 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
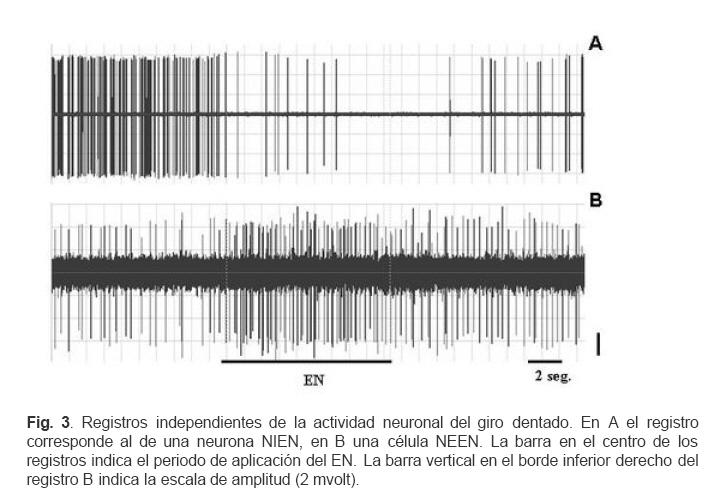
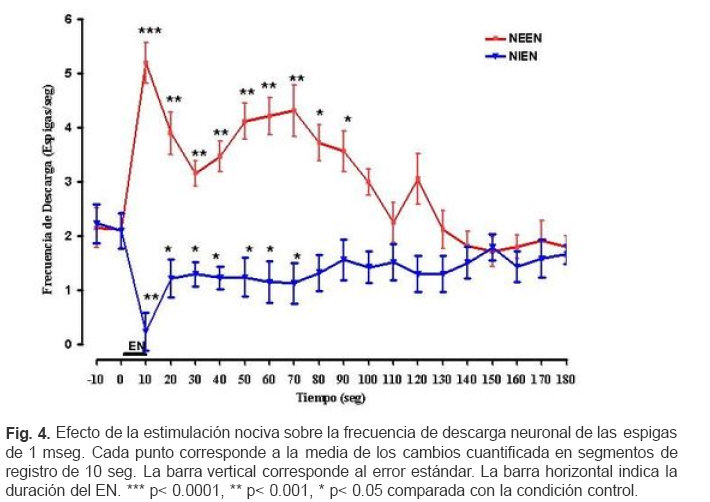
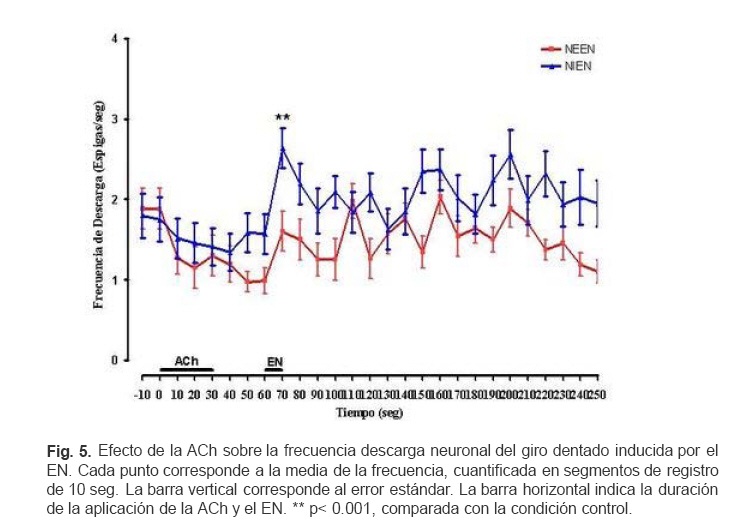
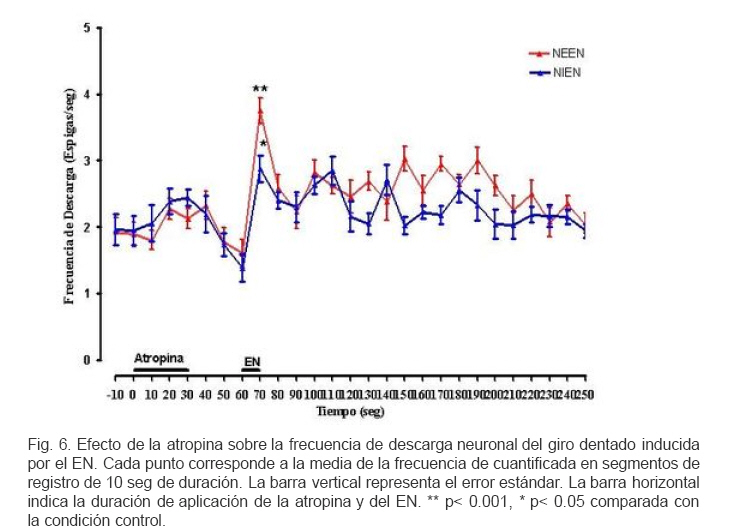
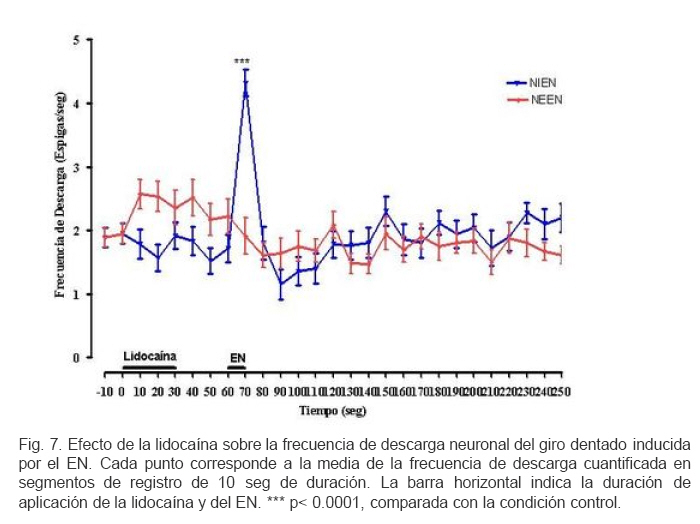
VITAE Academia Biomédica Digital, Número, No. 37, Enero-Marzo, 2009 Efecto de la lidocaina y la acetilcolina sobre la actividad neuronal del giro dentado inducida por estimulacion nociva Effect of lidocaine and acetylcholine on the neuronal activity of the dentate gyrus induced by noxious peripheral stimuli. Rafael Salas 1 1rasalas@cantv.net Cátedra de Fisiología, Escuela de Bioanálisis Facultad de Medicina, Universidad Central de Venezuela Fecha de recepción: 03/03/2009 Fecha de aceptación: 12/04/2009 Code Number: va09003 El giro dentado es la entrada principal de la formación hipocámpica, y destino de la mayoría de las aferencias de la corteza entorrinal que terminan haciendo contacto las células granulosas y que transportan información de múltiples modalidades sensoriales acerca del mundo exterior. Es de suponer que parte de la información nociceptiva que llega a la formación hipocámpica es procesada en el giro dentado, por lo que, cualquier cambio en su actividad neuronal, alterará la respuesta provocada por un estimulo nocivo (EN). Nosotros estudiamos los cambios en la descarga neuronal del giro dentado, inducidos por la aplicación de un EN, antes y después del pre-tratamiento con Acetilcolina (ACh), Atropina y Lidocaína, usando registros electrofisiológico extracelulares convencionales, en ratas anestesiadas con uretano. Nuestros hallazgos sugieren, que en el giro dentado, al menos en ratas anestesiadas con uretano, el procesamiento de la información nociceptiva, involucra la modulación de poblaciones neuronales que descargan en forma opuesta. Este proceso parece estar mediado en parte, por un mecanismo colinérgico, en el que podrían participar al menos dos tipos de receptores colinérgicos. Uno de ellos muscarínico, que produce un efecto inhibitorio en una determina población neuronal, y puede ser bloqueado por su antagonista no selectivo atropina, y el otro receptor sería no muscarínico. Por otra parte, el efecto inducido por inyección local de lidocaína en el giro dentado, parece depender de estos mecanismos colinérgicos. Estas observaciones muestran evidencias, del papel crítico que juega el giro dentado en la percepción del dolor. Palabras Claves:Giro dentado; ACh; Lidocaina; Nocicepción Abstract The dentate gyrus is the main input to the hippocampal formation and target of the majority of entorhinal cortex afferents that make contact on granule cell dendrites and convey sensory information from multiple modalities. Presumably nociceptive information reaching the hippocampal formation is processed in the dentate gyrus, and changes in neuronal activity, could reflect the the response induced by a noxious stimulus (EN). We studied the changes in neuronal discharge of gyrus dentate, induced by the application of an EN, before and after pretreatment with acetylcholine (ACh), atropine and