 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
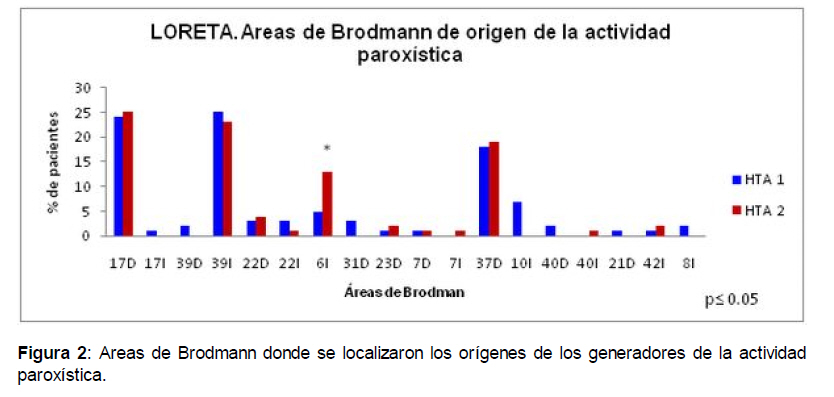
Academia Biomedica Digital, Vol. 0, No. 44, 2010 Neurociencia Localización de la actividad paroxística en pacientes con hipertensión arterial con el uso de la Tomografía Electromagnética de Baja Resolución (LORETA) Localization of paroxysmal activity using Low Resolution Electromagnetic Tomography (LORETA) in patients with High Blood Pressure María Esther de Quesada Martínez*, Mónica Reyes Moreno# Médico, Neurofisiología, Profesor Asociado, Cátedra de Fisiopatología, Escuela de Medicina “JM Vargas” Universidad Central de Venezuela. Fecha de recepción: 31/12/2000 Code Number: va10026 Por resultados anteriores en hipertensos sin complicaciones clínicas decidimos hacer un análisis mediante la Tomografía Electromagnética de Baja Resolución (LORETA) para determinar las áreas donde se originaban los paroxismos obtenidos en el Electroencefalogama digital (EEGd). 84 pacientes integraron dos grupos de acuerdo con su presión arterial sistólica (PAS) y diastólica (PAD): G1 (PAS=140-159; PAD=90-99), y G2 (PAS≥160; PAD≥100) vs controles sanos. En los EEGd de los pacientes se obtuvieron los paroxismos que se transformaron al sistema LORETA para su localización. Los paroxismos se localizaron principalmente en áreas 17 y 37 de Brodman del hemisferio derecho y en las áreas de 6, 39 y 10 de Brodman del hemisferio izquierdo. Estas localizaciones se corresponden a áreas limítrofes de la circulación cerebral más sensibles a la hipoperfusión cerebral que tiene lugar en la hipertensión arterial. Palabras Claves: Hipertensión arterial, problema inverso, hipoperfusión cerebral, actividad paroxística, tomografía eléctrica, LORETA, áreas limítrofes. Abstract Based in previous results on patients with High Blood Pressure (HBP), it was decided to use the Low Resolution Electromagnetic Tomography Analysis (LORETA) to map the cerebral areas where paroxysmal EEG activity was originated. 84 patients were divided into two groups: G1 (24 patients, systolic blood pressure (SP)=140-159; diastolic (DP) =90-99), G2 (60 patients, SP ≥160; DP ≥100) and 35 control subjects were studied with LORETA measures derived from their digital EEGs. The generators for the paroxysmal activity were mainly localized on the right Brodmannn´s 17 and 37 areas, and on the left Brodmannn´s 6, 39 and 10 areas. These localizations might be due to deficient cerebral blood flow, as these regions are very sensitive to hypoperfusion caused by arterial hypertension Key Word High Blood Pressure, inverse problem, cerebral hypoperfusion, paroxymal activity, electromagnetic tomography, LORETA. Introducción En estudios electroencefalográficos (EEG) realizados en pacientes con Hipertensión Arterial (HTA) (1) sin complicaciones clínicas neurológicas, se ha descrito la presencia de actividad paroxística focal a predominio de las regiones frontales (1). Otros autores han reportado, en las encefalopatías hipertensivas agudas la aparición de actividad paroxística y lenta en las regiones parieto-occipitales de ambos hemisferios (2). En pacientes con encefalopatía hipertensiva severa (T.A. 300/140) se ha reportado la aparición de actividad paroxística seguida por actividad lenta, lo cual mejora gradualmente después de estabilizar la condición clínica de los pacientes, aunque lo que disminuye es la presencia de los paroxismos y se mantiene la actividad lenta (3). Luego, la actividad paroxística es un elemento de frecuente aparición en el EEG de los pacientes con HTA en diferentes condiciones clínicas. Los paroxismos son grafoelementos transitorios, que no forman parte de la actividad de base del EEG, que se destacan de la misma por su inicio y terminación súbita, que alcanzan generalmente una amplitud destacada. La morfología y duración de los paroxismos es variable, la puntas (spikes) pueden durar entre 20 y 70 mseg y las ondas lentas angulares (sharp waves) pueden durar entre 70 y 200 mseg. Generalmente el componente principal