la medicina tradicional China para tratar bronquitis, nefritis, hepatitis, asma y dermatitis (Li, et al, 2000). Los flavonoides son compuestos comunes en plantas y presentan actividades biológicas interesantes como vasodilatadores, anti-inflamatorios, anti-alérgicos y antivirales entre otros (Lin et al, 1999). Son compuestos polifenólicos de 15 carbonos con estructuras sumamente diversas que se organizan alrededor de un núcleo común, constituido por 2 anillos de benceno unidos por una cadena de tres carbonos. Se ha determinado que las propiedades anti-inflamatorias de la baicalina parecen deberse a su capacidad para bloquear la interacción de ciertas quimiocinas con los receptores CCR5 y CXCR4, que también juegan un papel clave en la infección por VIH (Li, et al, 2000). Basado en lo anterior, Li y colaboradores estudiaron el efecto de este compuesto sobre la fusión del VIH con células diana. Para tal fin, emplearon dos líneas celulares: una co-expresaba el receptor CD4 y el co-receptor CCR5 (R5) y la otra el receptor CD4 y el co-receptor CXCR4 (X4). Ambas fueron además infectadas con un vaccinia virus recombinante que llevaba el gen que codifica para la RNA polimerasa del bacteriófago T7, el cual se encontraba bajo el control de un promotor del vaccinia virus que se expresa constitutivamente (Hruby, 1990). Paralelamente, se coinfectaron células Hela con dos vaccinia virus recombinantes: uno de ellos contenía el gen env de cepas M-trópicas y el otro contenía el gen reportero lacZ unido al promotor de la RNA polimerasa T7. Por otro lado, introdujeron un vaccinia virus con el gen reportero antes mencionado a una línea celular que ya poseía el gen env con tropismo hacia células T. De esta manera se logró generar un sistema de células capaces de fusionarse entre sí y de recrear el proceso de entrada del VIH-1, pudiendo monitorear y cuantificar la fusión celular gracias a la actividad de la enzima β-galactosidasa, que sólo podía expresarse a partir del promotor de la RNA polimerasa de T7 tras ocurrir la fusión. Midiendo los niveles de b-galactosidasa expresada como consecuencia de la fusión (Figura 7), Li y colaborados (2000) obtuvieron que la baicalina inhibía la fusión mediada tanto por CXCR4 como por CCR5. El grado de inhibición fue proporcional a la concentración de baicalina y la concentración de compuesto necesaria para inhibir en un 50 % la fusión fue similar para ambos tipos de co-receptores (4 µM), por lo que el efecto de la sustancia no dependió del tropismo del VIH-1. Estos resultados se vieron confirmados por la ausencia de células multinucleadas gigantes (sincitios) en los co-cultivos celulares realizados con baicalina. En los sistemas no tratados con el compuesto sí se observó este fenómeno causado por la fusión celular. Los investigadores sugirieron que ciertamente la baicalina inhibía eficientemente la fusión de los viriones de VIH-1 a las células, posiblemente al actuar en etapas tempranas del ciclo de replicación viral. No obstante, para descartar la posibilidad de que el extracto actuara en etapas tardías del ciclo, es decir, a nivel de la formación de sincitios que se producen una vez que las proteínas virales se expresan en la membrana de las células infectadas, los mismos autores realizaron un ensayo de detección del DNA viral durante la fase temprana de la infección viral. Para ello infectaron células con cepas de VIH-1 M y T-trópicas en presencia o ausencia de baicalina. Dejaron transcurrir el tiempo suficiente para que ocurriera la replicación e integración viral, aislaron el DNA celular total y realizaron un PCR anidado en presencia de cebadores específicos para la región R/U5 del LTR del VIH. No se detectaron fragmentos del DNA amplificado en las células tratadas con el compuesto, mientras que en las células no tratadas sí. Esto confirmó la hipótesis de una actuación temprana del compuesto sobre el ciclo viral. Hay que resaltar que, si bien los resultados obtenidos en esta investigación con la baicalina son alentadores, no se realizaron en ningún momento ensayos control para evaluar