lidocaine, using conventional extracellular electrophysiological methods in rats anesthetized with urethane. Our findings suggest that in the dentate gyrus, the nociceptive information processing involves the modulation of neuronal populations that discharge in opposite directions. This process seems to be mediated in part by a cholinergic mechanism, which could involve at least two types of cholinergic receptors. One muscarinic, producing an inhibitory effect in a given neuronal population, and can be blocked by the non-selective antagonist atropine and another non-muscarinic receptor. Moreover, the effect induced by local injection of lidocaine into the dentate gyrus, seems to depend on cholinergic mechanisms. These observations provide evidence of the critical role played by the dentate gyrus in pain perception. Key Word: Dentate gyrus; ACh; Lidocaine; Nociception Introducción Una de las estructuras del sistema límbico que parece tener un rol importante en el componente afectivo-motivacional de la conducta de un individuo ante un estímulo nocivo, es la Formación Hipocámpica (11). Lesiones de la corteza entorrinal o el hipocampo en animales de experimentación, incrementan el umbral de vocalización ante un estímulo nocivo (2,8), además estas lesiones reducen las conductas agresivas del animal (6,7). El procesamiento de la información nociceptiva a nivel de la formación hipocámpica podría estar siendo mediada en parte por el sistema colinérgico que se origina en el septum medial y proyecta sobre el giro dentado. Se ha observado que la respuesta de las células granulosas del giro dentado a la estimulación del septum medial es bloqueada por atropina, mientras que la evocada por estimulación de las fibras perforantes lo es por ácido glutámico dietileter (20). Por otra parte, la aplicación de estímulos nocivos incrementa la actividad de algunas neuronas del área de la banda diagonal del septum medial, no así con estímulos no nocivos; estas neuronas envían fibras colinérgicas hacia la Formación Hipocámpica (5). Por otra parte, la inyección iontoforética de atropina en el hipocampo o en el giro dentado, reduce las descargas de las células theta, pero no parece modificar la actividad de las denominadas células complejas (18). Estudios conductuales han mostrado que la inyección de lidocaína en el giro dentado de ratas no entrenadas, reduce la conducta relacionada con el dolor provocado por la inyección subcutánea de formalina en la pata del animal (10). Sí el giro dentado es el primer estadio del circuito neuronal intrahipocámpico y blanco principal de la mayoría de las aferencias de la corteza entorrinal, de aferentes subcorticales y commisurales, que transportan información de múltiples modalidades sensoriales acerca del mundo exterior, y terminan haciendo contacto con interneuronas o con las células granulosas (15, 9, 21), es de suponer que parte de la información nociceptiva que llega a la formación hipocámpica es procesada en el giro dentado, por lo que, cualquier cambio inducido en su actividad neuronal, alterará la descarga neuronal evocada por el estimulo nocivo. En