 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 19, No. 3, 2001, pp. 335-357 Parámetros reproductivos y poblacionales de Lutjanus synagris en el Golfo de Paria, Venezuela Reproductive and populational parameters of the lane snapper Lutjanus synagris, Gabriel Gómez, Ramón Guzmán y Ramón Chacón Instituto Nacional de Investigaciones Agrícolas (INIA), CIAE-Sucre/Nueva Esparta. Code Number: zt01019 SUMMARY The size at maturity, fecundity, growth rate and mortality were evaluated for the lane snapper, Lutjanus synagris, which is one of the main supports of the artisanal fishery in the Gulf of Paria. These parameters are fundamental in studies of population dynamics and evaluation of fishery resources. The sample of 1.298 fishes was collected between January and December 1999 from the landings of the artisanal fleet in Güiria. The sizes of fish varied between 210 and 467 mm. The weight length (W) relationship was W = 1,0147*10-4 TL2,64, without significant differences between sex (P>0,05). The size at maturity of 50 % of the population was 368 mm. The greatest reproductive activity took place between July and November. Mean fecundity was 928.854 ovules. The size variation evaluated using FISAT software allowed the estimation of growth parameters as K = 0,20 year -1 and L∞ = 512 mm TL. Mortality was estimated as natural (M=0,27 year–1), fishing (F= 0,39 year-1) and total (Z=0,66 year–1). The estimated rate of exploitation (E) was 0,62 year–1 and the size of first capture (Lc50) was 255 mm. The maximum exploitation rate (Emax), estimated from the yield per recruit model was 0,59 year–1. The size structure of the landings, in which 69% individuals are smaller than the size at maturity, and a current exploitation rate larger than the maximum exploitation rate (Emax) would indicate intense levels of exploitation of the species in the area. Key words: Lane snapper, Artisanal fishery, size of maturity, growth, mortality, exploitation rate RESUMEN La talla de madurez, fecundidad, tasa de crecimiento y mortalidad fueron analizados para el pargo guanapo, Lutjanus synagris, uno de los principales soportes de la pesquería artesanal en el Golfo de Paria, Venezuela. Estos parámetros son fundamentales en estudios de dinámica poblacional y evaluación de recursos. La muestra de 1.298 ejemplares, fue obtenida entre enero y diciembre de 1999 en desembarcos de la flota artesanal de Guiria. Las tallas oscilaron entre 210 y 467 mm. La relación peso – longitud fue P = 1,0147*10-4 *LT 2,64, sin diferencias significativas (P>0,05) entre sexos. La talla de madurez del 50 % fue 368 mm. La mayor actividad reproductiva ocurrió entre julio y noviembre. La fecundidad media fue 928.854 óvulos. Las tallas fueron analizadas usando el programa computarizado FISAT, permitiendo estimar los parámetros de crecimiento K = 0,20 año -1 y L∞ = 512 mm. La mortalidad natural fue M = 0,27 año –1, por pesca F= 0,39 año -1 y total Z = 0,66 año –1. Se estimó la tasa de explotación actual (E) en 0,62 año –1 y la talla de primera captura (Lc50 ) en 255 mm. La tasa máxima de explotación (Emax), derivada del modelo de rendimiento por recluta, fue 0,59 año–1. La estructura de tamaños con 69% de los ejemplares menores a la talla de madurez y una tasa de explotación (E) mayor que la tasa máxima de explotación (Emax), indicarían intensos niveles de explotación del recurso en el área. Palabras clave: Pargo Guanapo, Pesca artesanal, talla de madurez, crecimiento, mortalidad, tasa de explotación INTRODUCCIÓN El pargo guanapo (Lutjanus synagris) es uno de los más importantes representantes de la familia Lutjanidae, recurso que por su valor comercial es uno de los principales soportes de la pesquería artesanal en aguas del Golfo de Paria. Esta especie, cuya distribución comprende desde Carolina del Norte en los Estados Unidos hasta el sureste