 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Academia Biomedica Digital, Vol. 0, No. 43, 2010 Bioquímica Actividad Antihiperglicemiante de Bauhinia megalandra.(Revisión) Antihiperglycaemic activity of Bauhinia megalandra Freddy González-Mujica1 , Norma Motta2 1Médico-cirujano, Ph.D. en Bioquímica,
Sección de Bioquímica Médica, Instituto de Medicina Experimental, Facultad
de
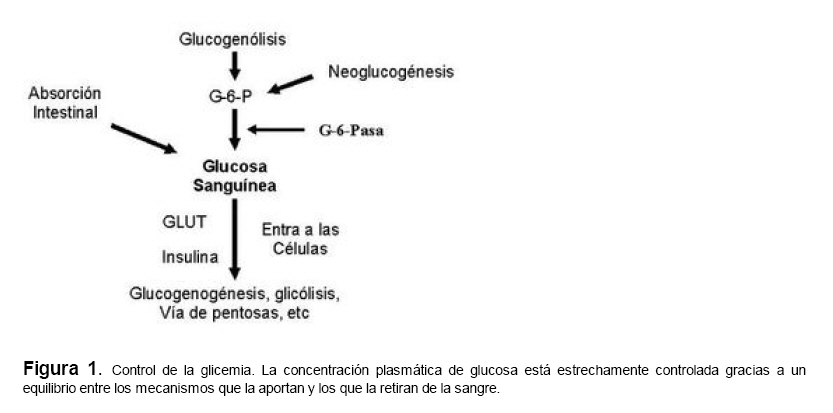
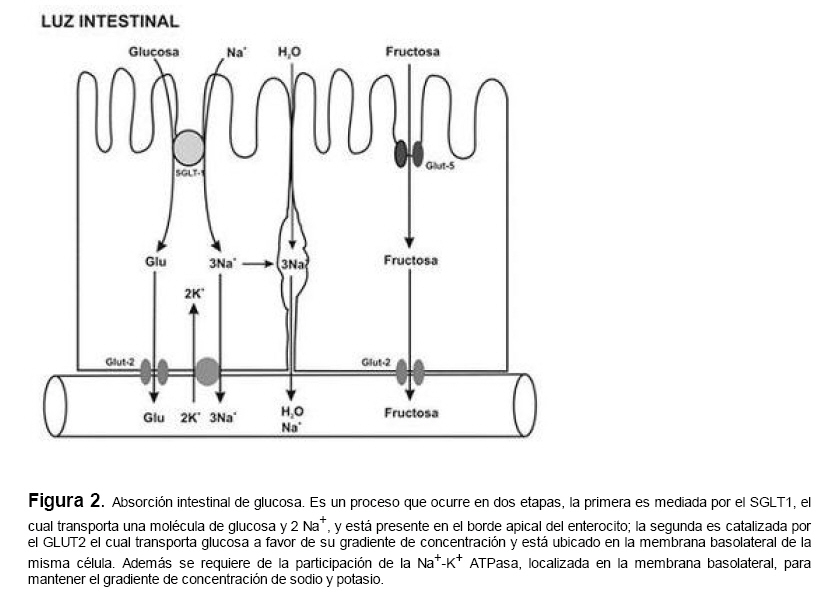
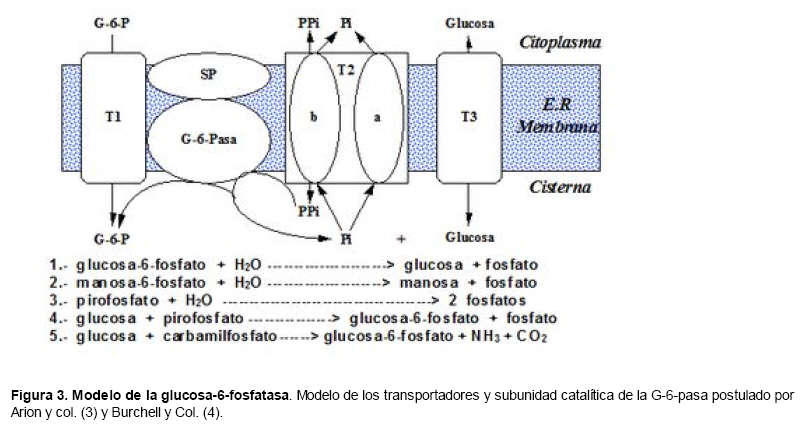
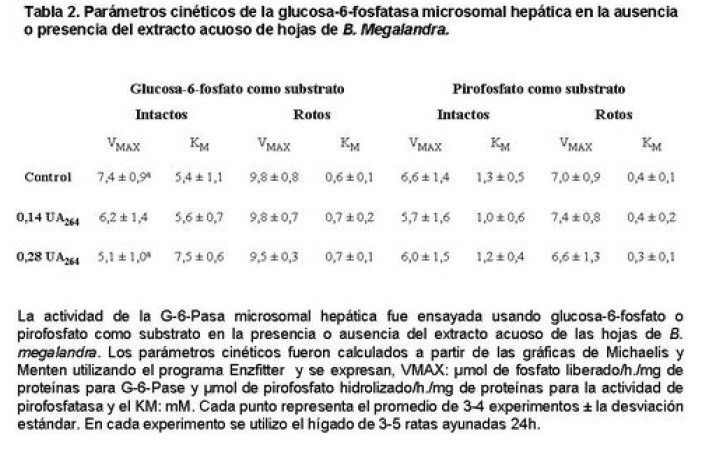
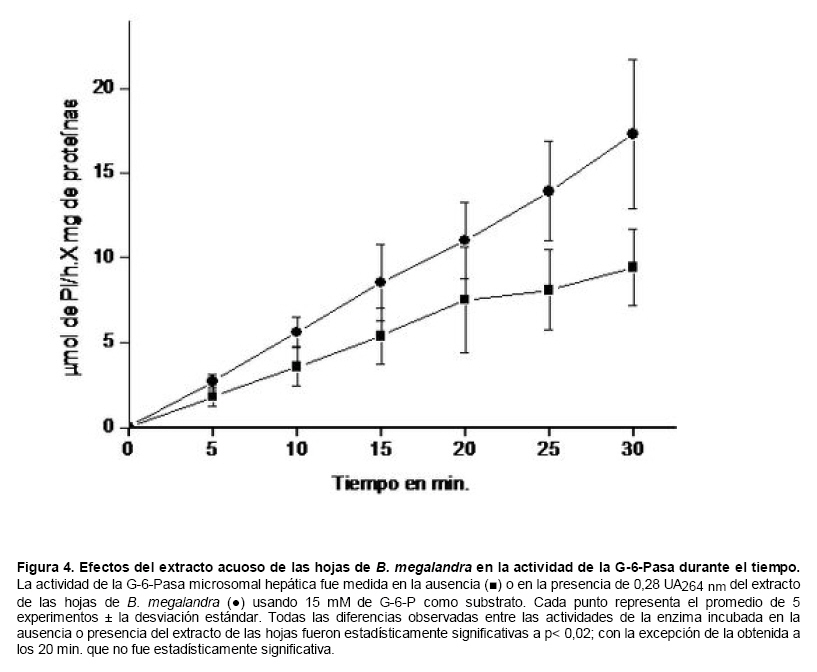
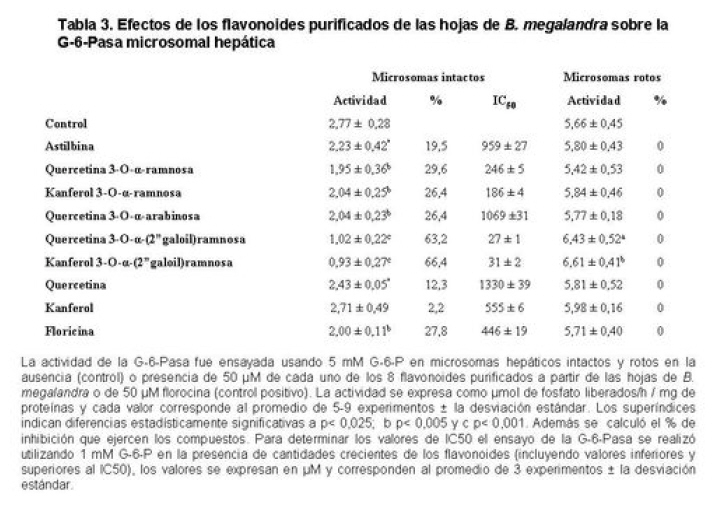
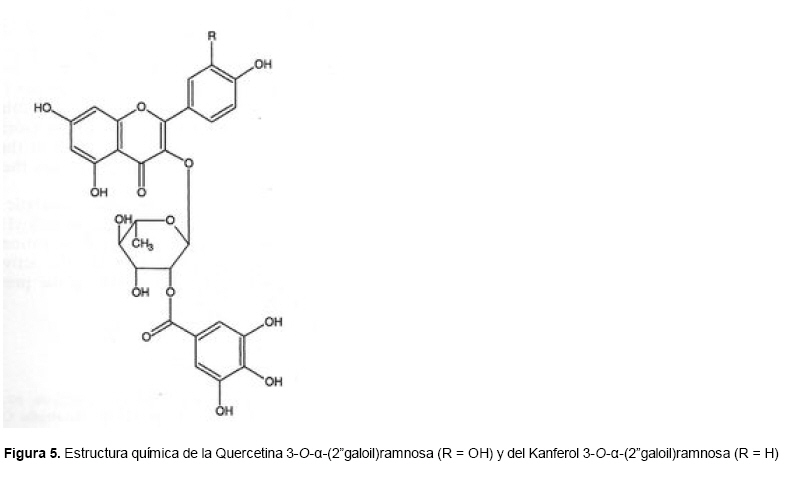
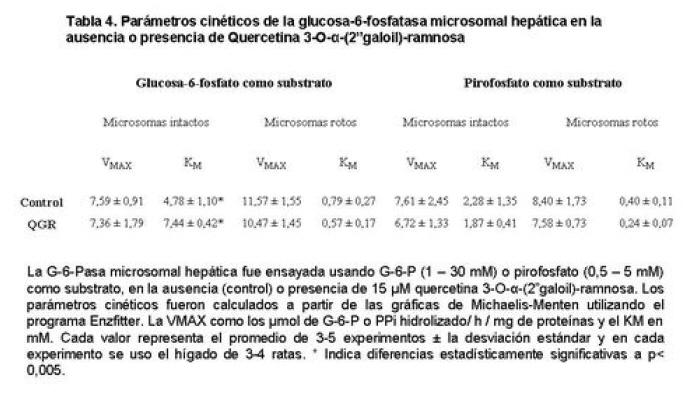
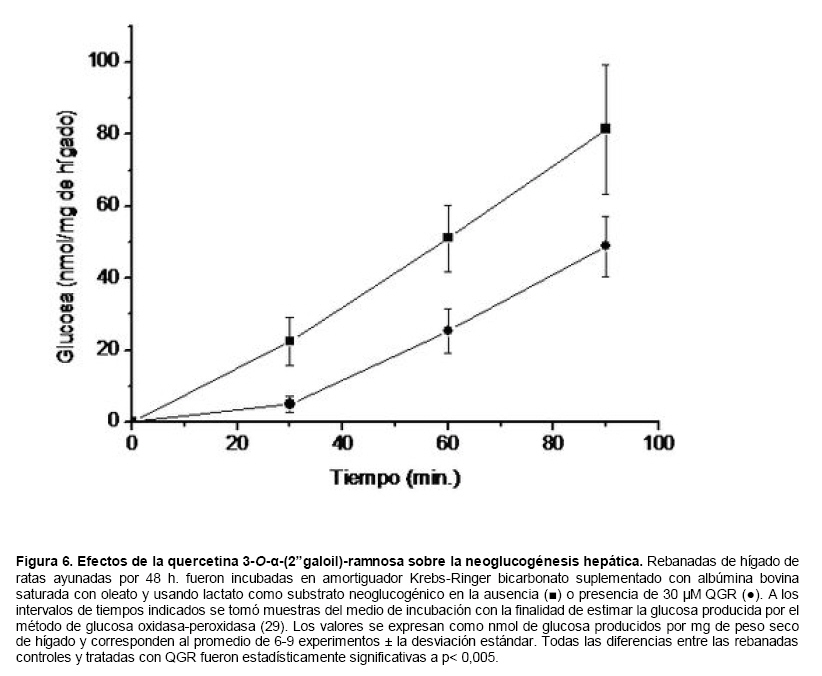
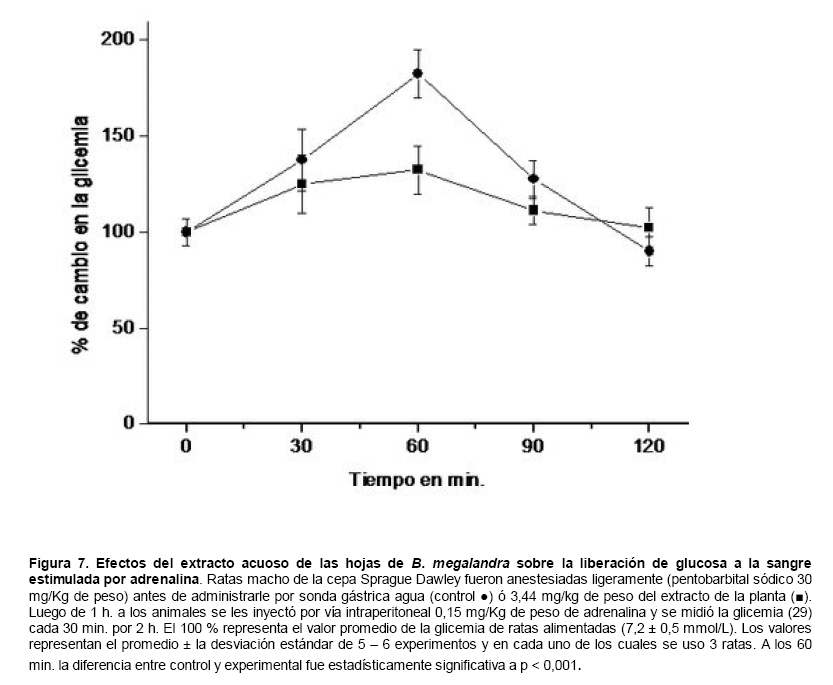
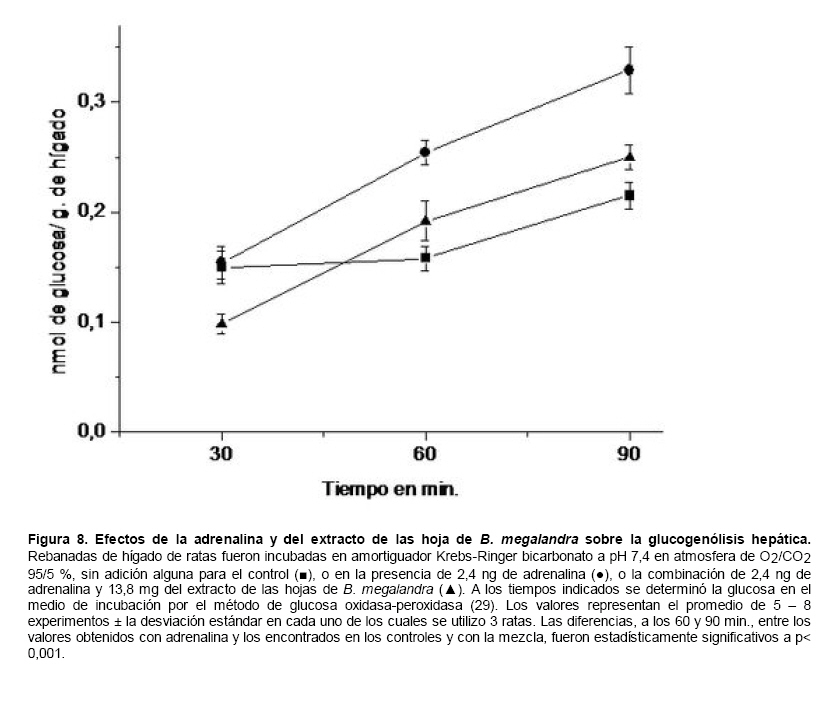
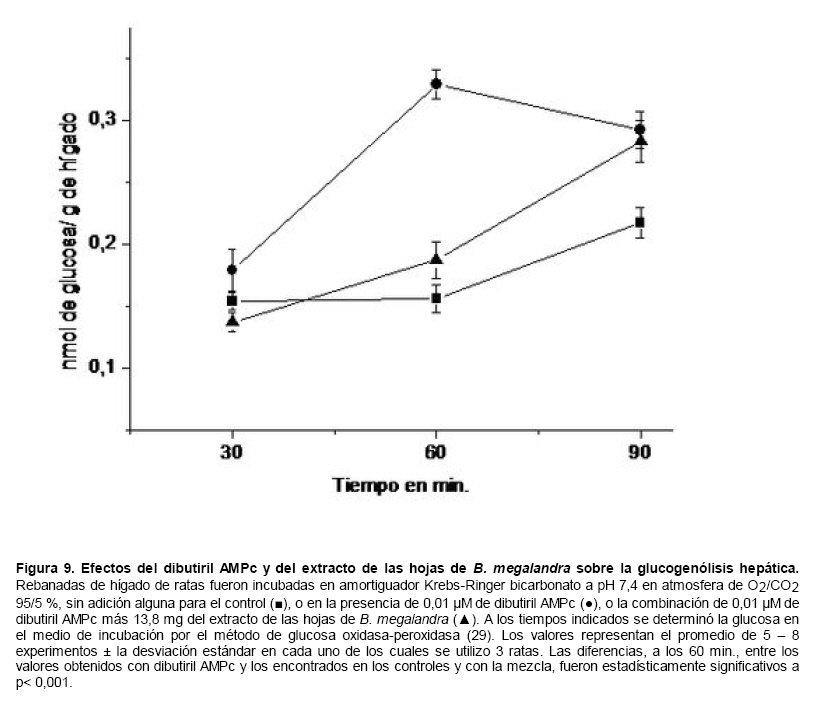
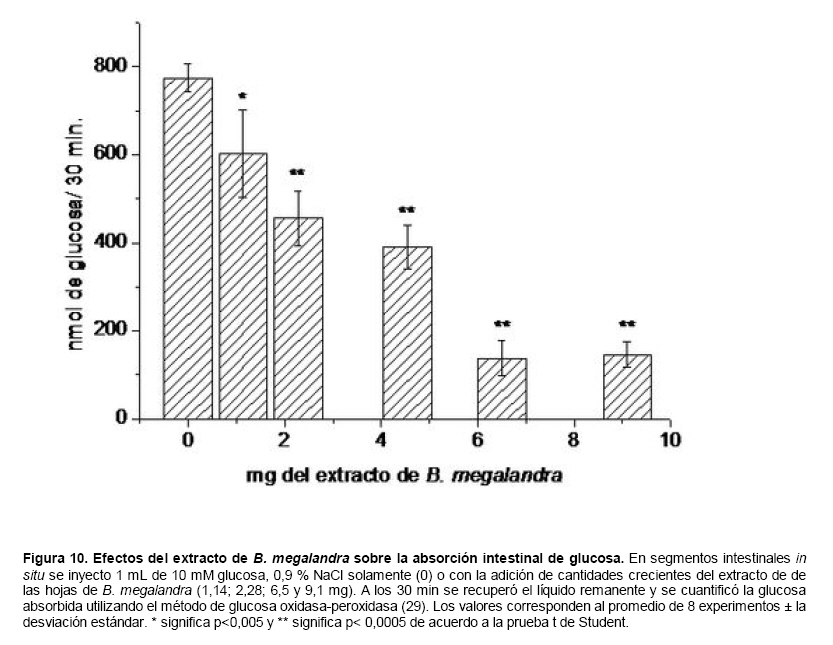
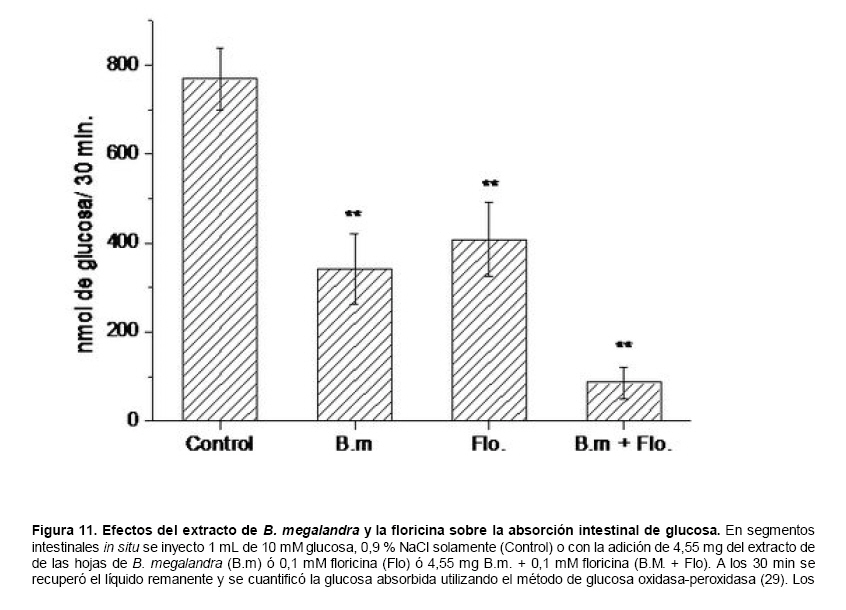
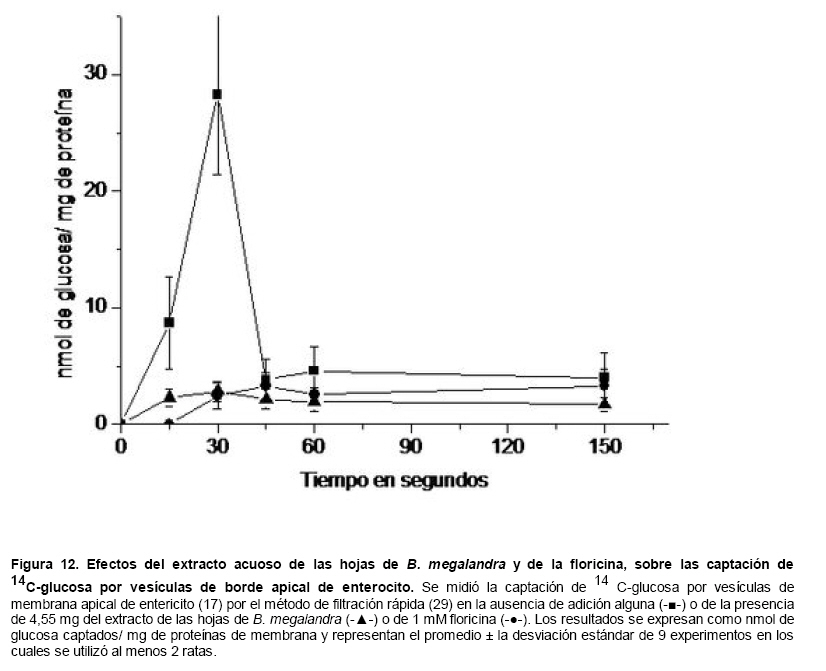
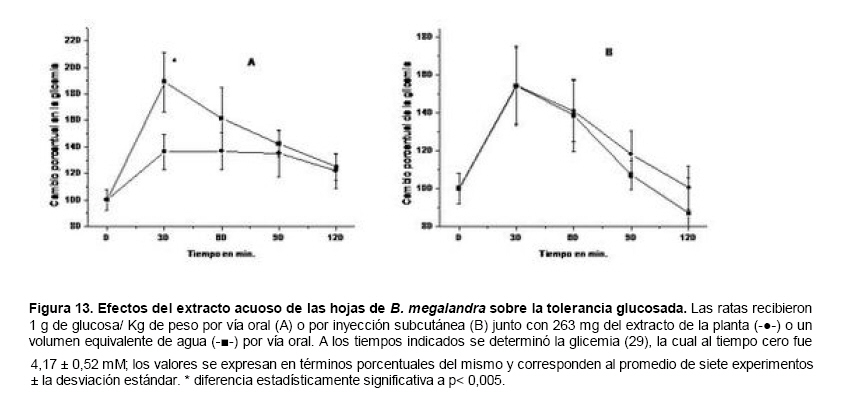
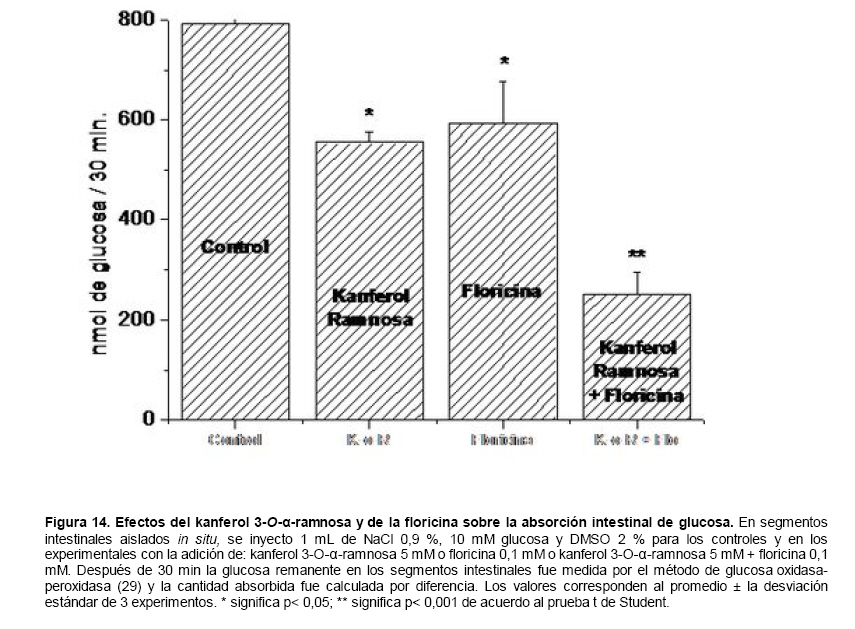
Medicina, Universidad Central de Venezuela, freddygonzalezmujica@gmail.com Fecha de recepción: 16/06/2010 Code Number: va10019 La infusión de las hojas de Bauhinia megalandra es usada por la medicina popular venezolana en el tratamiento empírico de la diabetes. En el presente trabajo se pasa revista a los resultados obtenidos por nosotros sobre las propiedades antihiperglicemiantes de esa planta. El extracto acuoso de las hojas de B. megalandra fue capaz de inhibir la capacidad neoglucogénica y glucogenolítica, estimulada por adrenalina o dibutiril AMPc, de rebanadas de hígado, de reducir la hiperglicemia promovida por la adrenalina en el animal intacto e inhibió la absorción intestinal de glucosa en una manera dosis dependiente afectando al transportador SGLT1. A partir del extracto de metanol de las hojas de B. megalandra purificamos e identificamos 8 flavonoides uno de los cuales, kanferol 3-O-α-(2"galoil)-ramnosa, en el mejor de nuestro conocimiento es la primera vez que se describe su aislamiento como un producto natural; todos ejercieron efecto inhibitorio sobre la enzima glucosa-6-fosfatasa, siendo más activos los compuestos galoilados con IC50 de aproximadamente 30 µM. La quercetina 3-O-α-(2"galoil)-ramnosa se comportó como un inhibidor competitivo del transportador T1 de la glucosa-6-fosfatasa y redujo drásticamente la neoglucogénesis hepática. El kanferol 3-O-α-ramnosa se comportó como un inhibidor competitivo de la absorción intestinal de glucosa. En ratas que ingirieron el extracto de la planta se produjo una discreta disminución de la glicemia y glucosuria renal y en la orina de las mismas identificamos un benceno ortodisustituido, posiblemente producto del metabolismo de alguno de los flavonoides. Se discuten las posibles implicaciones de estos efectos. Palabras Claves: Bauhinia megalandra, antihiperglicemiante, flavonoides, glucosa-6-fosfatasa, absorción intestinal de glucosa, SGLT 1, glucogenólisis, neoglucogénesis, diabetes. Abstract The infusion of Bauhinia megalandra leaves is used in the Venezuela folk medicine for the treatment of diabetic patients. In the present paper we review our results in the antihiperglycaemic