de los paroxismos es negativo (4). Hay diferentes tipos de paroxismos en dependencia de su morfología, su forma de presentación en el EEG, su localización en un grupo de derivaciones o en forma generalizada; si se trata de paroxismos que forman una crisis eléctrica (críticos o ictales) o si no forman parte de una crisis (intercríticos o interictales); si se propagan a partir de su localización original o no. Las características de la actividad paroxística define la significación semiológica de los mismos en la interpretación de los EEGs (5). La localización de los paroxismos en un registro electroencefalográfico de superficie no se corresponde necesariamente con el área donde se está generando esta actividad, esto quiere decir que si en el EEG se observa la actividad paroxística en la derivación parietal derecha, puede estar originándose en otra región de la corteza o de estructuras subcorticales y lo que registramos es su proyección sobre esa área, lo cual depende de la geometría de los generadores y del diseño por circunvoluciones de la corteza cerebral. Esto se ha comprobado por registros intracorticales e intracerebrales y su relación con los correspondientes registros de superficie (6, 7). Por lo tanto, se han desarrollado diferentes métodos para tratar de resolver el Problema Inverso para la actividad paroxística, esto quiere decir, que teniendo el registro del paroxismo en el EEG de superficie se puede determinar en qué punto de los hemisferios cerebrales se originó. El método más eficiente para resolver el Problema Inverso en el caso de los paroxismos es la Tomografía Electromagnética de Baja Resolución (LORETA) (8), que consiste en la estimación de norma mínima generalizada de distribución de generadores que usa más fuentes que electrodos. Utiliza un enrejillado tridimensional de fuentes, discreto, dentro del volumen del encéfalo, con un número considerablemente mayor de fuentes que de electrodos (8, 9). Se escoge una solución única minimizando el Laplaciano de la distribución de corriente con la ponderación de la intensidad del generador. Esto permite calcular soluciones al problema inverso, distribuidas con un grado de suavidad espacial, que según la hipótesis matemática en el operador laplaciano, es que puntos cercanos deben activarse de forma similar. Sin embargo, otros autores (10-12) consideran que dada la distancia entre los puntos del cerebro utilizada en esta implementación, la escala de suavidad del LORETA es realmente diferente de la verdadera escala neural. En su implementación el LORETA hace uso del modelo de las tres esferas concéntricas registradas para el Atlas Cerebral Humano de Talairach (13) disponible como RMN cerebral digitalizada del Brain Imaging Center del Instituto Neurológico de Montreal y se tuvo que ajustar el registro entre la geometría esférica vs. la realista de la cabeza, usando las coordenadas 10-20 y 10-10 para los electrodos del EEG (14). La solución inversa está restringida a la sustancia gris cortical y al hipocampo, como se determinó con la digitalización del Atlas Probabilístico también disponible en el Brain Imaging Center del Instituto Neurológico de Montreal, con corrección al espacio de Talairach (15). Las áreas de Brodmannn también se reportan. El volumen intracerebral se divide en 6239 voxels con 5 mm de resolución espacial. Un voxel se marca como sustancia gris si cumple las siguientes tres condiciones: 1) Su probabilidad de ser sustancia gris es mayor que la de ser sustancia blanca, 2) su probabilidad de ser sustancia gris es mayor que la de ser LCR, 3) su probabilidad de ser sustancia gris es mayor del 33 %. Sólo los voxels que pertenecen a la corteza y al hipocampo son usados para el análisis. Un total de 2394 voxels con 7 mm de resolución espacial se producen bajo estas restricciones neuroanatómicas (8, 9). Estas premisas restringen enormemente la detección de posibles fuentes subcorticales en la sustancia blanca, de origen de densidad eléctrica, cuando se le aplican estas restricciones