de Brasil, incluyendo todo el mar Caribe y el Golfo de México (Fisher, 1978; Cervigón, 1993), ha sido escasamente estudiada en Venezuela, citándose los trabajos realizados sobre aspectos biológicos y pesqueros por Méndez (1989) en el Archipiélago Los Roques y por Gómez et al. (1997, 1999) en el Golfo de Paria. Así como estudios sobre aspectos taxonómicos referidos por Cervigón (1993). Desconociéndose aspectos biológicos, poblacionales y de explotación tales como: estructura de tallas, talla de madurez época reproductiva y fecundidad; así como los parámetros de crecimiento, mortalidad y probabilidad o talla de primera captura; términos de referencia en los estudios de dinámica poblacional y evaluación, que aplicados en modelos analíticos permiten inferir sobre el estado y aprovechamiento de los recursos pesqueros. En razón de ello se desarrolló el presente estudio. MATERIALES Y MÉTODOS Los animales utilizados provienen de la pesca artesanal de la flota de Guiria en el Golfo de Paria ( Figura 1), utilizando nasas de una boca, construidas con alambre de gallinero y esqueleto de cabilla de ¼ de pulgada y palangres de anzuelos Nº 5 y 6. Los muestreos se realizaron de enero a diciembre de 1999. Como medida se tomó la longitud total (Lt en mm), considerando el lóbulo mayor de la aleta caudal y el peso (g) del ejemplar, utilizando respectivamente, un ictiómetro de 1 mm de precisión y una balanza de campo de 1,0 g de apreciación.La longitud y el peso de machos y hembras, se relacionó a través de regresión por mínimos cuadrados (Sokal y Rohlf, 1981) de acuerdo al ajuste de la curva de crecimiento relativo en peso P = a * Ltb (Ricker, 1975) y su expresión logarítmica: log (P) = log (a) + b * log (Lt), donde a y b representan constantes de la ecuación de regresión. Las ecuaciones se compararon mediante análisis de covarianza (Snedecor y Cochran, 1971), mientras que el coeficiente "b" se comparó con el valor constante "3" a través de una prueba t – student (Sokal y Rohlf, 1981), para determinar el tipo de crecimiento relativo. Mediante observaciones macroscópicas de las gónadas y usando la escala sugerida por Holden y Raitt (1975) para desovadores parciales, se identificaron los sexos y se determinó la madurez sexual. La evolución y frecuencia de ocurrencia de los diferentes estadios permitió inferir sobre la época reproductiva de la especie. La frecuencia porcentual de individuos maduros en función de la longitud total y el ajuste de los mismos a una ecuación logística, representados gráficamente, permitieron estimar la talla de madurez del 50 % de la población muestreada (Sparre y Venema, 1995).Se determinó la fecundidad con hembras en estadio IV de madurez, usando el método gravimetrico (Bagenal, 1971) para la obtención de los ovocitos y posterior aplicación de la formula indicada por Joseph (1963):
La fecundidad se relacionó con la longitud y con el peso, mediante un análisis de regresión (Sokal y Rohlf, 1981). El crecimiento, la mortalidad y el reclutamiento se estudiaron utilizando los métodos basados en longitudes incorporados en rutinas del paquete computarizado FISAT (Gayanilo et al., 1995). Los parámetros que definen la ecuación de crecimiento ( L∞ y K) de Von Bertalanffy fueron obtenidos del análisis de progresión modal, que involucra la descomposición de frecuencia de tallas por el método de Battacharya (1967), como también de estimaciones iniciales de ambas constantes por el método de Gulland y Holt (1959). Los estimados de L∞ y K se seleccionaron del mejor ajuste del análisis de superficie de respuesta incluido en FISAT. La estimación de mortalidad natural (M) se obtuvo de la ecuación empírica de Pauly (1980) que involucra valores de crecimiento (K), longitud asintótica (L∞) y la temperatura del agua en ºC. La mortalidad total (Z) se obtuvo por la curva de captura linealizada