 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Academia Biomedica Digital, Vol. 0, No. 42, , 2010 Hibridación Genómica Comparada: Una revisión de los estudios sobre las alteraciones cromosómicas en la neoplasia intraepitelial y el carcinoma cervical durante la infección con el VPH. Comparative Genomic Hibridization: a review of studies of chromosomal alterations in cervical intraepithelial neoplasia and carcinoma of the ceervix during VPH infection. Julia Molina Barrios1 , Asmiria Arenas 2, Eduardo Blasco Olaetxea 3, Jorge García Tamayo 4
1 Laboratorio
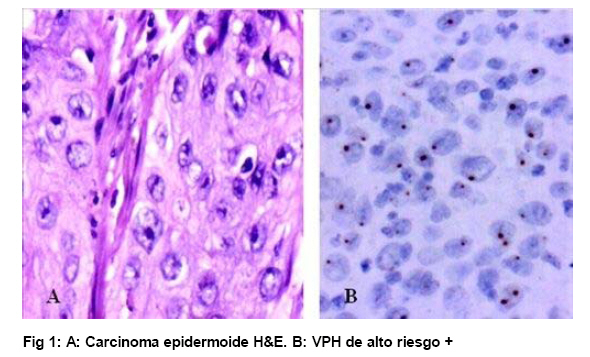
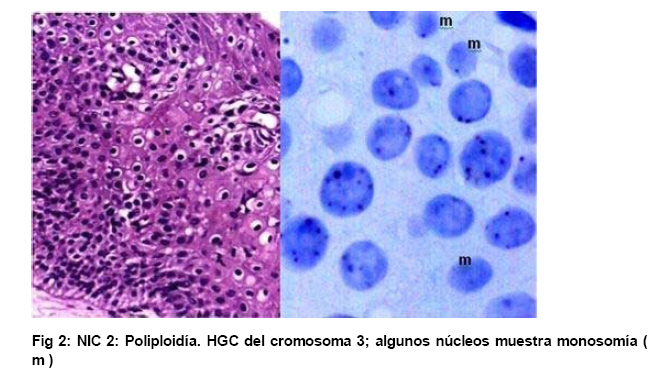
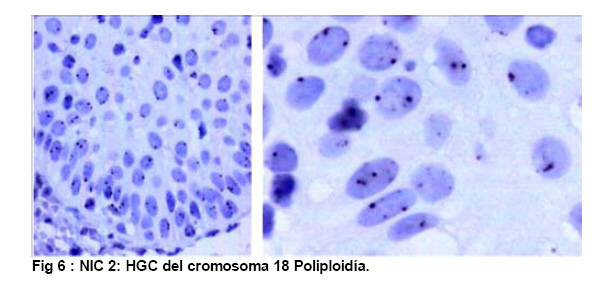
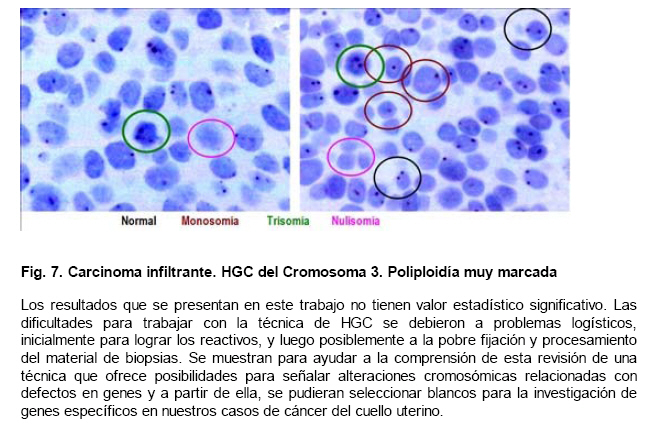
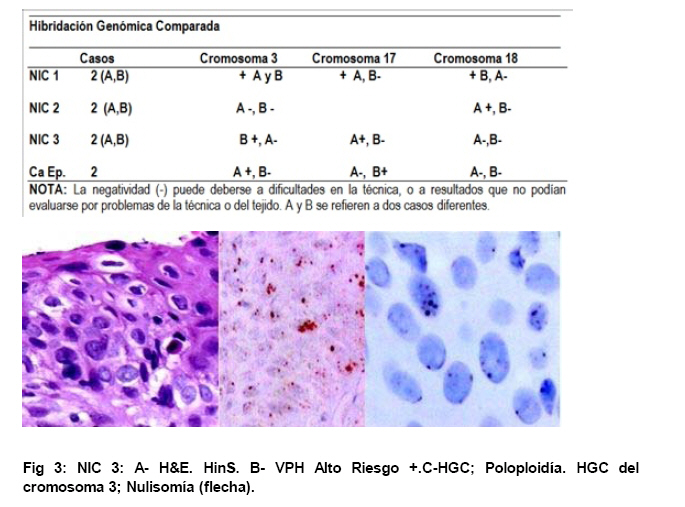
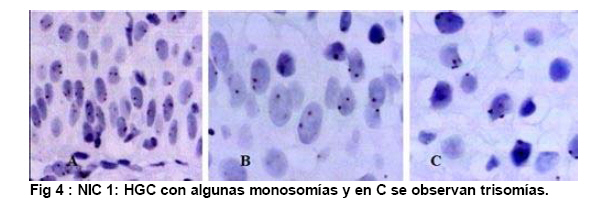
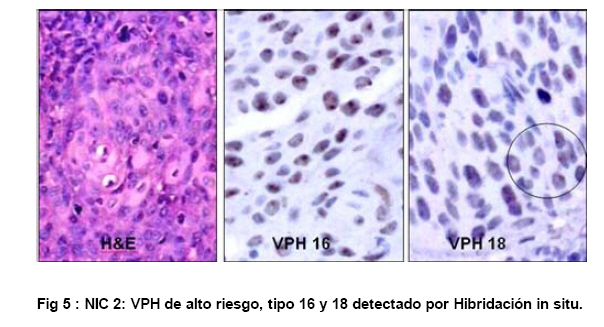
de Patología Molecular, Maracaibo, Venezuela Fecha de recepción: 28/05/2010 Code Number: va10010 La técnica de Hibridación in situ cromogénica se aplicó para hacer Hibridación Genómica Comparada con sondas para los cromosomas 3, 17 y 18 en algunos casos de neoplasia intraepitelial cervical (NIC1 a 3) y carcinoma espidermoide del cuello uterino. Se demuestran aneusomías cromosómicas en las diversas etapas de NIC y en el carcinoma, con las tres sondas de los cromosomas que se investigaron. Se señala la importancia de este tipo de investigación para mostrar alteraciones cromosómicas tempranas. Se revisa la literatura sobre las investigaciones descritas en relación con alteraciones de los diferentes cromosomas durante la infección con el VPH y asociadas al desarrollo de la neoplasia cervical. Palabras Claves: Hibridación Genómica Comparada, Alteraciones cromosómicas, Poliploidía, Aneusomías, VPH. Abstract Using in situ chromogenic hibridization, Comparative Genomic Hibridization with chromosomes 3,17 and 18 was examined in some cases of CIN (1 to 3) and squamous cell carcinoma of the cervix. Nuclei Aneusomy, and chromosomal abnormalities detected by CGH were related to loss of heterocigosys and chromosomal instability during carcinogenesis related to HPV infection of the cervix. Introducción Las observaciones hechas en Italia en el siglo antepasado (1842) sobre la aparición de cáncer del cuello uterino con mayor frecuencia en mujeres casadas y raramente visto en monjas, fue la primera sugerencia hecha sobre la posibilidad de que un agente infeccioso fuese el responsable del cáncer cervical(1). En 1971, en el VIII Congreso Latinoamericano de Patología, J.Nágel y E.Casale, en Maracaibo, Venezuela, demostraron con el microscopio electrónico la presencia de partículas virales en unas lesiones como manchas rosadas en el cuello uterino.A la luz de estos hallazgos, José Trinidad Núñez Montiel y Jorge García Tamayo, presentaron dos años después en el siguiente, IX Congreso Latinoamericano de Patología en Mérida, Yucatán, México (1973) en un par de trabajos libres, con evidencias inquívocas de la apariencia colposcópica, la histopatología y los cambios ultraestructurales con la identificación por tinción negativa