 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 19, No. 3, 2001, pp. 371-392 Aspectos reproductivos de la tonquicha (Cynosc ion jamaicensis) en la costa norte de la Península de Paria, estado Sucre, Venezuela Reproductive aspects of the jamaica weakfish, Cynoscion jamaicensis, Luis Marcano y José Alió Instituto Nacional de Investigaciones Agrícolas (INIA), Recibido: 30/5/01 Aceptado: 20/7/01 Code Number: zt01021 SUMMARY The reproductive biology of the Jamaica Weakfish, Cynoscion jamaicensis, was evaluated from a sample of 732 fishes. They were collected on board of shrimp trawl vessels from January to December 1997. Most fishes (84%) were found in the range 201 – 280 mm total length, with an average of 241 mm. General sex ratio did not deviate significantly from a 1:1 proportion; however, males were more abundant during the first quarter of the year. The variation of gonadosomatic index and proportion of individuals in advanced reproductive stage, indicated that reproduction extended from September to February, with a major spawning period between October and December. However, the small number of mature individuals in the samples suggests that spawning took place out of the fishing ground, hence the study area can be considered a zone for feeding, growth and maturation. Fecundity increased with size of fish, with an average absolute fecundity of 197,000 ovocites. The range of ovocites diameter evaluated from 26 ripe gonads was 0.25 to 0.4 mm, with an average of 0.35 mm, suggesting that the species has partial spawnings. Estimated relative fecundity was 635 ovocites/mm total length, and 583 ovocites/g total weight. Average fecundity factor was 244 ovocites. An increase in the mesh size of trawl nets is recommended in order to increase the mean size of captured fish of this and other demersal species in the area. Key words: Cynoscion jamaicensis, sex ratio, spawning time, gonadosomatic index, fecundity RESUMEN Se evaluó la biología reproductiva de la tonquicha, Cynoscion jamaicensis, basándose en una muestra de 732 ejemplares. Estos fueron recolectados a bordo de embarcaciones de arrastre, desde enero a diciembre 1997. La longitud total predominante (84% de los ejemplares) fue 201-280 mm, con un promedio de 241 mm. La proporción general de sexos no se desvió significativamente de una relación 1:1; sin embargo, la proporción de machos fue mayor durante el primer trimestre del año. Las variaciones del índice gonadosomático y de la proporción de ejemplares en avanzado estado de madurez, revelan que la reproducción se extiende desde septiembre a febrero, con un desove más intenso entre octubre y diciembre. El pequeño número de ejemplares maduros recolectados reveló que la reproducción tiene lugar en áreas fuera de la zona de pesca industrial, por lo que el área de estudio debe ser considerada como zona de alimentación, crecimiento y maduración. La fecundidad aumentó con el tamaño del pez, con una fecundidad absoluta estimada en 197.000 ovocitos. El intervalo de diámetros de los ovocitos en una muestra de 26 gónadas maduras fue 0,25 a 0,40 mm, con una media de 0,35 mm, revelando que esta especie tiene desoves parciales. La fecundidad relativa estimada fue 635 ovocitos/mm longitud total y 583 ovocitos/g peso total. El Factor de Fecundidad promedio fue de 244 ovocitos. Se recomienda un aumento en el tamaño de las mallas de las redes de arrastre para elevar el tamaño promedio de los ejemplares capturados de esta y otras especies demersales del área. Palabras Clave: Cynoscion jamaicensis, proporción de sexos, época de desove, índice gonadosomático, fecundidad INTRODUCIÓN La tonquicha, Cynoscion jamaicensis (Vaillant y Bocourt, 1883), es una especie demersal de la familia Sciaenidae, distribuida en la plataforma continental, donde habita principalmente en fondos fangosos, normalmente entre los 18 y 70 m de profundidad (Cervigón, 1993). Se distribuye desde las Antillas Mayores y Panamá, hasta Argentina, incluyendo las costas orientales del Mar Caribe. En el área del Caribe venezolano, es más abundante en la costa norte de la Península de Paria y la Plataforma de Unare-Píritu, donde representa un recurso de primer orden, llegando a constituir parte importante en las capturas