activity of this plant. The aqueous extract of B. megalandra leaves inhibit the neoglucogenic and glycogenolysis, stimulated by epinephrine or dibutyril AMPc, activity of liver slices, reduced the hiperglycamia promoted by epinephrine in the intact animal and inhibited the glucose intestinal absorption in a dose dependant way by affecting the SGLT 1 transporter. From the methanol extract of the B. megalandra leaves, we purified and characterized 8 flavonoids one of which, kaempferol 3-O-α-(2"galloyl)-rhamnoside, in the best of our knowledge this is the first report of it isolation from nature; all of them were able to inhibit the enzyme glucose-6-phosphatase, being the most active those with the galloyl group with IC50 of approximately 30 µM. Quercetin 3-O-α-(2"galloyl)-rhamnoside behaved as a competitive inhibitor of the glucose-6-phosphatase T1 transporter and drastically reduced the liver gluconeogenic activity. Kaempferol 3-O-α-(2"galloyl)-rhamnoside was a competitive inhibitor of the glucose intestinal absorption. The rats that drank the plant extract showed a slight decrease in the glycaeamia and renal glycosuria and in the urine we identified a benzene ortodisubstituted, probably product of the metabolism of one of the flavonoides. The implications of these findings are discussed. Key Word: Bauhinia megalandra, antihiperglycaemic, flavonoid, glucose.6.phosphatase, glucose intestinal absorption, SGLT 1, glycogenolysis, gluconeogenesis, diabetes. Introducción La glicemia es uno de los parámetros homeostáticos estrechamente controlados gracias a un equilibrio entre los mecanismos que aportan glucosa a la sangre y los que la retiran de ella (Fig. 1). La absorción intestinal es uno de los mecanismos que aportan glucosa a la sangre y ocurre en dos etapas (ver Fig.2): la primera es mediada por el transportador Na+-glucosa (SGLT1) ubicado en la membrana apical del enterocito y el cual transloca 2 iones Na+, a favor de su gradiente de concentración, por molécula de glucosa, a favor o en contra de su gradiente de concentración dependiendo del período de la digestión y de la cantidad de carbohidratos ingeridos, desde el lumen del intestino al interior de la célula. La segunda está a cargo del transportador de glucosa 2 (GLUT 2) ubicado en la membrana basolateral del enterocito y transporta glucosa por difusión facilitada desde el enterocito al espacio intersticial (1). Durante el ayuno corto, la degradación del glucógeno (glucogenólisis) hepático aporta glucosa a la sangre. Si el ayuno se prolonga la síntesis de nuevo de glucosa (neoglucogénesis), en particular en hígado y riñón, suplen glucosa a la sangre. La enzima glucosa-6-fosfatasa (G-6-Pasa EC 3.1.3.9), por catalizar la reacción final de la glucogenólisis y la neoglucogénesis juega un papel muy importante en el mantenimiento de la glicemia (2). La G-6-Pasa se encuentra en el retículo endoplasmático (RE) en un 80-90 % y en la envoltura nuclear (10-20 %) de células del hígado, riñón, mucosa intestinal, y células β de los islotes pancreáticos, principalmente. Con base en estudios cinéticos, genéticos y moleculares, Arion y col. (3) y Burchell y col. (4) propusieron que la G-6-Pasa (Fig. 3) está constituida por: una subunidad catalítica, proteína