la citotoxicidad del compuesto, por lo que no se puede excluir la posibilidad de que la baicalina tenga efectos nocivos sobre células sanas. Compuestos que actúan a nivel de la replicación intracelular Se han aislado compuestos de plantas que actúan sobre fases de la infección viral más tardías. Estas sustancias pueden inhibir etapas de la replicación posteriores a la entrada del virus en la célula hospedadora. Entre los blancos de tales compuestos se tienen las enzimas virales tales como las retrotranscriptasas y las proteasas, así como factores que intervienen en el proceso de translocación de los transcritos de RNA al citoplasma. También existen algunos de estos compuestos que pueden interferir en el proceso de integración del genoma viral. Los inhibidores de la retrotranscriptasa viral: las cumarinas aisladas en las plantas del género Calophyllum. Los compuestos naturales de plantas que actúan a nivel de le replicación intracelular del virus son diversos y sus blancos variados. Las cumarinas, por ejemplo, han demostrado tener una actividad anti-VIH muy específica inhibiendo efectivamente la acción de la retrotranscriptasa del VIH-1 (Pengsuparp et al, 1996; Buckheit et al, 1999). Estas moléculas no son análogos nucleosídicos, por lo que entran en la categoría de inhibidores de la retrotranscriptasa no nucleosídicos. Todas ellas comparten una estructura común de benzo-2 pirona. Existen diversas cumarinas que exhiben propiedades anti-RT del VIH-1, las cuales han sido aisladas de árboles de selvas tropicales pertenecientes al género Calophyllum. Uno de estos compuestos es el “soulattrolide”, que representa el 29% de los extractos del látex de Calophyllum teysmanii (Pengsuparp et al, 1996). Estudios llevados a cabo por Pengsuparp y colaboradores (1996), mostraron una inhibición selectiva del compuesto sobre la actividad dependiente de DNA y RNA de la RT del VIH-1, mediante ensayos con la enzima incubada en presencia de timidina trifosfato marcada y de templados adecuados. Independientemente del sustrato (secuencias de poli -rA o DNA de timo de becerro), en presencia de 200 µg/mL (o 495 µM) de “soulattrolide” la actividad de la RT se inhibió en más de un 80% con relación a los controles no expuestos al compuesto (Tabla 4). Las concentraciones necesarias para inhibir en un 50% la actividad retrotranscriptasa (IC50) dependiente de DNA y RNA (Tabla 4), fueron superiores a las obtenidas con AZT (IC50= 0,051 µM), un análogo nucleosídico inhibidor de la RT, pero similares a los de otras cumarinas aisladas en Calophyllum tales como la calanolida A (IC50=0,32 µM) (Pengsuparp et al, 1996). Asimismo, Pengsuparp y colaboradores (1996) también encontraron que el compuesto era capaz de inhibir en más de 80% la actividad RNAsa de E. coli, una enzima que comparte una estructura muy similar a la RNAsa viral (Luciw, 1996). DDDP: actividad DNA Polimerasa Dependiente de DNA; RDDP: actividad RNA Polimerasa dependiente de RNA. El compuesto se consideró inactivo cuando su porcentaje de inhibición fue cercano o inferior a 30% (Pengsuparp et al, 1996). La inhibición de la actividad de la RNAsa fue medida como la cantidad de polímeros sintéticos de RNA que no eran degradados. Para ello, se empleó una mezcla de reacción suplementada con un sustrato sintético de [3H]poli (rA).poli(dT) y ácido tricloroacético (ATC), una solución que permite precipitar los polímeros de ácidos nucleicos superiores o iguales a 20 nucleótidos (Ambion, http://www.ambion.com/techlib/misc/tcapptn.html) De tal manera, la cantidad de productos marcados precipitados obtenida en las mezclas de reacción era indicativa de la inhibición por parte del compuesto. Estos autores encontraron que la cantidad de producto marcado obtenida en mezclas expuestas a una concentración de 495 µM de “soulattrolide”, fue casi 100 veces menor a la registrada en los controles sin el compuesto sugiriendo la posibilidad de que dicha sustancia actuara sobre la RNAsa viral. El “soulattrolide” no inhibió de manera significativa la actividad dependiente de RNA de otras retrotranscriptasas