este contexto nos propusimos estudiar por medio de registros electrofisiológicos convencionales, los cambios en descarga neuronal y en el ritmo theta, en el giro dentado inducidos por estimulación nociva de la cola, bajo el efecto de inyección local de ACh, Atropina y Lidocaina en ratas anestesiadas con uretano. Materiales y métodos Los sujetos experimentales fueron 30 ratas machos de la cepa Sprague Dawley de peso entre 250-300 gramos, mantenidos en un ritmo luz:oscuridad 12:12 horas y alimentados ad libitum. Los animales fueron anestesiados con una solución de uretano (Ethyl Carbamate), a una dosis de 1 gr/Kgr de peso, vía intraperitoneal. Una vez el animal profundamente dormido, lo cual se comprobó por la arreflexia corneal, se colocó en un aparato estereotáxico con fijación craneana, luego se procedió a realizar una craneotomía de acuerdo a las coordenadas del atlas para el cerebro de ratas (12), usando como punto de referencia el Bregma. Se realizaron registros independientes con dos tipos de microelectrodos: Microelectrodos de vidrio borosilicato, llenos con una solución 2.5 M de azul de pontamina, y cuya resistencia medida con un voltímetro digital convencional fue de 4 a 10 MΩ. Microelectrodos de Tungsteno (W.P.I®) de 1MΩ de resistencia adosado a una microcánula de acero (ø 100 μm) alineados punta con punta. Con ayuda de un micromanipulador se colocaban los microelectrodos en la zona de registro en el giro dentado de la Formación Hipocámpica, de acuerdo a las siguientes coordenadas: AP = 3.3 – 4.3; L = 1-2 DV = 3 – 4. Previo a la colocación del microelectrodo de metal en la zona de registro en el giro dentado, la cánula metálica y el tubo plástico que la conectaba a una inyectadora (Hamilton®) de 1 μL, fueron llenados y purgados con la solución de la droga a inyectar. Los microelectrodos, fueron conectados a un pre-amplificador (Mentor® N-950). La señal obtenida en forma independiente de cada giro dentado, fue amplificada por medio de un amplificador (Tektronix®, AM502), y registrada en forma continua a través de una tarjeta genérica de sonido de dos canales con una velocidad de muestreo de 48 Kc/s conectada a una computadora Pentium que disponía de un programa editor de señales de audio (CoolEdit 1.5 and 95, syntrillium). Típicamente se registró 1 minuto de actividad control, luego se aplicó el estímulo nocivo (EN) el cual consistió en introducir la parte final de la cola en un vaso con agua a 54 oC por 10 segundos, habiendo transcurrido de 3 a 4 minutos se inyectó 0.2 µL de una de las drogas en la zona de registro del giro dentado, el siguiente EN se aplicó 30 segundos después de haber terminado la inyección. Todas las drogas fueron preparadas en solución fisiológica 0,9 % NaCl, manteniéndose la isosmolaridad de las soluciones finales. Solución de sulfato de Atropina (Sigma) (PM: 676.8) 0,5 mgrs/mL; Solución de Cloruro de Acetilcolina (Sigma), ACh; (PM: 201.7) 1 mgr/mL; Solución de Lidocaina® al 2%. Una vez concluido cada experimento se procedió a marcar la zona de registro, haciendo pasar una corriente a través de los microelectrodos. En el