del virus del papiloma humano en lesiones de displasia y de cáncer del cuello uterino (2,3). Los cambios citológicos de la infección con este virus fueron descritos posteriormente por Meisels y Fortín (4) y por Puroloa y Savia (5) en 1976. El año 1978 se hizo una publicación reinformando sobre nuestros hallazgos en una revista médica venezolana (6), el cual precedió algunos otros trabajos (7,8,9) hasta que Harald zur Hausen demostró la presencia de las secuencias de ADN viral en el cáncer del cérvix (10). ZurHausen unos años más tarde con algunos otros investigadores estudiaron la relación entre diversos tipos de VPH y la neoplasia cervical (11,12,13), hasta quedar firmemente establecida la relación entre el cáncer cervical y el VPH de los tipos 16 y 18 (14). Con nuevos métodos de biología molecular, Nubia Muñoz y sus colaboradores en la Agencia Internacional para Investigación sobre el Cáncer (IARC) en Francia, han concluido que en un 99.7% de todos los casos de cáncer cervical, el VPH es la causa necesaria para que estos tumores se hayan desarrollado (15 ). Diversas investigaciones en las células del epitelio del cuello uterino, han demostrado alteraciones cromosómicas, particularmente tetrasomías, no obstante, se ha descrito que estas no se observan en todas las lesiones de neoplasia intraepitelial cervical (NIC) y de carcinoma escamoso del cuello uterino y que ellas tampoco se relacionan con un tipo específico de VPH (16,17). Se ha señalado que los queratinocitos sometidos a cambios en el microambiente de la interfase epitelio-mesénquima, soportan influencias extracelulares que conducen a modificaciones fenotípicas en el epitelio y como estos cambios se pueden correlacionar con las etapas de transición de NIC1 a NIC3 y a carcinoma escamoso (18,19). Hemos propuesto que el análisis de la expresión del perfil génico del epitelio cervical en la infección por el VPH es importante para ofrecer claves que permitan entender mejor la evolución y el pronóstico del cáncer del cuello uterino (20) . En este trabajo, utilizando el material de algunas biopsias de cuello uterino con NIC y carcinoma infiltrante, mostramos la infección con VPH de alto riesgo, y utilizando la Hibridación Genómica Comparada señalamos algunas evidencias de alteraciones genéticas en los cromosomas 3, 17 y 18 con la intención de destacar la importancia de esta técnica en la detección de alteraciones descritas previamente en genes involucrados en la carcinogénesis del cuello uterino. Estos casos fueron examinados paralelamente para VPH de Amplio Espectro y de Alto Riesgo. Materiales y Métodos Las muestras de biopsias de cérvix, fueron obtenidas de material del Laboratorio de Patología Molecular de la ciudad de Maracaibo, Venezuela. El material del cuello uterino correspondió a muestras seleccionadas entre 70 casos de NIC en estadios 1, 2, y 3 y de carcinoma epidermoide infiltrante, provenientes del Laboratorio de Patología BIMECA de la ciudad de Mérida. Las muestras se fijaron en formol tamponado al 10%, se deshidrataron y se incluyeron en parafina. Se examinaron