virales tales como la del VIH-2, la del Virus de la Mieloblastosis Aviar y la RT mutante del VIH-1 resistente a la tiobencimidazolona (TIBO), un INNRT (Luciw, 1996). Esto reveló una acción selectiva del compuesto y sugirió la posibilidad de un solapamiento entre los sitios de unión de esta cumarina y del TIBO. El problema de muchos compuestos y medicamentos anti-VIH es que pueden llegar a ser tóxicos para los pacientes que los utilizan regularmente ya que interfieren eventualmente en procesos esenciales para el desarrollo celular. Este es el caso de la AZT, que muestra una IC50 relativamente baja para la DNA polimerasa celular β (11µ M). Para evaluar la toxicidad del “soulattrolide” se realizaron los mismos experimentos que para la RT pero con enzimas celulares tales como las DNA polimerasas α y β eucarióticas y la RNA polimerasa de E coli. Los valores de inhibición obtenidos para la DNA polimerasa α y para la RNA polimerasa fueron inferiores al 30% en presencia de 495 µM del compuesto. Sin embargo la DNA polimerasa β resultó ser más sensible al compuesto, presentando una IC50 cercana a 495 µM. Esto podría resultar problemático si se toma en cuenta que esta enzima parece intervenir en el proceso de reparación de errores durante la replicación del DNA eucariótico (Klug y Cummings, 1999). No obstante los valores de IC50 fueron altos al compararse con los obtenidos con el AZT y con otras cumarinas como la calanolida A (205 µM). La calanolida A, así como su forma isomérica calanolida B (o costatolida) y la dihidrocalanolida A (o dihidrocostatolida), aisladas de Calophyllum lanigerum, exhibieron al igual que el “soulattrolide”, la misma acción selectiva contra la RT de VIH-1 (Buckheit et al, 1999). No obstante, la propiedad más relevante de estas cumarinas fue su capacidad para inhibir sinergísticamente la citopatogenicidad del VIH-1 con una amplia gama de drogas anti-VIH, tal y como lo reveló el estudio realizado por Buckheit y colaboradores (1999) Estos investigadores midieron la viabilidad en células T infectadas con el VIH-1 en presencia de las tres cumarinas citadas anteriormente y una combinación de drogas a diferentes concentraciones. De esta forma observaron que todas las cumarinas inhibían la citopatogenicidad del virus de manera sinergística con los inhibidores nucleosídicos empleados en el ensayo, con e inhibidor no nucleosídico UC781, así como con los inhibidores de la proteasa Ritonavir y Saquinavir. El costatolide, en combinación con INRTs, mostró los mayores niveles de sinergismo entre las tres cumarinas, por lo que podría constituir la base de una terapia que permita incrementar el efecto de los INRTs. La ventaja de esta combinación es que los compuestos no compiten entre sí puesto que se unen a regiones diferentes de la retrotranscriptasa (Buckheit e al, 1999). Otra característica interesante descubierta por Buckheit y colaboradores (1999), fue que las cumarinas analizadas actuaban efectivamente sobre los virus con una mutación puntua en el residuo 181 de la retrotranscriptasa. Esta mutación consiste en el cambio de un residuo de tirosina por uno de cisteína, haciendo que el virus pierda sensibilidad hacia una serie de INNRTs. El residuo de tirosina de la posición 181 se encuentra en un bolsillo situado en la subunidad p66 de la RT, cerca del sitio activo de la polimerasa. A este bolsillo se unen la mayoría de los INNRTs por lo que las mutaciones que ocurren en esta región afectan la unión de estos compuestos y bloquean su efecto (Buckheit et al, 1995). Esto indica que las calanolidas A y B y la dihidrocostatolida actuaron en un sitio de unión distinto al de la mayoría de los INNRTs. Por otro lado, Buckheit y colaboradores (1999) observaron que al hacer sub-cultivos de células T infectadas con el VIH-1 en presencia de concentraciones crecientes de las tres cumarinas, se lograron aislar cepas virales resistentes a estos compuestos ya que no vieron reducida su citopatogenicidad en ensayos con XTT. Sin embargo, los virus resistentes a la calanolida A mantuvieron su sensibilidad hacia la mayoría