caso de los microelectrodos de vidrio llenos con azul de pontamina (2.5 M), se pasó una corriente negativa de 50 μAmp., durante 10 min. Para determinar el área de inyección, en algunos casos una vez concluido un registro de la actividad neuronal, se retiró el microelectrodo con su cánula y se procedió a llenar la cánula con azul de pontamina, luego se realizó un nuevo registro utilizando las mismas coordenadas del estereotáxico del registro anterior. Una vez colocado el microelectrodo en el giro dentado, y verificada la descarga neuronal, se inyectó 0.2 µL de azul pontamina. Luego se extrajo el cerebro y se fijó en formaldehído al 10 %. Posteriormente se realizaron cortes de 100-150 μm utilizando un Críostato, cada corte así obtenido, fue observado bajo microscopio de luz para ubicar la marca dejada por el colorante. Cada señal adquirida fue filtrada con un filtro pasa banda de 0.3 a 5 KHz. Se evaluaron aquellas espigas con la misma forma y duración de 1 mseg, utilizando el programa g-prime. La duración se determinó fijando en primer lugar un nivel basal de referencia, el inicio de la espiga se tomo justo al borde donde comenzaba la deflexión negativa de la espiga, el final cuando la espiga alcanzaba nuevamente el nivel basal. La actividad neuronal se expresó como la frecuencia de descarga en espigas/seg. La media y la desviación estándar de los cambios fueron cuantificados y graficados utilizando el programa GraphPad Prisma 4. Los datos obtenidos se compararon aplicando la prueba estadística “t de Student” para muestras no pareadas. Se consideraron estadísticamente significativos cuando el valor de p fue menor de 0,05. Resultados En un total de 30 animales experimentales, se registraron 120 células. El análisis histológico confirmó que la punta de los microelectrodos penetró hasta la región apropiada en el giro dentado. La figura 1 muestra una fotomicrografía de un corte coronal de cerebro de rata, donde se puede apreciar la marca dejada por la inyección de azul de pontamina en la región del giro dentado en el hemisferio derecho. Para el control del posible efecto de la difusión de la droga sobre algunas estructuras mas distales se realizaron algunos ensayos inyectando 0.2 uL de azul de pontamina en las mismas áreas de registro. La figura 2 muestra una fotomicrografía de un corte coronal de cerebro de rata, donde se puede apreciar las marcas dejada por la inyección de azul de pontamina en la región del giro dentado, representativas de los sitios de inyección de la droga. La aplicación de un EN, produjo efectos opuestos en la descarga neuronal unitaria de aquellas poblaciones neuronales que descargan espigas de la misma forma y duración. En unas neuronas se redujo la frecuencia de descarga de las espigas. En adelante llamaremos a estas neuronas que se inhiben por EN (NIEN). Otras poblaciones neuronales incrementaron su descarga, neuronas que se excitan por el EN (NEEN). La figura 3 muestra dos segmentos de registros continuos independientes de la descarga neuronal del giro dentado, en los que se pueden observar los cambios inducidos en la descarga neuronal