transmembrana cuyo centro activo mira a la cisterna del RE y que muestra poca especificidad, pudiendo hidrolizar varios ésteres fosfóricos así como pirofosfato. Un transportador T1 altamente específico para el substrato glucosa-6-fosfato (G-6-P). T2, un transportador de Pi, PPi y carbamilfosfato el cual se ha sugerido que está constituido por 2 subunidades, y un transportador para glucosa, T3. Se piensa que también existe una proteína estabilizadora de la subunidad catalítica (SP). La G-6-Pasa además de la actividad hidrolítica, en condiciones adecuadas de alta concentración de glucosa y de un donador de fosfato: PPi o carbamilfosfato es capaz de sintetizar G-6-P. La entrada de glucosa a los tejidos, mediada por los GLUTs, (5) disminuye la glicemia, siendo de especial interés la participación del GLUT 4 en el músculo cardíaco y esquelético así como en el tejido adiposo, en estos es translocado a la membrana plasmática desde vesículas intracelulares por acción de la insulina incrementándose la entrada de glucosa a dichos tejidos. Las drogas que disminuyen los mecanismos que aportan glucosa a la sangre se consideran antihiperglicemiantes y las que incrementen la entrada de glucosa a los tejidos, directa o indirectamente, son hipoglicemiantes. McCormack y col. (6) sugieren que la G-6-Pasa pudiera ser un blanco potencial para drogas antihiperglicemiantes. La fracción microsomal, la cual está enriquecida con RE, que se obtiene por centrifugación diferencial está formada por vesículas intactas y rotas, la proporción de cada una de ellas se puede medir utilizando manosa-6-fosfato como substrato de la G-6-Pasa (7), molécula que no es translocada por T1; en nuestras preparaciones como mínimo tenemos 95 % de vesículas intactas. Las preparaciones que se obtienen se denominan microsomas no tratados; los microsomas se pueden romper por diferentes métodos tales como el uso de detergentes, cavitación en nitrógeno, prensa francesa y más recientemente con el uso de histonas. El termino microsomas intactos es teórico y se obtiene substrayendo a la actividad de la G-6-Pasa presente en los microsomas no tratados la actividad de la enzima exhibida por las vesículas rotas presentes. Desde tiempos inmemoriales, el hombre ha utilizado plantas en el tratamiento empírico de distintas enfermedades y en particular de la diabetes En Venezuela, como en muchos países tropicales, las hojas del género Bauhinia, popularmente conocida como "casco de vaca" han sido utilizadas por la medicina tradicional en el tratamiento empírico de la diabetes; pertenece a la familia Fabaceae, que comprende las leguminosas. Estas plantas (8) son árboles que pueden alcanzar los 10 m. de altura, poseen hojas bien desarrolladas y grandes, siempre verdes, y divididas desde el ápice hasta más o menos un tercio de su longitud y asemejan la huella de un bovino, son vegetales hermafrodita. La especie B. megalandra se caracteriza por poseer flores blancas en racimos o solitarias con pétalos delgados y largos, el fruto se encuentra como legumbre compacta de color marrón. Es de crecimiento rápido, de vida larga y sistema radical profundo. Efectos de B. megalandra sobre la neoglucogénesis y la G-6-Pasa La B. megalandra utilizada en estos experimentos fue