de los INNRTs probados, así como hacia los INRTs como el AZT y el 3TC. En vista de estos resultados, hay que considerar las cumarinas como un posible complemento en los tratamientos anti-VIH, ya que actúan sinergísticamente con las drogas actualmente en uso y seleccionan virus susceptibles a otros compuestos. Otros inhibidores de la transcriptasa reversa: el ácido repandusínico de Phyllanthus niruri. El ácido AR, tal y como se mencionó anteriormente, es aislado de la planta Phyllantus niruri, la cual ha tenido un extenso uso medicinal. Ello motivó a un equipo de investigadores a estudiar e efecto del extracto acuoso de la planta mencionada sobre la infección por VIH-1 (Ogata et al 1992). Para ello, Ogata y colaboradores (1992) extrajeron, en primer lugar, los componentes de la planta en agua. Después de evaporar el solvente, resuspendieron el residuo sólido en metano generando una fracción soluble y una fracción insoluble que utilizaron para hacer ensayos de inhibición de la RT. Estos se realizaron con una RT recombinante del VIH-1 expresada en E.coli Los investigadores incubaron la RT en un medio de reacción con timidina trifosfato marcada y secuencias de poli (rA) y oligo (dT) como templados y cebadores de la enzima, respectivamente El DNA radioactivo sintetizado se fijó en filtros de intercambio iónico y se estimó su cantidad en un contador de centelleo. El residuo insoluble en metanol presentó la mayor actividad inhibitoria sobre la RT, por lo que Ogata y colaboradores (1992) lo sometieron a fraccionamientos sucesivos y evaluaron la actividad anti-RT de cada fracción. De esta manera se logró aislar e identificar e AR, que bloqueó de manera efectiva la actividad RT del VIH-1 mostrando valores de EC 50 inferiores a los del Foscarnet, un inhibidor específico de la RT del VIH-1. Sin embargo su actividad no fue tan selectiva como la de este último compuesto ya que también inhibió considerablemente la DNA-polimerasa eucariótica α, lo que podría explicar su efecto citotóxico a bajas concentraciones. La actividad DNA polimerasa se midió de la misma manera que para la RT, pero en el medio de reacción se agregó DNA de timo de becerro como sustrato de la enzima (Ogata et al, 1992). Los inhibidores de la interacción REV/RRE: el Niruriside de Phyllanthus niruri. Otro compuesto totalmente distinto a las cumarinas, es el denominado niruriside, el cual ha sido aislado de fracciones metanólicas de la misma planta de donde proviene el ácido repandusínico Phyllanthus niruri. El niruriside actúa de modo diferente sobre el VIH-1, ya que en un estudio se observó que interfiere con la unión de Rev a RRE (Qian-Cutrone et al, 1996). Tal y como se señaló en la primera parte del trabajo, Rev es una proteína se une a los transcritos del genoma viral que contienen el elemento de respuesta a Rev (RRE). Esta interacción protege los RNAm del corte y empalme y facilita su transporte hacia el citoplasma celular. Así, para medir el efecto del niruriside sobre la formación de los complejos Rev/RRE, Qian-Cutrone et al. (1996) utilizaron una proteína Rev expresada en E.coli y un transcrito de RNA marcado con 33P que presentaba e elemento RRE. La formación de complejos entre la proteína y el RNA, se pudo monitorear y cuantificar mediante la transferencia de la proteína Rev a filtros de nitrocelulosa y la medición de la radioactividad en un medidor de centelleo: las proteínas unidas al RNA marcado mostraron actividad radioactiva, mientras que las proteínas libres no. Así, los autores observaron que, s bien el IC 50 obtenido en ensayos de XTT con células T infectadas con VIH-1 fue significativamente alto (260 µM), la cantidad de compuesto necesaria para disminuir en un 50 % la interacción de la proteína Rev con su respectivo sitio de unión en el RNA viral fue de 3,3 µM (Qian-Cutrone et al, 1996). La diferencia entre estos resultados sugiere que la acción de niruriside se ve limitada por su incapacidad para penetrar en las células. Los inhibidores de la proteasa: los triterpenos de Geum japonicum. La planta Geum japonicum se usa en la medicina