 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnica Tropical, Vol. 20, No. 1, 2002, pp. 89-103 Biometria y talla de primera madurez de la tonquicha, Cynoscion jamaicensis, de la costa norte de la península de Paria, estado Sucre, Venezuela Biometry and size of first maturity of sea trout, Cynoscion jamaicensis, in the northern coast of Paria península, Sucre state, Venezuela Luis Márcano, José Alió y Douglas Altuve Instituto Nacional de Investigaciones Agrícolas (INIA), Centro de Investigaciones Agrícolas de Sucre y Nueva Esparta, Aptdo. 236. Cumaná, Sucre 6101, Venezuela. Correo electrónico: ciapes@sucre.udo.edu.ve. Recibido: 17/9/01 Aceptado: 20/12/01 Code Number: zt02007 SUMMARY Sea trout, Cynoscion jamaicensis, is an important fish in artisanal and industrial trawl fisheries of Northern Paria Peninsula, and knowledge of its population parameters is required to establish management plans of its exploitation. The sample consisted of 852 individuals, collected on board of industrial shrimp trawlers between January and December 1997. The biometry and macroscopic gonad maturity stage of each individual were evaluated. Average size was similar between sexes, with a general average of 241 mm of total length (TL). Parameters of the weight – length relationship did not differ significantly between sexes, and a common regression was estimated as W = 5,147 x 10-6 TL3,151, in which the relative growth factor (b) was significantly greater than 3. The condition factor (Kn) remained near or greater than 1 during the study period, with the largest values observed in March and August-December, corresponding to periods of greatest gonad development and sizes of individuals. The minimum size of first maturity were registered at 153 and 206 mm TL for females and males, respectively. The size at which 50% of the population is mature did not differ significantly between sexes, and was estimated at 251 mm TL. Beyond 265 mm TL, more than 75% of individuals were sexually mature. Considering that the size at which the population reaches maturity is larger than the mean size of the exploited population by the industrial shrimp trawlers, it is expected that the species' density undergoes severe deterioration in the zone, if indiscriminate capture of individuals is maintained. Key Words: Cynoscion jamaicensis, biometry, lenght-weigth relationship, condition factor, size of first maturity RESUMEN La tonquicha, Cynoscion jamaicensis, es un pez importante en las pesquerías artesanales e industriales de arrastre al Norte de la Península de Paria, y se requiere conocer parámetros poblacionales para establecer planes de manejo de su explotación. Se analizaron 852 ejemplares, recolectados a bordo de embarcaciones de arrastre, entre enero y diciembre 1997, evaluándose la relación longitud (Lt) – peso (W), el factor de condición fisiológico (Kn), estadíos y talla de primera madurez gonadal de la población. La talla de los ejemplares fue similar entre sexos, con un promedio general de 241 mm Lt. Los parámetros de la relación longitud - peso no difirieron significativamente entre sexos, estimándose una regresión común W = 5,147 *10-6 Lt3,151, en la cual el factor de crecimiento relativo (b) fue significativamente mayor que 3. Kn se mantuvo cercano o mayor a 1, con valores máximos en marzo y agosto–diciembre, en concordancia con los períodos de mayor desarrollo gonadal y talla de los ejemplares. La talla mínima de primera madurez fue de 153 y 206 mm Lt para hembras y machos, respectivamente. La talla de madurez del 50% de la población no fue significativamente diferente entre sexos y se estimó en 251 mm Lt. Por encima de 265 mm de Lt, más del 75% de los individuos está sexualmente maduro. Considerando que la talla de madurez poblacional supera la talla promedio de la población explotada por las embarcaciones industriales de arrastre, se estima que la densidad de la tonquicha puede sufrir un considerable deterioro en la zona, de continuarse con una pesquería indiscriminada de los ejemplares. Palabras clave: Cynoscion jamaicensis, biometría, relación talla-peso, factor de condición, talla de primera madurez. INTRODUCCIÓN La familia de peces Sciaenidae