histológicamente los casos y se seleccionaron 8 biopsias de casos de NIC1(2), NIC2 (2), NIC3 (2) y de carcinoma epidermoide infiltrante (2) en el Laboratorio de Patología Molecular de Maracaibo. En un microtomo de rotación, se realizaron cortes de los bloques de parafina para practicar en ellos la técnica de Hibridación Genómica Comparada (HGC) utilizando sondas centroméricas para los cromosomas 3, 17 y 18. El procedimiento de hibridación in situ cromogénica (HiSC) se realizó en los cortes de dos micras de espesor, colocados en una solución recuperadora precalentada a 98°C durante40 minutos. Después de lavarlos con agua desionizada se hizo una digestión breve con Proteinasa K 1/8.000. Los cortes en el agua desionizada fueron deshidratados con etanol, y se secaron al aire. Se aplicaron 15 microlitros de las sondas centroméricas correspondientes a los cromosomas 3, 17 y 18 ( Zymed SP.T-Light ® ) y se colocaron en un hibridizador durante 5 minutos a 95°C para su desnaturalización. El proceso de hibridación se realizó durante 14 horas. Los cortes se lavaron con PBS y se bloqueó la peroxidasa endógena con H2O2-metanol. Tras un nuevo lavado, se hizo un bloqueo de proteínas ( CAS-LOCK TM) por 10 minutos y la inmunodetección se hizo con un polímero de Estreptavidina por 30 min. Los cortes fueron lavados nuevamente, se colocaron en DAB por 30min, se lavaron con agua, se deshidrataron en alcoholes se contrastaron con hematoxilina, y fueron deshidratados y montados con medio de montaje. La observación y el análisis se realizó en un microscopio de luz. Estudiamos la presencia de infección genital con el virus del papiloma humano (VPH) por hibridación in situ utilizando sondas biotiniladas de ADN específicas para VPH, WS (de amplio espectro: Wide Spectrum HPV Biotinylated DNA Probe-Código Y1404 de DAKO ) que comprende los virus tipo 6,11, 16, 18, 31,33,35, 45, 51y 52 ; y AR ( de alto riesgo: The GenPoint™ HPV Probe-Código Y1443 de DAKO ) con los virus tipo 16,18, 31,33,35,39, 45, 51, 52, 56, 58, 59 y 68. Le dimos especial atención a las características del marcaje observado entre las dos sondas específicas WS y AR. Las señales de la Biotina se demostraron con el complejo primario de estreptavidina-peroxidasa, tiamina-biotinilada y secundariamente estreptavidina-peroxidasa (Sistema GenPoint™ para amplificación de la señal -DAKO). La distribución del marcaje en las células de los distintos estratos del epitelio se examinó en base a la apariencia histológica, el grado de la neoplasia intraepitelial cervical presente (NIC) o las características del carcinoma en casos francamente neoplásicos. Resultados Las ocho biopsias seleccionadas para estudiarlas simultáneamente con hibridación in situ cromogénica con VPH y con Hibridación Genómica Comparada utilizando sondas para los cromosomas 3, 17 y 18 correspondieron a dos casos de NIC1, dos casos de NIC2, dos casos de NIC3 y 2 muestras de de carcinoma epidermoide infiltrante. Todos los casos fueron examinados para VPH de Amplio espectro (WS) y de Alto riesgo (AR). Resultando positivos para WS todos y para AR