identificada por El Dr. Stefen Tillett del Herbario Ovalles de la Facultad de Farmacia de la Universidad Central de Venezuela. Inicialmente medimos los efectos del extracto acuoso de las hojas de B. megalandra (9) sobre la neoglucogénesis de rebanadas de hígado de rata, incubadas en Krebs Ringer bicarbonato suplementado con albúmina bovina saturada con oleato y utilizando lactato o fructosa como substrato neoglucogénico. Como se puede observar en la Tabla 1, la actividad neoglucogénica de las rebanadas controles es lineal hasta los 90 min. e igual con los dos substratos. En la presencia del extracto de la planta ocurre una drástica disminución de la neoglucogénesis, siendo más marcada a partir de lactato que de fructosa; estos resultados sugieren que en el extracto de B. megalandra están presente compuestos que bloquean dicho proceso en un punto más allá de la entrada de ambos substratos. Como se muestra en la Tabla 2, cuando se usó G-6-P como substrato de la G-6-Pasa de microsomas intactos, el extracto foliar disminuyó de manera estadísticamente significativa la VMAX con una moderada elevación del KM sin afectar la actividad de la enzima de microsomas rotos (9). El extracto de la planta, no tuvo efecto alguno cuando el substrato de la enzima fue PPi, tanto en microsomas intactos como rotos; lo cual sugiere que el efecto del extracto de B. megalandra se ejerce sobre uno de los transportadores del sistema de la G-6-Pasa probablemente sobre T1 (9). Posiblemente la inhibición de la neoglucogénesis hepática sea el resultado de la inhibición de la G-6-Pasa por los compuestos presentes en el extracto de la planta. Al medir el efecto que durante el tiempo, ejerce el extracto de la planta sobre la actividad de la enzima en microsomas no tratados (Figura 4) se observó que: en los controles la actividad de la G-6-Pasa fue lineal durante 30 min., en la presencia del extracto de B. megalandra la actividad de la enzima también fue lineal hasta los 20 min. pero por debajo del control, después de este punto ocurre una inflexión con una línea entre 20 y 30 min. con una pendiente menor, sugiriendo que por efectos de la presencia del extracto foliar probablemente se inhibe otro de los transportadores del sistema de la G-6-Pasa acumulándose los productos de la reacción enzimática los cuales inhibirían la subunidad catalítica (9). En vista de que los parámetros cinéticos de la G-6-Pasa usando PPi como substrato no fueron afectados por el extracto de la planta, posiblemente el transportador inhibido sea T3. Purificación de flavonoides de B. megalandra y cuantificación de sus efectos. A partir del extracto metanólico de las hojas frescas de B. megalandra logramos purificar 8 flavonoides (10), la mayoría de ellos como quercetina y kanferol o sus derivados y se midió el efecto que estos compuestos ejercen sobre la actividad de la G-6-Pasa. Las agliconas mostraron la menor actividad inhibitoria, los compuestos glicosilados con ramnosa presentan una actividad intermedia y cuando la ramnosa se encuentra esterificada al ácido gálico, la inhibición supera el 60 %. Vale la pena destacar que el efecto inhibitorio solo se observa en los microsomas intactos (Tabla 3) lo cual sugiere fuertemente que estos flavonoides afectan uno