está conformada por una gran variedad de géneros y especies, cuyo hábitat se encuentra particularmente en fondos fangosos o fangoso-arenosos, cercanos a las desembocaduras de los ríos. Algunas especies se localizan a profundidades considerables, hasta 135 m, mientras que la mayoría se distribuyen muy cerca de la costa (Cervigón, 1993; Márcano, 1988, 2001). En la región Centro Occidental del Océano Atlántico existen aproximadamente 21 géneros diferentes, siendo Cynoscion el más abundante, con un total de 11 especies (Fischer, 1978); ocho de estas especies se encuentran distribuidas en la Costa Atlántica de Sur América desde Colombia hasta Argentina (Cervigón y De Lima, 1972) y 13 en el Pacífico Oriental (McPhail, 1958). En aguas venezolanas esta familia es muy abundante en la plataforma de Margarita-Los Testigos, en el Golfo de Venezuela, en la región Atlántica Venezolana y la plataforma Unare-Píritu, donde forma parte importante del recurso pesquero, aportando mas del 6% de la producción pesquera marítima nacional (Novoa et al., 1998). Cuatro especies (Macrodon ancylodon, Micropogonias furnieris, Cynoscion virencens y C. jamaicensis) aportan la mayor contribución, cuya importancia comercial se ha venido incrementando significativamente debido al uso industrial, que a través del fileteado, se le ha venido dando desde hace mas de ocho años, particularmente en la región oriental del país (Novoa et al., 1998; Márcano, 2000). Cynoscion jamaicensis fue descrita por primera vez en Jamaica por Gunter en 1860 como Otholitus nothus. Posteriormente en 1883 Vaillant y Bocourt la identificaron con el nombre con que actualmente se conoce (Randall y Cervigón, 1967). Cynoscion jamaicensis es una especie demersal de la zona nerítica de la plataforma continental, donde habita principalmente en fondos fangosos, normalmente entre 18 y 70 m de profundidad (Cervigón, 1993). Se distribuye desde las Antillas Mayores y Panamá, hasta Argentina, incluyendo las costas orientales del Mar Caribe. En el área del Caribe oriental venezolano, es más abundante en la costa norte de la península de Paria, donde se la explota artesanalmente mediante el uso de varios artes de pesca, e industrialmente mediante el sistema de arrastre camaronero. Con éste último sistema se obtiene las mayores capturas en la zona (Novoa et al., 1998; Marcano, 1988, 2001). Actualmente se considera un recurso de primer orden, llegando a constituir parte importante en las capturas comerciales de la flota arrastrera local, conjuntamente con los crustáceos, moluscos y otras especies de peces de considerable valor dentro de la economía regional y nacional. Debido a que es un recurso de importancia pesquera para la región es necesario conocer su biología, dinámica poblacional y aspectos más importantes de su pesquería a fin de proporcionar elementos fundamentales para la ordenación de su aprovechamiento, de manera racional y responsable. El grado de desconocimiento sobre estos aspectos es tal, que en el cuadro de conocimientos de aspectos biológicos y pesqueros de las principales especies de interés comerciales indicadas por la Dirección General Sectorial de Desarrollo Pesquero, actualmente SARPA (MAC, 1978), aparece sin ningún tipo de estudios de los que allí se señalan. Actualmente, aunque se conocen algunos trabajos acerca de la especie (Colassaco, 1987; Marcano, 1988; Mendoza et al., 1994; Márcano y Alió, 2001), es muy limitado el conocimiento que se tiene de los aspectos más fundamentales de su biología, reproducción, biometría, dinámica poblacional y pesquería. Por estas razones, se describen en el presente trabajo aspectos relacionados con la biometría (composición por talla, relación talla-peso, tipo de crecimiento y factor de condición fisiológico), madurez y talla de primera madurez de C. jamaicensis, como parámetros fundamentales en los estudios de evaluación, y en la toma de decisiones para la implementación de regulaciones de manejo y ordenación de su explotación comercial. MATERIALES Y MÉTODOS Se recolectaron ejemplares a partir de la captura comercial realizada por la flota industrial de arrastre camaronera, en la región oriental. Los mismos fueron obtenidos durante el