un caso con NIC2, los dos casos NIC3 y los carcinomas. En los casos de NIC3 y carcinoma escamoso se examinaron los virus 16, 18 y 33,35, con positividad para VPH 16 en los casos de carcinoma y en un NIC3, y para 16 y 18 en uno de los casos de NIC3. Los resultados que se presentan en este trabajo no tienen valor estadístico significativo. Las dificultades para trabajar con la técnica de HGC se debieron a problemas logísticos, inicialmente para lograr los reactivos, y luego posiblemente a la pobre fijación y procesamiento del material de biopsias. Se muestran para ayudar a la comprensión de esta revisión de una técnica que ofrece posibilidades para señalar alteraciones cromosómicas relacionadas con defectos en genes y a partir de ella, se pudieran seleccionar blancos para la investigación de genes específicos en nuestros casos de cáncer del cuello uterino. Discusión En el fenómeno de la carcinogénesis cervical, además de la infección por el VPH de alto riesgo, parecen jugar un rol muy importante algunas alteraciones en genes supresores de tumores (11,21). Estudios sobre la pérdida de la heterocigosis en la neoplasia cervical ha demostrado pérdida de determinadas regiones cromosómicas, particularmente 3p, 4p, 5, 6, 11q y 17p, alteraciones estas que parece implicar a genes supresores de tumores (22,23,24,25). El análisis de la localización de los genes frecuentemente involucrados en estos cambios ha demostrado a TGM 4 y MLH1 en 3p, HLA, WAF1 y TNF en 6p, STMY, PGL, NCAM y CD3D en 11q, e igualmente los genes CRK, ABR y HIC1 en 17p (26). Hace varios años se describieron aneusomías cromosómicas en NIC y en el carcinoma cervical (26) y se sabía que éstas afectaban particularmente a los cromosomas 3, 7 y X (27). Investigaciones posteriores han enfatizado la relación entre éstas alteraciones cromosómicas y la inestabilidad genética, y se describió la presencia de aneusomías en el cromosoma 1, con micronúcleos, y la existencia de cromosomas ectópicos en casos de NIC (28). Se ha descrito que el 93% de los carcinomas epidermoides y adenocarcinomas del cuello ueterino mostraron pérdidas en diversos locus del cromosoma 4, la mayoría de ellos en los brazos largos; en estos casos, la detección del VPH 16/18 fue paralela a los resultados que mostraron una predominancia de estas alteraciones en los adenocarcinomas (82%) al compararlas con la del carcinoma epidermoide (31%), sin embargo, la prevalencia de la infección por VPH fue similar (85% en ADC y 73% en carcinoma epidermoide), lo cual refrenda la importancia de la participación viral en la progresión de la neoplasia. Las deleciónes del cromosoma 4 parecen ser frecuentes en el cáncer cervical y las diferencias antes señaladas posiblemente se deben a diferentes localizaciones de las regiones génicas afectadas en este cromosoma (22,29). En 23 de 31 lesiones NIC1 a NIC3 y carcinoma infiltrante del cérvix positivas a VPH 16 se observaron alteraciones cromosómicas, tetrasomías en 9 casos, aneusomías y polisomías en 8 de 10 casos de NIC3 y carcinoma inflitrante; estos resultados sugieren en su forma