por la aplicación del EN. El análisis estadístico de los registros de la actividad de 40 neuronas NEEN, indica que en promedio la frecuencia descarga se incrementa de manera significativa de 2.15 (± 0.3) espigas/ seg, en condición control a 5.2 (±0.36) espigas/seg durante la aplicación del EN, y permanece descargando a una frecuencia mayor a 2.15 espigas/seg hasta los 70 minutos después del EN. Mientras que en 43 células NIEN, su actividad redujo significativamente a 0.24 (± 0.35) durante la aplicación del estimulo. En la figura 4 se muestra el curso temporal de los cambios observados en la descarga de ambas poblaciones neuronales inducidos por la aplicación del EN. Para determinar el posible efecto de la solución salina como agente solvente, sobre la actividad neuronal, se realizaron 6 experimentos donde se evaluaron los cambios inducidos por la inyección local de solución salina al 0.9 % en la zona de registro en el giro dentado. El análisis estadístico de la frecuencia de descarga de 8 neuronas NEEN, y de 8 NIEN indica que no existen diferencias estadísticamente significativas entre los cambios inducidos por el EN previo a la inyección de solución salina y los observados después de su inyección. El análisis estadístico de los registros de la actividad de 13 neuronas NEEN, cuyas descargas fueron clasificadas como espigas de 1 mseg, muestra que después de la aplicación de ACh, el EN no induce cambios significativos en la frecuencia de la descarga de este tipo de espiga, en comparación con su condición control. Sin embargo, en 14 neuronas NIEN, el EN provocó un incremento significativo en la descarga neuronal de (1.8 ± 0.27) a (2.64 ± 0.25) espigas/seg, cuando se aplicó después de la inyección de ACh. La figura 5 muestra el curso temporal de los cambios observados en la descarga neuronal inducidos por la aplicación del EN bajo el efecto de la ACh. El análisis estadístico de la evaluación de los registro de la descarga de 11 neuronas NEEN, muestra que después de la aplicación de atropina, el estimulo nocivo provocó incrementos significativos en su frecuencia de descarga de (1.89 ± 0.18) a (3.75 ± 0.19), mientras que en 12 neuronas NIEN, aumentó de (1.95 ± 0.22) a (2.88 ± 0.20) espigas/seg. La figura 6 muestra el curso temporal de los cambios observados en el análisis estadístico de la descarga neuronal inducidos por la aplicación del estimulo nocivo en la cola, después de haber inyectado la atropina. El análisis estadístico del registro de la actividad de 9 neuronas NIEN, muestra que después de la aplicación de lidocaína la actividad evocada por el EN, aumentó en forma significativa de (2.12 ± 0.32) a (4.32 ± 0.5) espigas/seg. Mientras que en 8 neuronas NEEN, el estimulo nocivo redujo significativamente su descarga a 1 espiga /seg. La figura 7 muestra el curso temporal de los cambios observados en la descarga neuronal inducidos por la aplicación del estimulo nocivo en la cola después de haber inyectado la lidocaína. Discusión Los datos obtenidos a partir del marcaje histológico indican, por una parte, que los registros de la actividad neuronal estuvieron