de la flota artesanal e industrial de arrastre (Novoa et al., 1998). De acuerdo al interés comercial es una de las especies más importantes para la economía de la región, pues sus volúmenes de desembarque, reportados por las flotas artesanal e industrial de arrastre de la región oriental, oscilaron entre 150 y 600 Tm/año durante la década del 90 (Novoa et al., 1988; Marcano, 2001). Su aprovechamiento en fresco o salado y su utilización en la industria de procesamiento de productos pesqueros, por intermedio del fileteado, le han conferido mayor importancia en los últimos cinco años, ya que el valor agregado, además de generar empleo, ha servido para mejorar la rentabilidad de la flota que la explota, local y regionalmente. Durante el proceso de evaluación de un recurso pesquero, es necesario generar conocimiento acerca de la biología, ecología, dinámica poblacional, bioeconomía y pesquería de las especies que conforman dicho recurso, de manera que, al conjugarse todos estos elementos, permitan determinar su estado de explotación. La información así generada permite que las autoridades administrativas correspondientes puedan implementar medidas de manejo y ordenación, con el objeto de garantizar un aprovechamiento sustentable de los recursos explotados. Aunque la tonquicha representa un componente de primer orden dentro del recurso pesquero de la región oriental, se desconocen muchos de los aspectos antes señalados, específicamente los bioecológicos y poblacionales, lo que ha impedido que se evalué su estado actual de explotación y se puedan implementar medidas de manejo para su explotación racional y aprovechamiento sustentable. Por estas razones, se consideró conveniente desarrollar esta investigación en la que se evalúan aspectos de la biología reproductiva de la especie, relacionados con la fecundidad, índice gonadosomático, época reproductiva y tipo de desove, como parámetros fundamentales en los estudios de evaluación y manejo de un recurso sometido a explotación comercial. MATERIALES Y MÉTODOS Se recolectaron ejemplares a partir de la captura comercial realizada por la flota industrial de arrastre camaronera, en la región oriental. Los mismos fueron capturados entre enero y diciembre de 1997 por técnicos observadores, incorporados a las campañas de pesca realizadas por unidades arrastreras con base en Cumaná. El área de pesca se situó al norte de la Península de Paria, a 4 millas a partir de la Costa entre la localidad de Cangua (62o 55' O y 10o 44' N) y el promontorio de Paria (61o 50' O y 10o 32' 45'' N) (Figura 1). Durante el período de estudio, se procesaron 732 ejemplares de tonquicha. Los ejemplares en el momento de la captura se preservaron en hielo y luego fueron trasladados al laboratorio de pesca Demersal del INIA Sucre - Nueva Esparta, en Cumaná. Los ejemplares fueron medidos usando un ictiómetro de 1,0 mm de apreciación para determinar su longitud (mm) total, y pesados en una balanza digital de 1.0 g de apreciación. El sexo y la madurez sexual de cada ejemplar se determinó de acuerdo a las características morfológicas externas y coloración de las gónadas, según criterios de Holden y Raitt (1975), modificados por Overstreet (1983) para C. nebulosus. La proporción de sexo en la muestra mensual, se analizó mediante una prueba de Ji-cuadrado (Sokal y Rohl, 1996). El índice gonadosomático (IG) se determinó según la fórmula de Kaiser (1973):
donde: PG = Peso total de las gónadas (g) PC = Peso total del cuerpo (g) El tipo de desove de la especie fue determinado a partir de la distribución de frecuencia del diámetro de los ovocitos. La fecundidad se estimó en una submuestra de 26 hembras en estadio IV de madurez (recolectada entre octubre y diciembre), utilizando el método gravimétrico para la estimación del número de ovocitos (Holden y Raitt, 1975). Este método consiste en tomar una submuestra equivalente a un gramo de la parte central de las gónadas maduras (estadíos IV-VI), una vez que se tiene el peso total de las dos estructuras gonadales. La submuestra se coloca en solución Gilson, para la separación de los ovocitos. Una vez que estos quedan libres se colocan en una cámara de Bogorov, para su conteo final bajo un microscopio estereoscópico. El diámetro de los ovocitos fue medido con la ayuda de un ocular micrométrico incorporado al microscopio estereoscópico. Finalmente se aplicó la fórmula indicada por Joseph (1963):