convertida a longitudes a través de la ecuación de Von Bertalanffy; mientras que la mortalidad por pesca (F) se derivó de la ecuación Z= F + M. El análisis de la sub rutina de la probabilidad de captura basada en la ojiva de selección permitió estimar la longitud de primera captura (Lc50) y, conocidos los valores de k , L∞ , Lc50 y M se obtuvo la curva de rendimiento relativo por recluta de Beverton y Holt (1966) que permitió inferir sobre la tasa óptima de explotación o fracción de muertes por pesca (E= F/Z). RESULTADOS Y DISCUSIÓN Estructura de tallas La muestra analizada estuvo representada por 1.298 ejemplares cuyas tallas oscilaron entre 210 y 467 mm de longitud total (Lt), con promedio de 318 mm (Cuadro 1). Un número significativo de los ejemplares (57 %) correspondió a tallas ubicadas entre 250 y 368 mm, mientras que 31 % presentó tallas mayores que 368 mm (talla de madurez del 50 %). Respecto al valor promedio de longitud obtenido (318 mm), este es superior a los valores registrados para la especie por Gómez et al. (1999), Ginez (1982) y Cervigón y Fisher (1979), quienes encontraron longitudes promedio de 288, 200, 250 mm, respectivamente. En Venezuela, los registros de tallas para el pargo guanapo, son relativamente inferiores a las obtenidas en este trabajo; al respecto, Cervigón (1993) señaló para la especie un rango de tallas entre 198 - 300 mm. Por su parte, Méndez (1989) reporta para Los Roques un ejemplar de 446 mm de longitud total y 1.115 g de peso, valores muy próximos a los máximos registrados en este estudio.
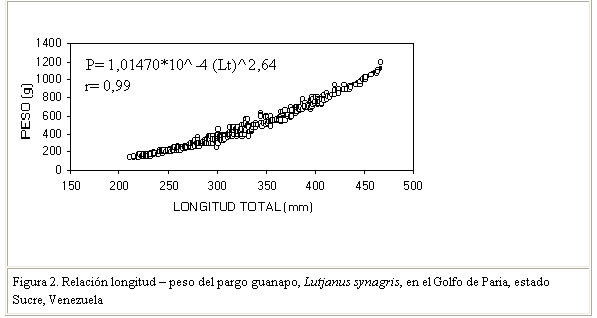
Las variaciones o diferencias en la talla promedio e intervalos de longitud del pargo guanapo en las distintas regiones, estarían relacionadas con las características de cada área en particular, con el tipo de arte utilizado y selectividad del mismo y por ende, con el efecto de la explotación sobre las tallas. En este contexto, una estructura de tallas como la observada en este estudio, donde predominan dos grupos modales con un 69% de tallas inferiores a la talla de madurez del 50 % de la población estaría indicando una actividad pesquera relativamente intensa, lo que pudiera tener efecto en los reproductores y por ende en el reclutamiento. Estas apreciaciones cobran mayor relevancia si consideramos que existe una pesquería artesanal orientada a este recurso, cuyos rendimientos han indicado descensos apreciables en los últimos años (Gómez, 1998). Relación talla – peso Esta relación se realizó con ejemplares de tallas superiores a 210 mm, la cual no mostró diferencias significativas entre sexos (P>0,05 ), por lo que se estimó una ecuación común representada como: P = 1,0147 * 10-4 * Lt2,64 (Figura 2). El valor de la pendiente "b" evidencia un crecimiento relativo en peso alométrico al diferir significativamente (P<0,05) de 3. El crecimiento alométrico ha sido indicado para L. synagris, en aguas de Jamaica y de la plataforma cubana (Thonson y Munro 1974 y Rubio et al. 1985), estos autores encontraron respectivamente, valores de b = 2,94 y b = 2,96 Otros valores de "b" para L. synagris ( b = 2,80, b= 2,64 b= 2,93, b=2,89, b=2,97 y b=2,94) fueron indicados por García-Arteaga et al. (1997) en aguas de Cuba. En Venezuela, este tipo de crecimiento fue señalado para otros Lutjanidos como L. buccanella (Gómez et al., 1995) registrando un valor de b= 2,83; no obstante, para L. vivanus y L. analis, se ha indicado la condición de isometría (Gómez et al., 1994; Palazón y González, 1986) al registrar, respectivamente, valores de b = 3,02 y b= 3,07. Sobre las variaciones del coeficiente de regresión son muchos los factores que pueden incidir, las mismas estarían relacionadas con situaciones de muestreo y con factores ambientales los cuales inducirían cambios espacio temporales, intra específicos e ínter específicos, para este valor.