circunscritos a la región del giro dentado, y que los cambios observados en su actividad celular no fueron debido a la infusión de la solución salina, ni a la difusión de la droga a estructuras adyacentes a la Formación Hipocámpica. En este estudio, se muestran evidencias de la activación de dos tipos de poblaciones neuronales en el giro dentado, que responden en forma opuesta al estimulo nocivo, células que se excitan ante el EN, mientras que otras se inhiben. Estos resultados sugieren que las proyecciones sobre el giro dentado, tanto de origen septal, como las que provienen de la corteza entorrinal, o de otras estructuras subcorticales, podrían estar modulando la descarga neuronal. Estos aferentes extrínsecos, parecen ejercer un poderoso control sobre la actividad eléctrica del giro dentado, posiblemente estos aferentes terminan en una población neuronal relativamente pequeña, pero capaz de controlar grandes áreas de la formación hipocámpica. La ACh provocó cambios opuestos en la descarga neuronal inducida por el EN. La población de neuronas denominadas por nosotros como NIEN, incrementaron su descarga, mientras que redujo la actividad de las NEEN. Evidencias experimentales permiten sugerir que, a nivel de la formación hipocámpica, coexisten varios tipos de receptores para la ACh. La aplicación microiontoforética en el hipocampo de ACh o Carbacol agentes que actúan tanto en sitios nicotínicos como muscarínicos, producen excitación de las células piramidales. Los antagonistas nicotínicos, dihidroxi-β-eritroidina y galamina bloquean la excitación de estas células, producidas por la ACh (1, 17). Otros resultados experimentales demuestran que la activación del receptor nicotínico α7 (nAChRs), por la administración i.c.v. de colina como precursor del neurotransmisor ACh, evoca un efecto antinociceptivo tanto en modelos de dolor agudo como en procesos inflamatorios. Por otra parte los antagonistas de este receptor como la α-bungarotoxina bloquean su efecto antinociceptivo (19). En las células NEEN bajo el efecto de atropina, la aplicación del EN indujo un cambio opuesto al observado después de la inyección de la ACh, mientras que sobre las células NIEN la atropina no parece modificar los cambios inducidos por el EN. Estos resultados podrían explicarse sobre la base de la existencia de, al menos, dos tipos de receptores colinérgicos en el giro dentado. Uno de ellos es muscarínico, produce un efecto inhibitorio, y puede ser bloqueado por su antagonista no selectivo atropina, el otro receptor sería tal vez colinérgico no muscarínico. Por ello, cuando la atropina bloquea el receptor muscarínico, se incrementaría la disponibilidad de ACh, pudiendo actuar sobre el receptor no muscarínico, este hecho justificaría el incremento que se observa el la descarga de las células NIEN. En base a estos planteamientos nuestros resultados sugieren que en el procesamiento de la información nociceptiva a nivel del giro dentado, participan mecanismos colinérgicos a través de diferentes tipos de receptores. La lidocaina es un anestésico local que induce un bloqueo reversible de la