La fecundidad se relacionó con la talla del ejemplar, mediante una ecuación exponencial (Bagenal, 1971):
Mientras que la relación de la fecundidad con el peso total del ejemplar, se describe mediante la ecuación lineal siguiente:
en las cuales, F = fecundidad Se realizó una regresión múltiple por pasos (Sokal y Rohlf, 1996) entre fecundidad, peso total y longitud total del ejemplar, a fin de determinar cual de las dos variables independientes explicaba mejor las variaciones de la fecundidad. El número y diámetro promedio de los ovocitos se utilizó para calcular: a) el índice de fecundidad respecto a la longitud total (IFLt); b) el índice de fecundidad respecto al peso del cuerpo (IFPc); y c) el factor de fecundidad (FF). Estos cálculos se realizaron a través de las siguientes ecuaciones:
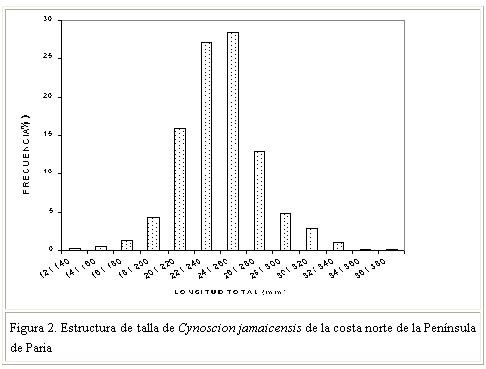
en las que: NO es el número de ovocitos RESULTADOS Y DISCUSIÓN Composición por talla de la población Se analizó la estructura poblacional de la especie sobre la base de una muestra de 732 individuos. La longitud total de la población muestreada osciló entre 136 y 362 mm, con una talla promedio de 242 mm. La estructura de tallas de machos y hembras no difirió significativamente entre sí [(Prueba de Kolmogorov-Smirnov, P>0,05), Sokal y Rohlf, 1996]. La mayor proporción de ejemplares (84, 3%) se ubicó entre 201 y 280 mm de longitud total, destacándose el intervalo 241-260, con el 28% de los ejemplares analizados (Figura 2). Así mismo, se pudo constatar que las clases de menor talla (121-200 mm) también se encuentran presentes en la zona durante todo el año, aunque en menores proporciones (6,4%), al igual que los ejemplares con talla superior a los 281 mm de longitud total, cuyo aporte a la población fue del 9, 3%. Lo primero podría sugerir una renovación permanente de la población a lo largo de todo el año, con la incorporación de individuos juveniles. Por otra parte, la baja proporción de ejemplares de gran tamaño pudiera estar sustentado por lo que han sugerido otros investigadores (Lowe-Mc Connell, 1966; Le Guen, 1971; Santos, 1968; Vazzoler, 1975), al afirmar que en esta especie, al igual que todos los representantes de la familia Sciaenidae, los individuos más grandes permanecen en los estuarios y desembocaduras de ríos hasta completar su proceso de desarrollo gonadal y posteriormente desovar. La fracción de la población con individuos mayores de 281 mm de longitud total, suele estar así fuera del área de pesca, haciéndose menos accesible a la flota industrial de arrastre. Marcano (1988) indicó que la talla de primera madurez de C. jamaicensis es de 261 mm de Lt para las hembras y 287 mm de Lt para los machos. De acuerdo con la estructura poblacional registrada en el presente estudio, la proporción de individuos inmaduros en la población explotada por el sector industrial de arrastre es de 77%. El ejemplar de mayor talla observado en la muestra fue una hembra de 362 mm de longitud total, con un peso de 542 g. Trabajos realizados al sur de Brasil, reportan ejemplares con tallas y pesos muy por debajo de las reportadas en el presente estudio (Santos, 1968; Vazzoler y Braga, 1983). Por su parte, Cervigón (1993), observó peces mayores en talla y peso (392 mm de longitud estándar y 1.000 g respectivamente), encontrando que la talla media comercial se ubicaba entre 250 y 300 mm de longitud estándar, intervalo éste superior al encontrado en el presente trabajo. Tales diferencias pudieran estar asociadas con los sistemas y artes de pesca utilizados en su explotación y con las características hidrológicas y ambientales del área de distribución de la especie. Proporción de sexos Los resultados señalan que de un total de 732 individuos, 382 (52%) fueron hembras y 350 (48%) fueron machos, proporción que no se desvía significativamente de una relación 1:1 (Prueba de Ji-cuadrado, P>0,05). Sin embargo, se observó que la variación mensual de la proporción de sexos presentó diferencias significativas, siendo mayor la proporción de machos en enero y la de hembras en marzo y abril (Cuadro 1).