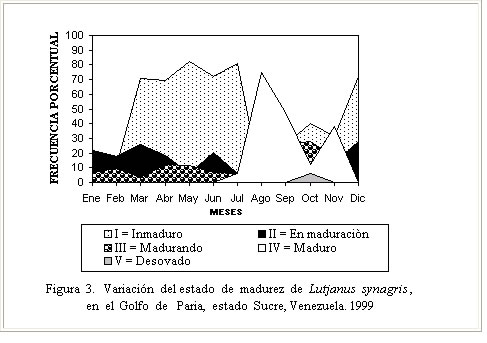
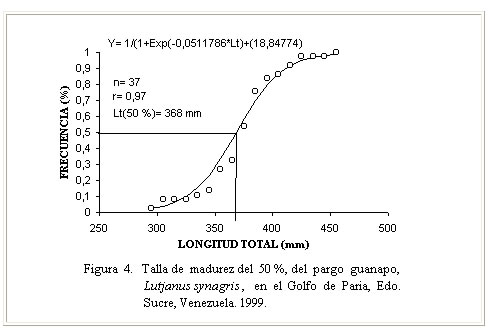
Época reproductiva El análisis del estado de desarrollo gonadal (Figura. 3), permitió inferir que durante todo el año se encontraron animales inmaduros, observándose que de estos un 58 % correspondió al estado de madurez I, un 15 % al estado II y un 13 % al estado III. Los ejemplares en condición de maduros IV representados por 13%, se presentaron a partir del mes julio hasta noviembre, con picos en agosto, septiembre y noviembre, mientras que los desovados en el estado V, 1%, se observaron en octubre. Estos resultados guardan relación con los obtenidos por Gómez et al. (1999) quienes señalaron la mayor actividad reproductiva hacia el segundo semestre del año; sin embargo no se corresponden del todo con los obtenidos por Méndez (1989) para la especie en Los Roques, quien afirmó que el desove ocurre entre marzo y octubre, en dos períodos parciales principales, el primero entre mayo y junio y el segundo entre julio y septiembre; probablemente las condiciones ambientales y características de los ecosistemas estudiados expliquen estos resultados. Es importante destacar que para aguas de Campeche (México) el período reproductivo de L. synagris ocurre de mayo a octubre (Rivera - Arriaga et al., 1996) mientras que para la plataforma cubana, Rodríguez Pino 1962, citado por Borrero et al. (1978) señaló para la especie un desove masivo y sincrónico en primavera; no obstante, este último autor confirmó la presencia de ejemplares por desovar y desovados entre mayo y septiembre, con algunos desoves en noviembre. Claro (1982) indicó que el proceso reproductivo de L synagris se inicia al cumplir el primer año de vida (un 20 a 25 %) y al segundo año el resto se convierte en adulto y comienza el proceso de desarrollo gonadal, el cual se produce en un período corto de tiempo a partir de abril, desde los días de luna en cuarto creciente con la aparición de los primeros grupos en estadios de madurez III, IV y V, hasta septiembre. Druzhinin (1970) por su parte señaló un período de desove entre marzo y septiembre, con picos en junio y agosto. El análisis comparativo de los resultados sobre la época reproductiva indica que los mismos son similares y que las pocas diferencias pudieran estar referidas al área de estudio, patrón de migración y situaciones de muestreo. Talla de madurez Este representa uno de los parámetros a través del cual se pueden hacer inferencias sobre los efectos que pudieran causar los niveles de esfuerzo de pesca aplicados a un determinado recurso. En este estudio, la talla de madurez del 50 % (Figura 4) se determinó en 368 mm de Lt, el ejemplar maduro más pequeño midió 294 mm y el de mayor talla 452 mm. Estos resultados difieren de los obtenidos para la especie por Méndez (1989) en el Archipiélago de Los Roques, al indicar una talla de madurez de 305 mm en hembras, de 290 mm en machos y una talla mínima de madurez de 224 mm . Tal diferencia pudiera estar relacionada con la procedencia de las muestras analizadas (pesca comercial, pesca experimental), selectividad de los artes, actividad pesquera y nivel de explotación, así como con las características intrínsecas de cada área de estudio. Sobre la talla de madurez en Lutjanidos existen algunos