generación y conducción del impulso nervioso por reducción de la permeabilidad del ión sodio (14). Estudios conductuales demuestran que la inyección de lidocaína en el giro dentado produce un efecto analgésico, ante la inyección subcutánea de formalina. Estos resultados confirman la participación de la formación hipocámpica en la estructura neural implicada en la percepción de dolor (10). Nosotros evaluamos el efecto de la inyección local de lidocaína en giro dentado, sobre la actividad neuronal evocada por el EN. La frecuencia de la descarga neuronal de las célula NIEN se incrementó significativamente por la aplicación del EN, mientras que la descarga de las células NEEN no parece modificarse. Estos hallazgos, sugieren que el efecto de la lidocaína a nivel del giro dentado implica un mecanismo que activaría a más de un tipo de receptor colinégico. Uno muscarínico como lo planteamos anteriormente, y otro no muscarínico. Basados en este análisis podríamos sugerir que la ACh, activa circuitos neuronales tanto excitatorios como inhibitorios., Sería el interjuego entre estas dos corrientes lo que determinaría el procesamiento de la información nociceptiva en el giro dentado. Varios reportes apoyan el rol de la ACh en la modulación de la transmisión de la información nociceptiva. Se ha demostrado que agentes antinociceptivos, tales como la morfina o la clonidina, son capaces de producir un incremento en la liberación de ACh en la médula espinal (4, 7, 3; 16). Mecanismos similares serían activados por las drogas antiinflamatorias no esteriodeas (NSAIDs), como diclofenac, piroxican, y paracetamol, ya que su efecto antinociceptivo es antagonizado por la atropina (13). Conclusiones: La información nociceptiva induce en el giro dentado, la activación de al menos dos tipos de poblaciones neuronales, que descargan en forma opuesta. Neuronas colinégicas, y receptores colinérgicos muscarínicos en el giro dentado participan en el procesamiento de la información nociceptiva. El efecto antinoceceptivo inducido por la lidocaina, parece estar mediado por neuronas colinérgicas, a través de la activación de receptores muscarínicos y no muscarínicos. Agradecimientos: Este trabajo fue parcialmente financiado por el CDCH y la Coordinación de Investigación de la Facultad de Medicina de la Universidad Central de Venezuela. Referencias
NOTA: Toda la información que se brinda en este artículo es de carácter investigativo y con fines académicos y de actualización para estudiantes y profesionales de la salud. En ningún caso es de carácter general ni sustituye el asesoramiento de un médico. Ante cualquier duda que pueda tener sobre su estado de salud, consulte con su médico o especialista. Copyright 2009 - Centro de Análisis de Imágenes Biomédicas Computarizadas CAIBCO, Instituto de Medicina Tropical – Facultad de Medicina, Universidad Central de Venezuela The following images related to this document are available:Photo images[va09003f7.jpg] [va09003f1.jpg] [va09003f5.jpg] [va09003f6.jpg] [va09003f3.jpg] [va09003f4.jpg] [va09003f2.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}