Las desviaciones significativas a una relación 1:1 en la proporción de sexos observados durante ciertos meses pueden estar relacionados con la conducta reproductiva de la especie y especialmente con la época de desove. La mayor proporción de machos en el área de pesca observada en enero, puede estar asociada a una migración reproductiva de las hembras hacía la costa y áreas estuarinas durante la época de desove, haciéndolas por ende menos vulnerables a la pesquería industrial que se ejerce en el área de muestreo, pero elevando su posibilidad de ser capturadas por la flota artesanal que opera cerca de la costa. Yamaguti (1967) sugirió una explicación similar, en el caso Macrodon ancyclodon en el sur de Brasil. Por otra parte, Vazzoler y Braga (1983), encontraron que en el caso de C. jamaicensis del Brasil, la diferencia en la proporción de sexos era debida a que la explotación realizada por la flota actuaba en ciertas épocas del año en áreas específicas donde el predominio de uno de los sexos era evidente. Antoniutti et al. (1985), sugirieron que las diferencias encontradas a favor de los machos de guaraguara, Plecostomus plecostomus, se debía a que las hembras buscaban áreas de protección durante la reproducción, haciendo más difícil su captura. Colassaco (1987) encontró mayor proporción de hembras de C. jamaicensis en la plataforma Píritu-Unare en cuatro meses del año, sugiriendo que los machos se encuentran en lugares periféricos, de manera que podían quedar más expuestos a los depredadores. La mayor proporción de hembras en los meses de marzo y abril, coincide con un aumento en la actividad de la flota arrastrera en el área, que en busca del camarón (el recurso de mayor valor económico) en la zona, penetra a las áreas costeras donde se reproduce C. jamaicensis. De esta manera la flota industrial ejerce mayor efecto de mortalidad sobre el estrato hembras de la población (Marcano, 2001). La proporción de sexos, parámetro de gran significancia en la estructura poblacional de las especies, varía considerablemente de especie a especie, pero en la mayoría de ellas ésta tiende a ser de 1:1. También difiere entre poblaciones de una misma especie, y varía con la edad de los ejemplares en la misma población. Igualmente puede experimentar diferencias con el período reproductivo, con los grupos de tallas y con la regulación ambiental y la variabilidad genética de las especies (Christiansen y Couseau, 1971). Madurez sexual y época de desove La mayoría (83%) de los ejemplares examinados se encontraban en los estadíos I, II y III de madurez gonádica (Cuadro 2). La baja incidencia de casos de madurez avanzada (IV - VI) en las muestras analizadas, indican que el área de máximo desove de C. jamaicensis se encuentra fuera de la zona de pesca de la flota industrial de arrastre. El desove se produce en el área de estudio en forma parcial durante el período de máxima actividad reproductiva, comprendida entre septiembre y febrero del siguiente año, coincidiendo con la época de baja salinidad del medio, como consecuencia de la descarga del río Orinoco. Resultados similares para Umbrina coroides, fueron encontrados por Gómez (1997) en la misma área de estudio.