episomal, el VPH induce trisomías y tetrasomías y cuando está integrado aumenta el número de aneusomías y de polisomías (30). Se ha estudiado el gen PICK3CA y se describe en un 43% de las copias de 3q en el cáncer cervical (31, 32,33 ). Este gen se detecta en la amplificación de 3q26.3, y su expresión es la proteína p110α asociada con incremento de la actividad de kinasa y alteraciones del crecimiento celular con disminución de la apoptosis (33,34). El gen de la triada frágil-histidina FHIT localizado en el cromosoma 3 (3p14.2) también ha sido implicado en la carcinogénesis cervical (35). La expresión aberrante del gen FHIT ha sido descrita como un factor de mal pronóstico en el cáncer cervical asociado a la infección con VPH (36). La presencia temprana en NIC del gen FIHT ha planteado utilizar su detección en muestras de grandes poblaciones como una guía para señalar alteraciones tempranas en la carcinogénesis cervical (37). En un trabajo más reciente realizado en México se señala a través de micromatrices de ADN y con HiGC al gen FHIT de 3p14.2, e igualmente se describen alteraciones por ganancias o pérdidas en el cromosoma 4 (4q24), también del gen KIT en 4q11-q12, los cromosomas 1p32.3, 8p23.3, 9qtel, 13q14, y Xq12; curiosamente alteraciones del brazo largo del cromosoma 18 (18qtel) (38). A pesar de ser poco mencionado en relación con el cáncer cervical, seleccionamos este cromosoma para ser examinado con HiGC en nuestro trabajo y con el pudimos demostrar marcadas poliploidías en un caso de NIC2 ( Ver Figs. 6 y 7). A través de una línea celular de queratinocitos del cérvix que adquirieron VPH16 por infección natural, se ha logrado demostrar como la incorporación del genoma viral se produce en las células tras una larga fase episomal y cuando esto ocurre, es a través de una deleción en el cromosoma 3 como consecuencia de la represión post trascripccional ejercida por E2 la cual conlleva a la desregulación del promotor homólogo con incremento de los niveles de proteínas en E7(39). Se sabe que las proteínas E6 y E7 del VPH-16 son capaces de provocar amplificación del centrosoma con desfase de los cromosomas durante la mitosis. Estás anormalidades provocan inestabilidad cromosómica. La poliploidía se produce como consecuencia de una falla en la conformación del huso, secundaria al efecto de E6 y E7 sobre las funciones de p53 y de las proteínas de la familia del gen del retinoblastoma, con desregulación de genes como Plk1, Aurora-A, cdk1 y Nek2 que controlan la transición de la fase G2-M en la progresión hacia la mitosis (40). Inestabilidad cromosómica con tetraploidías es un evento temprano en la carcinogénesis cervical, y los queratinocitos alterados por la infección con VPH de alto riesgo tienden a hacer aneuploidías frecuentemente con pérdidas en el cromosoma 17 (41). En la piel vecina al carcinoma epidermoide de vulva se ha detectado inestabilidad cromosómica ( aneuploidías ) en sincronía con la neoplasia y en particular se han descrito anormalidades del cromosoma 17 que no se observan en áreas de piel normal; este tipo de estudios