para su consideración como fuente. Un software (datos y ejecutable) implementando LORETA en el espacio de Talairach está a disposición de la comunidad científica en Internet, en la página Web del KEY Institute for Brain Mind Research de Zurich, Suiza, (http:www.z.ch/keyinst/loreta.htm) para calcular la distribución tridimensional de la densidad de corriente en la corteza cerebral, lo cual produjo una utilización amplia de este método con muchos partidarios y algunos detractores (9). El LORETA-S es un método estandarizado, discreto, tridimensional (3D), lineal, para obtener la solución inversa. La forma de estandarización utilizada en LORETA-S permite la localización tomográfica exacta para examinar las fuentes, produciendo imágenes de densidad de corriente con localización exacta, aunque con baja resolución espacial: las fuentes neuronales vecinas pueden estar altamente correlacionadas (8). Las pruebas de su exactitud matemática se describen en las referencias al respecto hechas por el autor (8, 16, 17). Es importante destacar que LORETAs no tiene distorsión en su localización aún en presencia de ruido biológico. En este sentido LORETA-S superó a las primeras versiones de este método (8). Se debe enfatizar que las propiedades de localización de cualquier solución inversa 3D (ej. Tomografía) pueden ser siempre determinadas por errores de localización para examinar puntos de fuente. Si dicha tomografía tiene cero error de localización para dichos puntos de fuentes (16), la localización en cualquier parte del cerebro entonces, excepto por la baja resolución espacial, se localizará correctamente en cualquier distribución 3D arbitraria. Esto es debido a los principios de linealidad y superposición. Estos principios no se aplican a soluciones inversas no lineales, ni se aplican a esquemas que parezcan lineales pero no lo sean, como por ejemplo los ajustes de dipolos. LORETA ha sido combinada con estudios de Resonancia Magnética funcional (RMNf) y estructural, Tomografía de Emisión de positrones, implantación de electrodos profundos en epilepsia y potenciales evocados cognitivos (8, 18-21) con una adecuada validación de su exactitud en localización. En el caso de la versión estandarizada LORETAs, estructuras profundas como la corteza cingulada anterior y la corteza Mesial temporal pueden ser correctamente localizadas (22). En múltiples estudios en los que se han realizado comparaciones con localización en RMN, registros con electrodos intracorticales, estudios con técnicas neuropsicológicas, se ha visto que LORETA determina correctamente la localización de los generadores y su autor hace una extensa revisión de esos logros (9, 23). La principal crítica a LORETA es la suavidad de las soluciones, lo cual localiza las fuentes puntuales con mucha dispersión. Esto es inherente a que se utiliza al Operador Laplaciano Superficial (24) para la estimación de los generadores. Sin embargo, la utilización de LORETA para la localización del origen de los paroxismos ha sido validada con la utilización de electrodos colocados directamente en la corteza, intracorticales y subcorticales, durante intervenciones quirúrgicas en las cuales la localización determinada por LORETA se ha validado con la localización intracraneal (21). Por esto decidimos utilizar LORETA como un método para confirmar el origen de la actividad paroxística de nuestros pacientes. Materiales y métodos Se hizo un corte transversal, descriptivo de una muestra de pacientes tomadas por tres expertos en el área. Se estudiaron 84 pacientes con HTA que fueron clasificados por tres especialistas en Medicina Interna en dos grupos, de acuerdo con las normas del VII Reporte del Comité Conjunto Nacional sobre la Prevención, Evaluación y Tratamiento de la Hipertensión Arterial (25), en: Grupo 1 (PAS=140-159 y PAD=90-99) n=24 pacientes, (14 mujeres, edad media 49±4 años) Grupo 2 (PAS≥160 y PAD≥100) n=60 pacientes, (31 mujeres, edad media 50±6 años). Todos los pacientes estaban controlados de su HTA con tratamiento (que en todos los casos fueron IECAs ó ARA II) y no presentaron signos electrocardiográficos de hipertrofia ventricular izquierda. Los criterios de inclusión en el estudio fueron los siguientes: 1. No tener elementos clínicos sugestivos de una Encefalopatía Vascular Hipertensiva aguda ni crónica. 