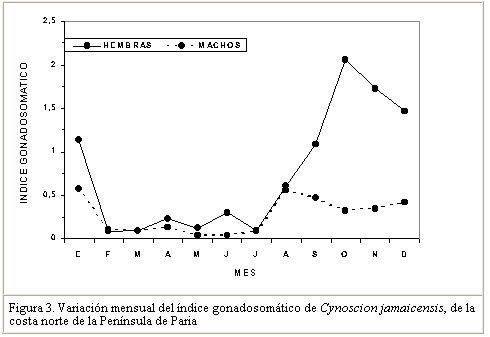
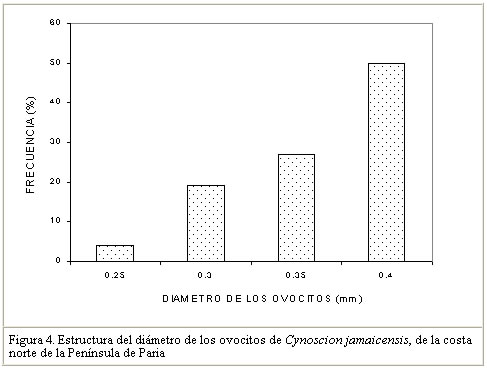
Este resultado corrobora lo observado por Ginés (1972), quien no encontró ejemplares de esta especie en estadíos avanzado de madurez, en muestras procedentes del área de estudio. Por el contrarío, Colassaco (1987), encontró mayor proporción de hembras de C. jamaicensis, en avanzado estado de madurez en la Plataforma de Unare-Píritu. Al respecto, parece existir en esta especie un tipo de migración reproductiva al observarse que, a medida que va avanzando la temporada de reproducción, los ejemplares de gran tamaño se van haciendo menos frecuentes en el área de pesca de la flota industrial de arrastre al norte de la Península de Paria. En este contexto, Santos (1968), en un estudio poblacional de C. jamaicensis, encontró en el sur de Brasil, que la especie migra hacia la costa cuando está próxima al desove y que una vez realizado este, los ejemplares retornaban mar afuera a alimentarse y reiniciar el proceso de madurez. Vazzoler y Braga (1983) observaron que C. jamaicensis se encontraba en la desembocadura de río Grande (Sur de Brasil) para desovar, y que la migración siempre se realizaba desde y hacia la costa y nunca a lo largo de ésta. Vazzoler (1975) también encontró que varias especies de la familia Sciaenidae buscaban las áreas costeras (estuarios y desembocaduras de los ríos) de baja salinidad, donde realizaban sus procesos reproductivos. Así mismo, Lowe-Mc Connell (1966) reporta un patrón similar de migración reproductiva de C. jamaicensis en la desembocadura del río Surinam. Otras especies pertenecientes al mismo género, como Cynoscion nebulosus aparecen en grandes concentraciones reproductivas en las bahías salobres y lagunas del Golfo de México (Tabb, 1971). La totoaba, Cynoscion macdonaldi, penetra al río Colorado (Golfo de California), para desovar, influenciada por los cambios de salinidad del medio (Arvizu y Chávez, 1972). Estos resultados y los del presente trabajo permiten inferir que Cynoscion jamaicensis realiza migraciones reproductivas en busca de estuarios o áreas de baja salinidad durante el proceso de desove. Las fluctuaciones en el período reproductivo de C. jamaicensis, parecen estar justificadas por lo establecido por Nikolsky (1963) quien afirma que las variaciones en el tipo y temporada de desove son determinadas por diferentes condiciones ecológicas durante el proceso de reproducción de los peces. Las especies que representan desoves parciales poseen capacidad de adaptación más elevada, asegurando la perpetuidad de la especie. Basile-Martins et al. (1975) afirman que la época de desove depende de factores físicos y químicos del agua, además de las condiciones propias del organismo. Índice gonadosomático El índice gonadosomático para ambos sexos presenta los máximos valores durante los meses de agosto a enero (Figura 3). Las hembras muestran diferencias mensuales significativas en el desarrollo de este índice (ANOVA de una vía, F = 14,64; P<0,01). A través de una prueba a posteriori de SNK (P<0,01; Sokal y Rohlf, 1996), se demostró que los mayores valores tienen lugar en los meses de octubre a enero del siguiente año. Estos valores coinciden con los meses de mayor frecuencia de ejemplares maduros en el área de pesca, por lo que se podría inferir que existe una relación directa entre los períodos de máxima reproducción y este índice. Se observa además que las hembras presentan valores más altos de este índice (2,058) que los machos (0,572; F = 3,099) (P<0,01). Colassaco (1987) encontró que el período de madurez de C. jamaicensis está comprendido entre septiembre y febrero, en la plataforma Píritu – Unare. Por otra parte, al comparar los valores del índice gonadosomático en hembras de C. jamaicensis del presente estudio al norte de la Península de Paria, con los encontrados por esta autora, se observa que los valores estimados por esta última son más altos (4,67) que los observados en el presente trabajo (2,06). Esta diferencia podría estar asociada a las características de las áreas de pesca, ya que la plataforma Píritu - Unare recibe el aporte de agua dulce de los ríos de los estados Anzoátegui y