registros. Al respecto, Gómez et al. (1994) indicó para L. vivanus de Venezuela un rango entre 540 y 565 mm, Thompson y Munro (1974) para L. buccanella de Jamaica, señalaron una talla de madurez entre 230 y 270 mm y para L. vivanus, entre 500 y 600 mm. Estos autores indicaron que la talla de madurez era mayor en machos que en hembras, lo cual explicaron asumiendo una tasa de crecimiento más rápida en los machos que en las hembras. Fecundidad La determinación de la fecundidad o capacidad de producción de huevos en peces es de gran interés para conocer el ciclo o historia de vida de una especie, tales estudios han sido utilizados además para la identificación y separación de especies, estudios de supervivencia, en acuicultura y para evaluación de stock (Bagenal, 1978; Alvarez-Lajonchere, 1982; Marcus, 1982; Coates 1988, citados por King, 1997). Este parámetro se estableció en base a una muestra de 20 ejemplares sexualmente maduros (estadio IV), cuyo análisis permitió determinar una fecundidad promedio de 928.854 óvulos, con valores entre 510.872 y 1.225.118 óvulos en ejemplares con tallas que oscilaron entre 294 y 452 mm (Cuadro 2). Estos resultados muestran diferencias con los registrados para la especie por Méndez (1989) en el Archipiélago de Los Roques, al indicar una fecundidad absoluta entre 114.726 y 278.298 ovocitos para ejemplares en un rango de tallas entre 321 y 375 mm, respectivamente. Probablemente los rangos de tallas analizados en ambos estudios expliquen las diferencias, sobre todo en los ejemplares de tallas mayores, en los cuales hubo diferencias hasta de 7cm respecto a la máxima talla del rango señalado por este autor.
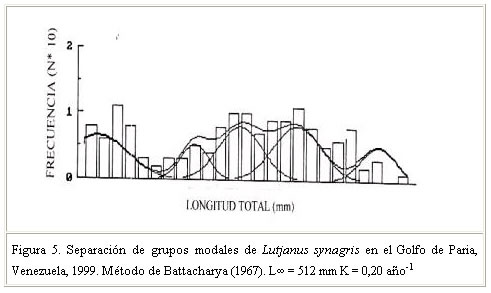
Las referencias en general señalan que los pargos tienen una alta fecundidad. Al respecto, en aguas cubanas, Rodríguez (1962) registró un rango de 347.000 a 995.000 huevos a partir de 6 ejemplares de L. Synagris. Rubio et al. (1985) para la misma especie estimó valores que oscilaron entre 300.000 y 1000.000 de huevos promedio, mientras que Carrillo y González (1981) determinaron una fecundidad media que fluctuó entre 153.712 y 1.125.152 huevos. Pozo et al. (1983) señalaron que la alta fecundidad en los pargos está relacionada con los hábitos de reproducción, con los depredadores de huevos y larvas y con las condiciones ambientales de las zonas de desove, de tal manera que la misma garantiza la sobrevivencia y por ende la renovación y mantenimiento de las poblaciones en el tiempo. Tales antecedentes sobre la fecundidad en los Lutjanidos indican cierta correspondencia con los resultados del presente trabajo. Valores de fecundidad para otros Lutjanidos fueron señalados por Piedra (citado por Druzhinin, 1970), quien indicó un rango de fecundidad entre 100.000 a 1.473.000 huevos en ejemplares de Ocyurus chrysurus de aguas cubanas, y por Moraes (1970) y Gesteira e Ivo (1973) (citados por Correa y Hanson, 1982) que registraron para L. purpureus una fecundidad media de 2.169.000 y 290.000 ovocitos, respectivamente, resaltando una gran diferencia en los valores encontrados. Destacaron estos autores que las diferencias en los valores de fecundidad pudieran relacionarse con la metodología utilizada (gravimetría o volumetría). Se señala además que el estado de