2. No haber sido nunca paciente de un servicio de Neurología y/o Psiquiatría. 3. No tomar medicamentos anticonvulsivantes ni que afecten al Sistema Nervioso. 4. No tener una enfermedad sistémica diferente de la HTA. 5. No haber tenido fracturas de cráneo ni conmoción cerebral. 6. No estar embarazada o lactando. 7. No ser drogadicto ni alcohólico. Además, se estudió un grupo de sujetos funcionalmente sanos (n=35) (17 mujeres, edad media 47±9 años). Los sujetos controles no presentaron antecedentes de enfermedades sistémicas, neurológicas ni psiquiátricas, intervenciones neuroquirúrgicas, traumatismos cráneo-encefálicos ni antecedentes de dependencia a drogas y alcohol y realizaron las mismas pruebas que los pacientes. Ninguna mujer estuvo menstruando o embarazada en el momento de realizarse el estudio. Todos los sujetos de este estudio tenían, al menos, octavo grado de escolaridad. El registro electroencefalogáfico (EEG) se realizó en vigilia, con los ojos cerrados, en un ambiente con luz natural,en un montaje monopolar utilizando como referencia ambas orejas cortocircuitadas en el panel de electrodos. La frecuencia de muestreo del EEG fue de 200 Hz (5 mseg) con un pasabanda entre 0.5 y 30 Hz. Para la adquisición del EEG se colocaron electrodos de disco de oro con pasta conductora en cada una de las derivaciones del Sistema Internacional 10-20 (26) incluyendo la posición Oz, con una impedancia menor de 5 kohms. Los movimientos oculares se registraron colocando un electrodo 1 cm por encima y por fuera del canto externo del ojo izquierdo y otro electrodo 1 cm por debajo y por fuera del canto externo del ojo derecho, para poder eliminar los segmentos del registro con movimientos oculares. El ECG se grabó durante todo el estudio. El paciente se mantuvo sentado, en una poltrona reclinable, durante el tiempo del registro. Se utilizaron como maniobras de activación la apertura y cierre de los ojos, la Hiperventilación Pulmonar (HVP) durante 3 minutos y la fotoestimulación intermitente a distintas frecuencias. Los pacientes estaban normotensos en el momento del registro. En el análisis de la actividad paroxística se aplicó la Tomografía Electromagnética de Baja Resolución (LORETA) para determinar el área cortical de origen de la actividad paroxística, para lo cual se utilizó el software LORETA-S del Brain Research Institute de Zurich, Suiza, desarrollado para tales propósitos. Para esto se tomaron los segmentos de EEG de cada paciente en la localización donde había un paroxismo detectado visualmente por el electroencefalografista y se “cortaba” el EEG 250 mseg antes y después del inicio y final del paroxismo, en un segmento de EEG que comprendía todas las derivaciones del Sistema Internacional 10-20 incluyendo Oz. Estos segmentos se transformaban a ficheros de texto y después a ficheros lor con el sistema LORETA-S, con el cual se hacía el procesamiento para determinar el origen del paroxismo. En la figura 1 se presenta la imagen tomográfica de LORETA que es solamente de tipo ilustrativa, porque el resultado es un fichero texto donde se dan las coordenadas en los tres ejes (x,y,z) del espacio de Talairach donde se localiza el origen del paroxismo, el área de Brodmann donde está este punto y la circunvolución cerebral a que pertenece. Se cuantificaron las localizaciones que determinaba el LORETA-S según el área de Brodmann y el hemisferio cerebral y se hizo un análisis de Chi cuadrado entre los grupos de pacientes. Los sujetos sanos no presentaron actividad paroxística en el EEG. El nivel de significación estadística se tomó para un valor de p<0.05. Resultados Con esta técnica se encontró que en ambos grupos de pacientes hipertensos los paroxismos