tradicional China como un antidiurético y se sabe que contiene compuestos con actividad antiviral (Xu et al, 1996). Por tal motivo, el grupo de investigación liderado por Xu (1996), decidió separar los componentes activos de Geum japonicum partiendo de un extracto metanólico de la planta. Guiados por bioensayos, lograron determinar que la actividad antiviral del extracto venía dada por cinco triterpenos. Éstos son lípidos emparentados con los esteroides que derivan de unidades de isopreno. Xu et al. (1996) decidieron estudiar el efecto de estos compuestos sobre la proteasa de VIH-1. Para ello incubaron la enzima en presencia de cada uno de los triterpenos en un medio con un heptapéptido sintético de secuencia aminoacídica idéntica a la del sitio de escisión de las proteínas p24-p17 del precursor Gag. La presencia de los cinco triterpenos durante la reacción inhibió la cantidad de péptido que se obtuvo en ausencia de los compuestos. Los porcentajes de inhibición variaron entre un 42% para el compuesto con menor actividad, hasta un 100 % para e triterpeno más activo (Xu et al, 1996). Los inhibidores de la transcripción: los taninos, las calconas y las cumarinas. La transcripción del genoma viral una vez que se ha integrado al DNA de la célula hospedadora también constituye el blanco de ciertos compuestos. Algunos actúan a nivel de proteínas que participan en la regulación de la transcripción. En efecto, ciertas proteínas de células eucariotas se encuentran unidas covalentemente a homopolímeros de poli(ADP-ribosa) que juegan un pape importante en la reparación y transcripción del DNA (Althaus, 1992). Por ejemplo, la poli-ADPribosilación de algunos factores de transcripción tales como la proteína de unión a TATA de complejo de la RNA polimerasa II, inhibe la transcripción genética (Oei et al, 1998). Se sabe que los taninos de plantas como el té verde inhiben la poli(ADP-ribosa) glicohidrolasa (PARG), una enzima que participa en la despolimerización de las moléculas de poli(ADP-ribosa) Esta inhibición suprime la expresión del gen del virus del tumor mamario murino (VTMM) inducida por glucocorticoides (Tsai et al, 1992). Sobre la base de estos antecedentes, Uchiumi y colaboradores (1996) decidieron estudiar el efecto del ácido tánico sobre la transcripción de una construcción que poseía el gen reportero cloranfenicol acetil transferasa (cat) fusionado a secuencias promotoras del LTR del VIH-1 (plásmido pHIVCAT). Para observar mejor el efecto inhibitorio, estos autores sometieron ciertas células T infectadas con VIH-1 a la acción del TPA (12-O-tetradecanoilforbol-13-acetato), ya que se sabe que induce la actividad promotora de las LTR. El TPA activa proteínas quinasas que fosforilan la proteína IκB citoplasmática liberando el factor de transcripción NF-κB. Este migra al núcleo en donde se puede unir a las LTR y promover la transcripción de los genes virales (Uchiumi et al, 1996; Luciw, 1996). Los resultados obtenidos por estos autores se muestran en la Figura 8, en donde se aprecia que el ácido tánico inhibió la transcripción de CAT inducida por TPA de manera dependiente de la concentración. En las células no tratadas con TPA el efecto no se observó puesto que los niveles basales de expresión de CAT fueron muy bajos. Para identificar que elementos del promotor del VIH respondían al ácido tánico se construyeron plásmidos con deleciones de 318 (pHCATVII) y 358 (pHCATVIII) pares de bases en la región LTR. Los plásmidos se incorporaron a células T que se trataron con TPA en presencia o en ausencia de ácido tánico. La medición de la actividad de CAT reveló que la carencia de una secuencia nucleotídica entre las posiciones 133 y 93 anulaba la inhibición de la transcripción causada por el ácido tánico (Figura 9). Ello indicó que los elementos sensibles a la acción del ácido tánico se encontraban dentro de la región suprimida de 40 pares de bases (Uchiumi et al, 1996). Debido a que las secuencias de unión del factor NF-κB se encontraban en dicha región, los mismos autores decidieron estudiar el efecto del ácido