madurez gonadal puede influir también en los resultados, toda vez que no se descarta la posibilidad de que se examinen gónadas en las que se haya iniciado la etapa de desove; esto si se considera que las especies tropicales por lo general presentan un patrón de fecundidad fraccionado. Crecimiento El análisis de progresión modal, Battacharya (1967) permitió la separación de cinco cohortes o grupos de individuos de la misma edad (Cuadro 3, Figura 5). Es importante destacar que estos en su mayoría presentaron una talla media inferior a la de madurez (368 mm) lo cual evidencia la intensa actividad pesquera sobre el recurso. Los parámetros de crecimiento constituyen parte importante en los estudios de biología de peces, ya que la determinación e incorporación de los mismos en los modelos analíticos para evaluación de stock, permite inferir en los niveles de explotación y manejo de un recurso (Valle et al. 1997). En este estudio, los valores de L¥ y K fueron 512 cm y 0,20 año –1, respectivamente, que indican un crecimiento lento y una vida longeva de la especie. La proyección de la curva de crecimiento sobre los histogramas de frecuencia mensual (Figura 6) así lo evidencian. Estos resultados son comparables a los obtenidos para L. synagris y otros Lutjanidos del Caribe (Cuadro 4).
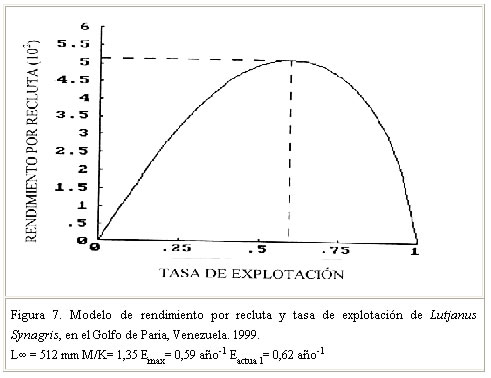
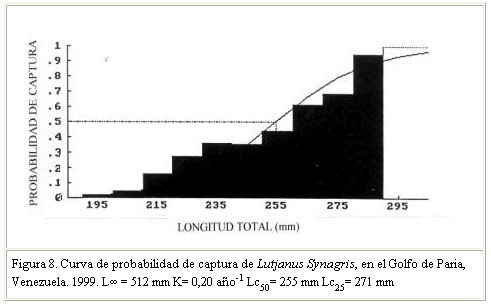
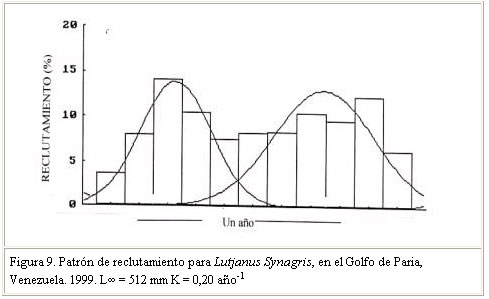
Se menciona que la tasa de crecimiento puede sufrir variaciones en una misma especie, entre períodos anuales y localidades. Al respecto, Sparre y Venema (1995), citado por Guzmán y Gómez (2000), señalaron que tales diferencias pueden atribuirse a cambios de temperatura y disponibilidad de alimentos, asumiendo que existe una relación entre K y la tasa metabólica del animal y esta a su vez con la temperatura. Guzmán y Gómez (2000) señalaron por su parte, que en el caso particular de Venezuela, tales variaciones de temperatura y alimentos guardarían correspondencia con los eventos de surgencias y con la alta productividad y producción biológica de los distintos ecosistemas que conforman la plataforma continental del nororiente del país. Mortalidad Vetter (1988) citado por Guzmán y Gómez (2000), señala que la mortalidad es un parámetro difícil de estimar, por lo que existen diferentes métodos de análisis basados en datos de la captura comercial y en correlación con otros parámetros biológicos como: talla de madurez, tasa de crecimiento, longitud asintótica y longevidad e indica además la importancia de determinar el número de muertes por predación. Manooch and Mason (1984) citado por Acosta y Appeldoorn (1992) señala que los Lutjanidos son especies longevas, de crecimiento lento y tasa de mortalidad natural relativamente baja, apreciación que coincide con los resultados de otros investigadores sobre estudios de parámetros poblacionales en Lutjanidos (Claro y Reshetnikov, 1981; Manickchand-Dass, 1987; Torres-Lara y Chávez, 1987; Torres-Lara y Salas-Márquez, 1990; citados por Rivera-Arriaga et al., 1996) (Cuadro 4). El valor de mortalidad natural obtenido en este trabajo (M = 0,27 año–1) confirma tal apreciación. Al respecto, Claro (1983) señala que una baja mortalidad natural en peces, probablemente esté relacionada con una baja tasa