Miranda, convirtiéndose en hábitat propicio para el desarrollo del ciclo reproductivo. Por su parte, la ausencia de grandes ríos en la costa norte de la Península de Paria, le darían mas bien características de zona para la recuperación gonadal, después del desove ocurrido fuera de esta área. Vazzoler y Braga (1983) encontraron que, de acuerdo a los valores del índice gonadosomático, existía un período de desove un poco más corto para C. jamaicensis en Brasil, comprendido entre septiembre y diciembre. Mientras que Santos (1968) señala, para esta misma especie en Brasil, un período de desove comprendido desde noviembre a febrero del siguiente año. Gómez (1997) observó la misma conducta reproductiva en Umbrina coroides, en el área de estudio. Los resultados de los diversos estudios concuerdan con la existencia de un patrón común del período reproductivo para C. jamaicensis, comprendido desde septiembre a febrero, demostrando con ello una prolongada actividad reproductiva. Tipo de desove Se observó una heterogeneidad en los diámetros de los ovocitos de los ejemplares analizados, encontrándose diámetros desde 0,25 hasta 0,40 mm, con un valor medio de 0,35 mm (Figura 4). La distribución de frecuencia del diámetro de los ovocitos señala que un 23% de ellos poseen diámetros menores de 0,30 mm (inmaduros), el 27% presentaron diámetros entre 0,31 y 0.35 mm (maduración), mientras que el 50% presentaron diámetros superiores a los 0.36 mm (maduros). La variabilidad encontrada en el diámetro de los ovocitos de C. jamaicensis, permite señalar que la especie presenta un desarrollo asincrónico de sus ovocitos, lo cual es indicativo de un desove parcial o fraccionado. Este mismo tipo de desove fue señalado para Micropogonias furnieri, Macrodon ancyclodon y en Umbrina coroides (Vazzoler, 1963, 1969; Yamaguti, 1967; Gómez, 1997). Bagenal (1971) señaló que en un ovario maduro pueden concurrir grupos de ovocitos, representados por ovocitos pequeños blancos y opacos, ovocitos maduros y una tercera porción de ovocitos atrésicos. En cuanto al desove fraccionado, Nikolsky (1963) señala que el mismo es característico de especies tropicales, adoptado como un mecanismo de estrategia que garantiza el potencial reproductivo de las especies. Fecundidad La fecundidad promedio estimada fue de 196.552 ovocitos, con una variación entre 46.354 huevos en un ejemplar de 215 mm de longitud total y 554.400 huevos, en un individuo de 362 mm (Cuadro 3).
Al comparar la fecundidad de C. jamaicensis con otros representantes de la familia Sciaenidae, se encontró que esta es muy superior a la fecundidad estimada en otras especies de esta familia. Vazzoler (1963), en un estudio de fecundidad de Macrodon ancyclodon, estimó que el número de ovocitos maduros listos para el desove estaba alrededor de 178.114, en una hembra de 500 gramos, mientras que Gómez (1997) estimó una fecundidad de 213.247 ovocitos en una hembra de Umbrina coroides de 352 mm de longitud total. Estos resultados indican que Cynoscion jamaicensis es una especie de elevada fecundidad. La fecundidad constituye un parámetro extremadamente lábil sufriendo grandes influencias por efectos de factores intrínsecos (metabólicos) y extrínsecos (ambientales). Varía acentuadamente entre poblaciones distintas de una misma especie dentro de cada grupo y entre períodos sucesivos de reproducción (Nikolsky, 1963). La relación entre la fecundidad y la longitud total fue del tipo potencial, y puede describirse por la ecuación:
Por otra parte, la relación entre la fecundidad y el peso total fue del tipo lineal, representada por la ecuación:
La regresión múltiple por pasos (Sokal y Rohlf, 1996) entre fecundidad, longitud total del pez y el peso total indicó que la longitud total fue la única variable que debe ser considerada por el modelo, y el mismo explicó el 88% de la varianza. El índice de fecundidad en función de la longitud total del pez (IFLt) fluctúo entre 216 y 1.531 ov./mm pez, con un promedio de 635 ov./mm pez. Los valores del índice de fecundidad con respecto al peso del cuerpo (IFPt) oscilaron entre 304 y 1.042 ov./g pez, con una media de 583 ov./g pez. Además, el factor de fecundidad (FF) varió entre 64 y 536 con una media de 244 ovocitos. Colassaco (1987) encontró valores de fecundidad y sus relaciones por encima de los encontrados en el presente estudio. La variación del número de ovocitos por gramo y milímetro de pez en organismos de la misma especie, es característica de un desove parcial o fraccionado. Este tipo de desove aumenta la fecundidad individual, cuando una porción de huevos que se encuentra en los ovarios es expulsada. El desove fraccionado y períodos prolongados de desove son características principales de peces tropicales (Nikolsky, 1963). Cynoscion jamaicensis es la especie de menor talla del género, por lo tanto el número promedio de ovocitos encontrado en esta especie resulta ser relativamente pequeño al compararla con especies del mismo género. Merriner (1975) reportó una fecundidad promedio de 1.725.950 ovocitos, en ejemplares de aproximadamente 400 mm de longitud standard de Cynoscion regalis, determinando una fecundidad relativa respecto al peso mucho mayor a la de C. jamaicensis. Overstreet (1983) encontró en ejemplares de C. nebulosus de aproximadamente 536 g, un promedio de 2.882.172 ovocitos, con un total de 5.360 huevos por gramo de pez. Otros autores (Tabb, 1961; Sundarajab y Suttkus, 1962) estimaron en 1.100.000 el número de ovocitos de un ejemplar de C. nebulosus de 620 mm de longitud standard. De las evaluaciones efectuadas en este trabajo se observa que mientras la pesquería de los adultos de tonquicha se realiza mayormente con trasmallo y filetes de ahorque, la de los juveniles se ejerce por embarcaciones industriales de arrastre. Esto último puede conllevar a un deterioro de la sostenibilidad de la especie y su pesquería en el área, por lo que se recomienda un aumento en el tamaño de las mallas de las redes de arrastre para elevar el tamaño promedio de los ejemplares capturados de esta y otras especies demersales del área. Estos resultados constituyen elementos para conformar una base para realizar la ordenación de la pesquería de esta especie. CONCLUSIONES La estructura de tamaños de la población de Cynoscion jamaicensis de la costa norte de la Península de Paria no fue diferente entre machos y hembras, presentando una talla promedio de 242 mm. La proporción de sexos no fue diferente a la de la esperada 1:1 (P>0,05). Las variaciones del índice gonadosomático y de la proporción de ejemplares en estadío de madurez avanzado, indican que la especie tiene un prolongado período de reproducción, el cual se extiende desde noviembre hasta febrero del siguiente año, con desoves más intensos entre octubre y diciembre. La distribución del diámetro de los ovocitos indica un desove fraccionado en esta especie. Este tipo de desove puede ser una de las causas del prolongado período de reproducción. La baja incidencia de hembras en estadíos de madurez gonadal avanzado, así como la baja proporción de ejemplares en desove y desovados en el área de estudio, indican que C. jamaicensis se reproduce fuera del área de pesca de la flota industrial de arrastre. Esta última pareciera ser un área de maduración y crecimiento por el alto porcentaje de ejemplares en estadíos de madurez I y II encontrados en la muestra. AGRADECIMIENTOS Deseamos agradecer la colaboración de los técnicos del INIA, M. Márquez, R. Chacón, A. Urbaneja, G. Vizcaíno y R. Marcano, durante la realización de este trabajo, así como de N. Romero, L. Ortiz y C. Torrens por su participación como observadores a bordo y la recolección de las muestras biológicas. Así mismo, agradecemos la cooperación los señores Jaime Rodríguez, Giuseppe Spinalli, Luis Garayar y Tomasso Natoli, empresarios de la pesca industrial de arrastre, quienes permitieron las labores de investigación a bordo de sus unidades de pesca. Por último, queremos agradecer a G. Gómez, R. Guzmán y D. Altuve por las recomendaciones en la redacción del manuscrito. BIBLIOGRAFÍA
Copyright 2001 - Zootecnia Tropical. Free full-text also available online at http://www.ceniap.gov.ve/bdigital/ztzoo/ztindice.htm The following images related to this document are available:Photo images[zt01021f3.jpg] [zt01021f4.jpg] [zt01021f2.jpg] [zt01021f1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}