de los transportadores, probablemente T1, del sistema de la G-6-Pasa. Hasta donde sabemos, esta es la primera vez que se describe la presencia de kanferol 3-O-α-(2"galoil)-ramnosa como producto natural cuya estructura, conjuntamente con la de quercetina 3-O-α-(2"galoil)-ramnosa, se muestra en la Figura 5. Se encontró una correlación entre el % de inhibición de la actividad de la G-6-Pasa de microsomas intactos y el valor del IC50 ejercidos por los compuestos estudiados; las agliconas: kanferol y quercetina, la quercetina glicosilada con arabinosa (una pentosa) y la astilbina presentan valores de IC50 elevados y porcentajes de inhibición bajos; la quercetina y el kanferol glicosilados con ramnosa (una hexosa) poseen valores intermedios de IC50 y porcentajes de inhibición intermedios y los flavonoides unidos a ramnosa y ácido gálico exhibieron IC50 del orden de 30 μM y mostraron inhibición de la actividad de la enzima cercana al 60 %. Se utilizó como control positivo floricina, un conocido inhibidor de T1 de la enzima (11), el cual mostró un efecto inhibitorio similar al ejercido por el kanferol y la quercetina unidos a ramnosa pero un valor de IC50 (466 μM) el cual es cercano al doble del mostrado por los primeros; siendo la primera vez que se reporta el IC50 de la floricina para la G-6-Pasa de microsomas intactos. Como se muestra en la Tabla 4, quercetina 3-O-α-(2"galoil)-ramnosa (QGR) incrementó considerablemente el KM para G-6-P sin modificar la VMAX de la G-6-Pasa de microsomas intactos, tampoco afectó los parámetros cinéticos de la enzima de microsomas rotos. Cuando se usó PPi como substrato (Tabla 4), los parámetros cinéticos de la G-6-Pasa tanto de microsomas intactos como rotos no fueron afectados por la presencia de QGR. Estos resultados sugieren fuertemente que QGR es un inhibidor competitivo del transportador T1 del sistema de la G-6-Pasa (12). La capacidad neoglucogénica de rebanadas de hígado de ratas ayunadas por 48 horas e incubadas en la ausencia (control) o presencia de 30 μM QGR se muestra en la Fig. 6. La producción de glucosa por las rebanadas controles fue prácticamente lineal y muy similar a lo reportado antes (9). QGR inhibió drásticamente la capacidad neoglucogénica de rebanadas de hígado de ratas a todos los tiempos estudiados: a 30 min. la inhibición fue de aproximadamente 85 %; cercana al 50 % a los 60 min. y de un 41 % a los 90 min. Aun cuando la inhibición de la capacidad neoglucogénica de las rebanadas de hígado por la presencia del flavonol en el medio de incubación parece disminuir con el tiempo de incubación, sin embargo una observación detallada de la Fig. 6 muestra que la pendiente de la línea que representa el efecto del flavonol, luego de los 20 min. iniciales, es 0.74 lo cual es considerablemente menor que 0.97 de la condición control. Efectos de B. megalandra sobre la glucogenólisis hepática Como podemos observar en la Figura 7, al inyectar adrenalina intraperitonealmente (13), se incrementó la glicemia con un máximo de 82 % a los 60 min. La administración oral del extracto de las hojas de B. megalandra 60 min. antes de la inyección de la catecolamina, redujo en aproximadamente un 50 % la elevación de la glicemia producida por el efecto de la adrenalina (14). Estos resultados son una evidencia indirecta de la absorción intestinal de alguno(os) compuesto(s) presentes en el extracto vegetal los cuales al alcanzar el hígado disminuyen la glucogenólisis por lo cual no se eleva la glicemia por efecto de la adrenalina. La presencia de adrenalina en el medio de incubación de rebanadas de hígado de ratas alimentadas, incrementa la liberación de glucosa en un 60 % y un 53 % a los 60 y 90 min de estudio respectivamente (Figura 8). Por el contrario la liberación de glucosa por las rebanadas de hígado en la presencia simultánea de adrenalina y del extracto vegetal, fue paralela y siempre significativamente menor a la producción de glucosa en presencia de adrenalina sola, mostrando 38; 25 y 24 % de inhibición a los 30; 60 y 90 min. respectivamente. Así mismo a los 30 min. de incubación, la presencia simultanea de adrenalina y el extracto de la planta reduce en un 38 % la producción de glucosa en relación al control (14). En la Figura 9 se muestra el efecto del dibutiril-AMPc sobre la producción de glucosa por rebanadas de hígado de rata (14); como puede observarse la presencia del nucleótido condicionó un incremento en la liberación de glucosa por las rebanadas de hígado a todos los tiempos estudiados, con un máximo a los 60 min. La presencia simultanea del extracto acuoso de B. megalandra y dibutiril AMPc provocó una disminución del efecto del nucleótido la cual fue más marcada a los 60 min cuando se observó una reducción de un 43 %. Los resultados obtenidos en estas condiciones ex vivo (Figuras 8 y 9) evidencian que los compuestos presentes en las hojas de B. megalandra afectan un evento posterior a la producción del AMPc, probablemente la actividad de la G-6-Pasa. La única reacción que tienen en común las vías glucogenólisis y neoglucogénesis es la catalizada por la enzima G-6Pasa. En consecuencia el conjunto de resultados, presentados hasta ahora, sugieren fuertemente que los flavonoides presentes en las hojas de B. megalandra inhiben la G-6-Pasa lo cual se traduce en una reducción de la capacidad de aportar glucosa a la sangre por la glucogenólisis y la neoglucogénesis hepática. Dichos flavonoides pudieran ser útiles en el tratamiento de la diabetes no insulino dependiente ya que muestran un efecto antihiperglicemiante. Efectos de B. megalandra sobre la absorción intestinal de glucosa. Usando como modelo experimental segmentos intestinales aislados in situ, demostramos (15) que la absorción intestinal de glucosa es inhibida por el extracto acuoso de las hojas de la planta en una manera dependiente de la dosis (ver Figura 10).Al estar presente simultáneamente el extracto acuoso de B. megalandra y floricina, un conocido inhibidor de SGLT 1 (16), se observó un efecto inhibitorio aditivo lo cual sugiere que en la planta están presente compuestos capaces de inhibir dicho transportador (ver Figura 11). Al incubar vesículas de membrana apical de enterocitos en un medio externo con 14C-glucosa y Na+ comprobamos el típico comportamiento con un pico máximo de captación de glucosa a los 30 seg. para luego decaer (17). En presencia del extracto de la planta o de floricina se elimina el pico de captación, resultado que claramente indica que en el extracto de la planta