se originan principalmente en: 1) el área 17 de Brodmann del hemisferio derecho en la porción más central del cuneus, en la cara medial de los hemisferios (figura 1); 2) en el área 39 de Brodmann del hemisferio izquierdo, en la parte de la circunvolución temporal medial; 3) en el área 37 de Brodmann del hemisferio derecho, en la circunvolución temporal medial; 4) en el área 6 de Brodmann del hemisferio izquierdo (más marcado para los pacientes del grupo HTA 2), en la circunvolución frontal medial y 5) en el área 10 de Brodmann del hemisferio izquierdo en la circunvolución frontal medial (sólo para el grupo HTA 1). (figura 2)Se determinó entonces la irrigación de estas áreas de acuerdo con las normas de Duvernoy (27, 28) para humanos y se pudo apreciar que: 1) El área 17 de Brodmann, en la porción más central del cuneus en la cara medial de los hemisferios, es un área limítrofe (interterritorial) entre la rama leptomeníngea de la Arteria Cerebral Anterior (ACA) y la rama leptomeníngea de la Arteria Cerebral Posterior (ACP). 2) El área 39 de Brodmann, en la parte de la circunvolución temporal medial, es un área limítrofe (interterritorial) entre la rama leptomeníngea de la Arteria Cerebral Media (ACM) y la rama leptomeníngea de la Arteria Cerebral Posterior (ACP). 3) El área 37 de Brodmann, en la circunvolución temporal medial, es un área limítrofe (interterritorial) entre las ramas perforantes de la Arteria Cerebral Media (ACM) y la rama leptomeníngea de la Arteria Cerebral Posterior (ACP). 4) El área 6 de Brodmann, en la circunvolución frontal medial, es un área limítrofe (interterritorial) entre la rama leptomeníngea de la Arteria Cerebral Media y la rama leptomeníngea de la Arteria Cerebral Anterior. 5) El área 10 de Brodmann, en la circunvolución frontal medial, es un área limítrofe (interterritorial) entre la rama leptomeníngea de la Arteria Cerebral Media (ACM) y la rama leptomeníngea de la Arteria Cerebral Anterior (ACA). Discusión Actualmente el método más exacto de localización del lugar de origen de la actividad paroxística es la Tomografía Electromagnética de Baja Resolución (LORETA), método que ha sido validado comparando sus resultados con la localización intracerebral con electrodos implantados intracorticalmente (8,9,17,18,21). La localización que fundamentalmente se determinó con este método correspondió a regiones de la corteza cerebral que son áreas limítrofes entre dos arterias y particularmente entre dos ramas distales de dos de las arterias cerebrales principales: 1) El área 17 de Brodmann, en la porción más central del cuneus en la cara medial de los hemisferios, es un área limítrofe entre la rama leptomeníngea de la ACA y la rama leptomeníngea de la ACP. 2) El área 39 de Brodmann, en la parte de la circunvolución temporal medial, es un área limítrofe entre la rama leptomeníngea de la ACM y la rama leptomeníngea de la ACP. 3) El área 37 de Brodmann, en la circunvolución temporal medial, es un área limítrofe entre las ramas perforantes de la ACM y la rama leptomeníngea de la ACP. 4) El área 6 de Brodmann, en la circunvolución frontal medial, es un área limítrofe entre la rama leptomeníngea de la ACM y la rama leptomeníngea de la ACA. 5) El área 10 de Brodmann, en la circunvolución frontal medial, es un área limítrofe entre la rama leptomeníngea de la ACM y la rama leptomeníngea de la ACM. El área 6 de Brodmann en la circunvolución frontal medial (CFM) está más afectada para el grupo HTA 2 (Fig 2) porque este grupo tiene valores de presión arterial más elevados y por tanto el stress de pared es mayor, lo cual determina una mayor hipoperfusión. La CFM ha sido descrita como un área de origen de generadores de actividad lenta en pacientes con HTA (29) utilizando la Tomografía Eléctrica de Resolución Variable (VARETA), estando más comprometida en los pacientes hipertensos del grupo 2. La CMF es un territorio limítrofe muy sensible a la hipoperfusión que se asocia a la HTA. Las áreas distales siempre son afectadas por la