tánico sobre la transcripción del gen ca asociado a estas secuencias, ya que podría afectarse la expresión de otros genes. Observaron que el compuesto no inhibió la actividad de CAT, por lo que las regiones reconocidas por NF-κB no estaban involucradas en la inhibición de la transcripción mediada por el ácido tánico. De all concluyeron que el elemento responsable del efecto inhibitorio del ácido tánico era una secuencia de 30 pares de bases que se encontraba corriente arriba de los elementos de unión a NF-κB Esto condujo a Uchiumi y colaboradores (1996) a sugerir que el mecanismo de acción del ácido tánico en la disminución de la expresión a partir del LTR, ocurría mediante la inhibición de la acción de PARG. Se ha descrito que la poli-ADP-ribosilación de algunos factores transcripcionales conlleva a una disminución de la transcripción. La PARG se encarga de despolimerizar a dichos factores volviéndolos activos. Al parecer, en la expresión del DNA del VIH están involucrados factores asociados a ciertas regiones de las LTR que son susceptibles a la ADP ribosilación, por lo que requieren ser despolimerizados por PARG para promover la transcripción. Por lo tanto, la presencia del ácido tánico, al inhibir la PARG, impediría la despolimerización de dichos factores disminuyendo la expresión de los genes virales. Motivados por esta investigación y por otros trabajos que mostraban que los ligninos también inhibían la PARG, Uchiumi et al. (2003) decidieron estudiar el efecto de una serie de compuestos de plantas relacionados con los taninos y ligninos de estructura conocida. Para dichos estudio realizaron los mismos ensayos que en su trabajo de 1996, pero emplearon un gen reportero distinto: el de la luciferasa (luc). Esta es una enzima aislada de la especie de luciérnaga Photinus pyralis, que genera luz al monooxigenar un sustrato de luciferina. La intensidad de la luz que se genera es proporcional a la cantidad de enzima y puede medirse en un luminómetro (MGT, sin fecha). E gen luc se fusionó con las LTR del VIH-1 en un vector y fue transfectado a células T. De esta manera se observó que el ácido tánico, los elagitaninos (taninos hidrolizables), las cumarinas y las calconas (de la familia de los flavonoides) aislados de distintas plantas, inhibieron la actividad luciferasa inducida por TPA de manera dependiente de la concentración (Figura 10). Los elagitaninos, que mostraron efecto inhibitorio, compartieron una estructura circular compuesta por fenoles y sacáridos responsable de la supresión de la expresión de ciertos genes virales al actuar sobre PARG, por lo que este podría ser el mecanismo involucrado en la inhibición de la expresión de la luciferasa (Uchiumi et al, 2003). Por otro lado, a pesar de que el mecanismo de acción mediante el cual las calconas y las cumarinas ejercen su efecto inhibitorio no ha sido determinado, se ha sugerido que podría involucrar la unión selectiva a ciertos factores proteicos de transcripción o transducción, ya que compuestos de estructura similar no tuvieron efecto inhibitorio (Uchiumi et al, 2003). Los inhibidores de la integración del genoma viral: la proteína MAP 30. La ricina es una proteína que ha sido tradicionalmente conocida por ser extremadamente tóxica a inactivar los ribosomas e impedir la síntesis proteica celular. Se trata por lo tanto de una proteína inactivadora de los ribosomas (PIR) que es capaz de causar un 50 % de muerte celular a concentraciones por debajo de 1 ng/mL (Peumans, et al, 2001). El interés por el efecto citotóxico de la ricina llevó al aislamiento de proteínas funcionalmente relacionadas en una gran variedad de plantas. Una de ellas es la proteína MAP 30 aislada de Momordica charantia, una planta originaria de China cuyos extractos se han utilizado por siglos para el tratamiento de infecciones y tumores (Lee-Huang et al, 1995). A diferencia de la ricina, MAP 30 exhibe una toxicidad selectiva hacia células tumorales e infectadas con virus sin afectar las células normales, por lo que constituye una excelente candidata para la elaboración potencial de