existen compuestos capaces de inhibir el transportador SGLT 1, confirmando lo sugerido de acuerdo al experimento anterior (15). La inhibición de la absorción intestinal de glucosa por el extracto de ésta planta, afectando al SGLT 1, es similar al reportado para los polifenoles del te verde (18) y las isoflavonas de soya (19) pero diferente al ejercido por la acarbosa, la cual inhibe la digestión de los carbohidratos y no la absorción de los mismos (20). La administración simultánea, por vía oral, de glucosa y del extracto foliar condiciona una curva de tolerancia glucosada casi plana (Figura 13). Por el contrario, cuando la glucosa se administró subcutáneamente y el extracto vegetal por vía oral no se observó efecto alguno sobre la curva de tolerancia glucosada. Estos resultados muestran claramente que el extracto de la planta afecta la absorción intestinal de glucosa y no otros factores relacionados con la glicemia tales como la producción y/o liberación de insulina ni el consumo de glucosa por los tejidos (15). Estudiamos el efecto que los flavonoides: kanferol 3-O-α-ramnosa, quercetina 3-O-α-ramnosa y kanferol 3-O-α-(2" galoil)-ramnosa, purificados antes por nosotros (10), ejercen sobre la absorción intestinal de glucosa, encontramos que solo el kanferol 3-O-α-ramnosa disminuyó de una manera apreciable (28 %), la absorción intestinal de glucosa (Figura 14). En relación a la estructura química y la actividad biológica vale la pena destacar: a) la presencia del grupo galoilo unido a la ramnosa anula la capacidad inhibitoria del kanferol 3-O-α-ramnosa sobre la absorción intestinal de glucosa; b) la ramnosa no es determinante para condicionar la actividad biológica, ya que esta presente en ambos flavonoides y solo uno de ellos inhibe la absorción intestinal de glucosa y c) la presencia de un H (kanferol) en lugar de un OH (quercetina) en la posición 3' del anillo B del flavonoide, es determinante para condicionar la inhibición de la absorción intestinal de glucosa. (21) Como se muestra en la Figura 14, la presencia conjunta de kanferol 3-O-α-ramnosa y floricina inhiben la absorción intestinal de glucosa (68 %) en más del doble del efecto de cualquiera de los dos compuestos solos (kanferol 3-O-α-ramnosa 28 % y floricina 25 %). Estos resultados son una clara evidencia del efecto sinérgico del kanferol 3-O-α-ramnosa y la floricina, un inhibidor conocido del SGLT 1 (16). En la Tabla 5 se muestran los valores cinéticos: KM y VMAX, de la absorción intestinal de glucosa en ausencia, control, y presencia de kanferol 3-O-α-ramnosa. Los valores controles son diferentes a los reportados por Ader y Col. (22) y por Li y Col. (23), al respecto es importante destacar que en ambos trabajos los parámetros cinéticos publicados corresponden a los del SGLT1 y en nuestro caso conciernen a todo el proceso de la absorción intestinal de glucosa. El kanferol 3-O-α-ramnosa se comporta como un inhibidor competitivo de la absorción intestinal de glucosa ya que incrementa el KM con una disminución de la VMAX no significativa estadísticamente (21). De los resultados anteriores podemos concluir que el kanferol 3-O-α-ramnosa es un inhibidor competitivo del SGLT 1. Utilizando extracciones con solventes de polaridad creciente y