hipoperfusión, por la marcada reducción de la presión hidrostática en los capilares que disminuye la presión neta de filtración hacia los tejidos, pero las áreas limítrofes están entre dos territorios distales y dependen de la difusión tisular desde los capilares de las ramas distales, con lo cual son las áreas más afectadas ante un evento de hipoperfusión como el que se produce en el vasoespasmo arterial y arteriolar cuando la presión arterial está elevada en los pacientes con HTA. Además, se ha reportado que las marcadas oscilaciones en el stress de pared durante el ciclo cardíaco pueden acelerar el proceso ateroesclerótico en las zonas más distales de la circulación carotídea (30). El incremento en la excitabilidad en la corteza cerebral se produce como consecuencia de la reducción del Flujo Sanguíneo Cerebral (FSC) sin isquemia (31). Estos factores determinan que en estas áreas se incremente la excitabilidad que es registrada en el EEG como actividad paroxística. Cualquier grafoelemento que se registre en la superficie requiere de la activación entre 4 y 20 cm2 de corteza (32, 33), pero esta área tiene determinadas características no sólo por la hipoperfusión, sino porque la descarga sincrónica de grandes grupos neuronales que se produce en forma repetitiva, que produce paroxismos intercríticos o interictales y que se mantiene en el tiempo, induce alteraciones en el tejido nervioso que han sido muy estudiadas. Utilizando técnicas de Espectroscopía de Resonancia Magnética, se ha encontrado que en las áreas donde se localiza la actividad paroxística interictal hay disminución de NAA (compuestos N-acetil) sin pérdida neuronal, lo cual se relaciona con stress oxidativo a nivel neuronal (30). Otro estudio (34) demostró stress oxidativo en áreas de generación de paroxismos, con altos niveles de lactato, en la Resonancia Magnética Cerebral de corteza occipital normal en pacientes con paroxismos fotosensibles. Se ha observado que la recuperación de los niveles de NAA en los focos paroxísticos está relacionada con la recuperación de la función mitocondrial y la resolución del stress oxidativo (34). En adición al stress oxidativo, puede haber cambios estructurales ligeros, como es la reducción de las ramificaciones dendríticas, que también pueden contribuir a la disminución del NAA (35). Por otra parte, se ha demostrado que en las zonas donde se origina la actividad paroxística intercrítica, hay hipometabolismo e hipoperfusión que se extiende más allá del área de origen de los paroxismos (37-43). Esto se ha postulado que está asociado a alteraciones funcionales y no estructurales del tejido nervioso (37, 38). Por lo tanto, la condición de generación de actividad paroxística en el tejido nervioso está asociada con alteraciones metabólicas y hemodinámicas que se pueden incrementar si los paroxismos se originan como consecuencia de la hipoperfusión crónica de un área cortical, en nuestro caso por el vasoespasmo producido por la HTA. Conclusiones: 1.- Con la Tomografía Eléctrica de Baja Resolución (LORETA) los generadores de actividad paroxística en los pacientes hipertensos se localizaron en las áreas 17 y 37 de Brodmann del hemisferio derecho, 39, 6 y 10 de Brodmann a predominio del hemisferio izquierdo. 2.- La actividad paroxística del electroencefalograma del paciente con Hipertensión Arterial se origina principalmente en áreas limítrofes de irrigación de las arterias cerebrales. 2.- Técnicas como la Tomografía Eléctrica de Baja Resolución (LORETA) son útiles para el estudio neurofisiológico del paciente con factores de riesgo para desarrollar Encefalopatías Vasculares. Este trabajo fue financiado por: Fondo Nacional de Ciencia y Tecnología (FONACIT) Proyecto S1-97000758. y por el Consejo de Desarrollo Científico y Humanístico (CDCH) Proyecto 09.11.5494.2004 Referencias
Copyright 2010 - Academia Biomedica Digital The following images related to this document are available:Photo images[va10026f2.jpg] [va10026f1.jpg] |
| |||||||||
{kind=link}
{kind=link}