de crecimiento y agrega que esto pudiera estar asociado con una alta intensidad de pesca. Vetter (1988) citado por Guzmán y Gómez (2000) señaló que la tasa de mortalidad natural puede variar de un rango de insignificante a considerable, dependiendo en parte del modelo usado y de los valores seleccionados para los parámetros que involucra la misma, así como también de la forma como se seleccionen los estimados de M. En tal sentido, el valor de M obtenido en este estudio se considera aceptable, basado en la correlación existente entre este parámetro y la tasa de crecimiento, así como en la condición de crecimiento lento y longevidad señalada para los Lutjanidos por los autores referidos. La mortalidad total (Z) determinada fue 0,66 año –1. Este valor, similar al obtenido por Manickchand-Dass (Z = 0,678 año–1) citado por Acosta y Appeldoorn (1992) se considera alto y permitió inferir sobre intensos niveles de explotación del recurso; el mismo estaría directamente relacionado con una alta tasa de mortalidad por pesca y de explotación (F= 0,39 año –1 y E= 0,62). Rendimiento por recluta El modelo predictivo de rendimiento relativo por recluta, permite obtener información sobre los efectos biológicos y/o económicos de la pesca sobre los recursos pesqueros, información que puede servir de base en la toma de decisiones y medidas de administración de los mismos ( Sparre y Venema, 1995). En este sentido, los resultados del modelo (Y/ R) de Beverton y Holt (1966) para esta especie, usando L¥ = 512 mm y M/K = 1,35 indican que la tasa óptima de explotación (Emax), a la cual se alcanza el máximo rendimiento por recluta fue Emax = 0,59 año-1 (Figura 7). El valor de la tasa de explotación actual determinado E= 0,62 año -1 en comparación con el Emax , estaría indicando que el recurso está en una situación de intensa actividad pesquera. La longitud de primera captura (Lc50 =255 mm Lt) estimada (Figura 8), al igual que la talla de madurez del 50% (368 mm) confirmarían los altos niveles de explotación de la especie en el Golfo de Paria. Patrón de reclutamiento El análisis de la información permitió identificar dos pulsos amplios de reclutamiento (Figura 9) lo que coincide con los señalamientos de Acosta y Appeldoorn (1992) para L. synagris de Puerto Rico, quienes reportaron un reclutamiento durante todo el año, con un patrón bimodal. En tal sentido, puede señalarse que el patrón de reclutamiento encontrado en este trabajo, pudiera estar asociado con el período reproductivo de la especie en el área. Al respecto, Gómez et al. (1999) señalaron para L. Synagris en el Golfo de Paria, una intensa actividad reproductiva entre julio y noviembre con un pico en julio que pudiera corroborar tal afirmación; sin embargo, se considera pertinente profundizar más en estudios sobre la dinámica reproductiva de la especie en el área de estudio. CONCLUSIONES
BIBLIOGRAFÍA
Copyright 2001 - Zootecnia Tropical. Free full-text also available online at http://www.ceniap.gov.ve/bdigital/ztzoo/ztindice.htm The following images related to this document are available:Photo images[zt01019f9.jpg] [zt01019f4.jpg] [zt01019f1.jpg] [zt01019f7.jpg] [zt01019f3.jpg] [zt01019f5.jpg] [zt01019f2.jpg] [zt01019f6.jpg] [zt01019f8.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}