cromatografía en silica gel, purificamos parcialmente apigenina 8-glucósido, el cual inhibió la absorción intestinal de glucosa en una manera sinérgica con la floricina, lo cual sugiere que afecta al SGLT 1 (24) El uso de los flavonoides kanferol 3-O-α-ramnosa y apigenina 8-glúcosido pudiera ser útil en el tratamiento de la diabetes tipo II ya que disminuiría el aporte de glucosa a la sangre al reducir la absorción intestinal de misma, comportándose como antihiperglicemiantes. Absorción intestinal de flavonoides de B. megalandra y metabolismo de los mismos. Para que los flavonoides, presentes en el extracto de la planta, ejerzan su actividad biológica deben ser absorbidos a nivel intestinal, alcanzar la sangre y luego llegar a los tejidos blancos de dichas moléculas. Durante su permanencia en los organismos vivos, los flavonoides pueden ser modificados químicamente por la maquinaria enzimática de las células con las cuales entren en contacto. Hasta el momento hemos presentado evidencias indirectas, de la absorción intestinal de flavonoides como es el caso destacado en la Figura 7, experimento en el cual la administración oral del extracto de las hojas de B. megalandra antes de inyectar adrenalina por vía intraperitoneal reduce drásticamente la hiperglicemia condicionada por la catecolamina (14). Intentando obtener evidencias directas de la absorción intestinal de los flavonoides presentes en el extracto acuoso de las hojas de B. megalandra, analizamos la orina de ratas controles y de otras a las cuales se les administró durante una semana el extracto de la planta en lugar de agua. Usando fraccionamiento con varios solventes y cromatografías en silica gel, encontramos (25) en la orina de los animales que recibieron el extracto acuoso de las hojas de B. megalandra, la presencia de una mancha de color azul-grisáceo en las cromatografias de capa fina que no estaba presente ni en la orina de los controles ni en el extracto vegetal original, indicando la presencia de un metabolito de alguno de los compuestos, probablemente un flavonoide, presentes en las hojas de la planta. La estructura de ese compuesto se estableció, utilizando resonancia magnética nuclear de 1H y 13C, como un benceno orto disustituido; sin embargo en el presente no tenemos información sobre el tipo de sustituyentes y en consecuencia tampoco sobre la posible vía metabólica involucrada (25) en la transformación del compuesto original. Este benceno orto disustituido es diferente a los ácidos fenólicos, que se han reportado (26) son eliminados por la orina de animales que ingieren flavonoides. En los animales así tratados encontramos una discreta disminución de la glicemia del 9 % y un incremento importante de la eliminación urinaria de glucosa, de aproximadamente unas 4 veces. El efecto hipoglicemiante está de acuerdo con lo reportado por otros investigadores para otras especies del genero bauhinia (27,28) y la glucosuria pudiera ser interpretada como una evidencia indirecta de la absorción intestinal de los flavonoides los cuales por vía sanguínea alcanzan el riñón donde inhibirían los SGLT 1 condicionando glucosuria renal. Referencias
Copyright 2010 - Academia Biomedica Digital |
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}