fármacos anticancerosos y antivirales. Los mecanismos de reconocimiento de las células alteradas por parte de MAP 30 aún no han sido establecidos (Wang et al, 1999). Las PIR ejercen su mecanismo inhibitorio gracias a su actividad RNA N-glicosidasa que consiste en la ruptura del enlace glicosídico N-C entre una adenina específica y su base nucleotídica. Esto ocurre al nivel de una adenosina situada en la posición 4324 de la subunidad 28 S del RNA ribosomal. Como consecuencia de esta deadenilación los ribosomas son inactivados y la síntesis proteica bloqueada. No obstante, varios indicios sugieren que el efecto anti-VIH de MAP30 no se debe a la deadenilación del rRNA, ya que la proteína es capaz de inhibir la infección por VIH-1 en células T y macrófagos a concentraciones que no muestran efecto sobre los ribosomas (Wang et al, 1999). Además, MAP 30 es capaz de inactivar topológicamente las LTR del DNA del VIH-1, haciéndolas insensibles a la acción de la DNA girasa, una topoisomerasa celular que interviene en el desenrollamiento de DNA durante los procesos de replicación y transcripción del DNA (Lee-Huang et al, 1995) Basándose en estas observaciones, Lee-Huang y colaboradores (1995) decidieron estudiar s también existía algún efecto de la proteína MAP 30 sobre la actividad de la integrasa viral. Para eso sintetizaron fragmentos de DNA marcados con fósforo radioactivo, con secuencias de 21 nucleótidos correspondientes a los extremos de las regiones U3 y U5 del las LTR del VIH-1 Estos fragmentos sirvieron de sustrato para la integrasa viral, cuyo primer paso catalítico consiste en realizar cortes escalados a nivel de los extremos 3’ del DNA de doble cadena viral (ver mecanismos de replicación del virus), suprimiendo específicamente el par de nucleótidos GT (Figura 11). Por lo tanto, la acción de la integrasa sobre los fragmentos sintéticos generaría oligonucleótidos de 19 bases que podrían separarse y visualizarse por electroforesis y autorradiografía (Lee-Huang et al, 1995). Los investigadores antes mencionados observaron que, al incubar la integrasa en un medio de reacción con los sustratos en presencia de MAP 30, hubo una inhibición en la generación de fragmentos escindidos con relación a los controles no expuestos a la proteína. Vieron que e efecto, tanto en los sustratos U5 como U3, fue proporcional a la concentración de proteína (Figura 11). Esto demostró que la proteína impedía el procesamiento inicial de las LTR por parte de la integrasa. Para estudiar sí la MAP 30 también era capaz de impedir el proceso de transferencia de hebras entre el genoma viral y el genoma del hospedador catalizado por la integrasa, estos autores incubaron la enzima junto a diversas concentraciones de proteína en presencia de los sustratos U5 y U3 carentes de los nucleótidos terminales y de un plásmido receptor. Bajo estas condiciones cualquier inhibición detectada se debería a la interferencia de MAP 30 sobre la transferencia de hebras y no sobre la escisión nucleotídica. Debido a que los sustratos U5 y U3 se encontraban marcados radioactivamente, se pudo hacer un seguimiento de la integración mediante electroforesis y autorradiografía. Conclusiones
CC50: Concentración de compuestos que causa una disminución de un 50% Referencias
NOTA: Toda la información que se brinda en este artículo es de carácter investigativo y con fines académicos y de actualización para estudiantes y profesionales de la salud. En ningún caso es de carácter general ni sustituye el asesoramiento de un médico. Ante cualquier duda que pueda tener sobre su estado de salud, consulte con su médico o especialista. Copyright 2007 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela The following images related to this document are available:Photo images[va07012f4.jpg] [va07012t1.jpg] [va07012f12.jpg] [va07012f9.jpg] [va07012f5.jpg] [va07012f6.jpg] [va07012f11.jpg] [va07012f8.jpg] [va07012f1.jpg] [va07012f7.jpg] [va07012t4.jpg] [va07012f3.jpg] [va07012t2.jpg] [va07012f2.jpg] [va07012f10.jpg] [va07012t3.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}