hechos con FISH han demostrado pérdidas en el cromosoma 17 en un 5% de los casos y siempre asociadas a la infección con VPH (42). Todas las evidencias de amplificación descritas en el cromosoma 3, con tetrasomías del mismo y con simultaneas trisomías del cromosoma 17 valieron para que estos fuesen seleccionados por nosotros para estudiarlos con HGC. La poliploidía como evento temprano durante la carcinogénesis cervical predispone a la aneuploidía, la cual conduce directamente hacia la malignización, conclusiones estas señaladas por Liu Y y col., quienes demostraron que E6 es capaz de inducir poliploidía por la vía de p53 pero también independientemente de ese mecanismo(43). Otros creen que pueden ser las aneuploidías las directamente inductoras de la inestabilidad genética y de los cambios neoplásicos (44); cualquiera de las dos hipótesis puede ser válida y de aquí que destaquemos la importancia de la poliploidía en la evolución hacia la carcinogénesis (45,46). G Marzano y col demostraron la importancia de las aneusomías de los cromosomas 3 y X en el desarrollo y la progresión de las lesiones de NIC hacia la malignidad (47). El análisis del cromosoma-1 ya mencionado previamente ( reveló una tendencia lineal positiva entre la frecuencia de aneusomía y el grado de lesión cervical y una asociación entre aneusomía y la infección de VPH de alto riesgo, resultados estos que han sugerido la utilidad del cuantificar el número de células cervicales con aneusomía del cromosoma-1 como un biomarcador de riesgo de cáncer (48). Nuestras observaciones utilizando sondas centroméricas para los cromosomas 3, 17 y 18 se muestran en varias imágenes expuestas en este trabajo. Un mayor número de alteraciones en las muestras de NIC2 y NIC3 con poliploidía en el cromosoma 3 coinciden con los cambios cromosómicos tempranos previamente descritos por otros investigadores, y son similares a nuestros hallazgos ( Ver Fig 3 ). En 1996 NG Mullokanddov y col., describió pérdida de la heterocigosis en el 15% de los cromosomas 17, en un 35% en el cromosoma 18 y en un 39% del cromosoma 3 en el carcinoma del cuello uterino asociado a la infección con el VPH (49). La observación ocasional de poliploidías y más frecuentemente de pérdidas tardías en el cromosoma 17 coinciden con evidencias ya señaladas y también con los cambios observados en los casos de cáncer. ( Ver Fig 4 ). Los estudios con micromatrices de ADN han servido para precisar la localización de diferentes genes que sufren alteraciones en el cáncer del cuello uterino (33,34,38,41). La utilización de la técnica de HGC puede ser útil para detectar los cromosomas que poseen genes que muestren ser importantes en el desarrollo de los cambios asociados al proceso de malignización en el carcinoma del cuello uterino. Referencias
Copyright 2010 - Academia Biomedica Digital The following images related to this document are available:Photo images[va10010f7.jpg] [va10010f2.jpg] [va10010f5.jpg] [va10010f4.jpg] [va10010f3.jpg] [va10010f6.jpg] [va10010f1.jpg] |
| |||||||||
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}