período comprendido de enero a diciembre de 1997, por técnicos observadores, incorporados a las campañas de pesca realizadas por unidades arrastreras con base en Cumaná. El área de pesca fue descrita anteriormente por Márcano y Alió (2001). Posterior a la captura, los ejemplares se conservaron en hielo y luego fueron trasladados al laboratorio de pesca demersal del CIAE Sucre-Nueva Esparta, en Cumaná. En el laboratorio, los ejemplares fueron medidos usando un ictiómetro de 1,0 mm de apreciación, para determinar su longitud total, y pesados en una balanza digital de 1,0 g de apreciación. El sexo y la madurez sexual de cada ejemplar se determinaron de acuerdo a las características morfológicas externas y coloración de las gónadas, establecidas por Holden y Raitt (1975), modificada por Overstreet (1983) para C. nebulosus. Biometría Los aspectos biométricos de la especie fueron analizados en función a la composición por talla y peso, relación talla-peso y factor de condición de los ejemplares. a) La longitud total y el peso de los ejemplares procesados permitió determinar la composición por talla y peso de la muestra, determinándose las tallas y pesos mínimos y máximos, así como el promedio de cada parámetro por sexo. b) La estimación de la relación de la longitud total (Lt) con el peso (P) para cada sexo se realizó mediante el ajuste de estos parámetros a la ecuación alométrica P^ = a * Ltb (Teissier, 1948; Ricker, 1975). Las pendientes "b" e interceptos "a" de ambas ecuaciones se compararon a través de un análisis de covarianza para establecer posibles diferencias entre las mismas. Una prueba t-Student a la constante "b", permitió determinar el tipo de crecimiento de la especie (Zar, 1996). Si el valor de la constante "b" (pendiente) es igual a tres (b=3) el crecimiento es isométrico. Caso contrario el crecimiento es alométrico, mayorante (b>3) o minorante (b<3; Ricker, 1975). c) Factor de condición fisiológico (Kn): Este factor se determinó mediante la ecuación Kn = P/P^ descrita por Le Cren (1951), donde P representa el peso total observado del individuo y P^, el peso predictivo que debería tener, según la talla promedio de su grupo. En esencia, P^ = a * Ltb, representa el peso predictivo, donde "a" y "b" son constantes obtenidas de la relación talla-peso. Madurez sexual y talla de primera madurez La talla mínima de primera madurez sexual se define como la menor longitud a la cual al menos un ejemplar evidencia un desarrollo gonadal máximo. Este parámetro se determinó registrando las tallas de los organismos más pequeños con las gónadas en pleno desarrollo (Pitt, 1964). La talla de primera madurez del 50% (L50) de la población se estimó para cada sexo, ajustando la proporción de individuos maduros en cada intervalo de tallas a una función logística, de acuerdo a los criterios establecidos por Somerton (1980), Gaertner y Laloe (1986) y Sokal y Rohlf (1996):
donde: Y: proporción de individuos maduros X: Marca de clase de la longitud de referencia A y B: constantes del modelo e: base del logaritmo neperiano Línea rizando la ecuación anterior, mediante una transformación logarítmica:
Se estimaron los parámetros A y B por regresión de mínimos cuadrados (Sokal y Rohlf, 1996). Así, la longitud a la cual el 50% de la población se encuentra sexualmente madura (X0.5) corresponde a:
La ecuación original se modifica para incluir X0.5, obteniéndose:
lo cual hace que los estimados de A y X sean independientes y la ecuación tenga un sentido más biológico. RESULTADOS Y DISCUSIÓN Biometría
Se evaluaron 852 ejemplares, de los cuales 452 (53%) fueron hembras y 400 (47%) fueron machos. La longitud total de las hembras osciló entre 138 y 362 mm, con un promedio de 240 mm, mientras que el peso fluctuó entre 27 y 542 g, con un promedio de 174 g. Los machos presentaron un intervalo de longitud total entre 175 y 345 mm, con una media de 243 mm, mientras que el peso osciló entre 54 y 484 g, con promedio de 180 g. La longitud total promedio de la muestra total fue de 241 mm (Cuadro 1). Trabajos previos sobre C. jamaicensis, han señalado tallas superiores a las observadas en el presente estudio. Al respecto, Cervigón (1993) encontró peces con talla de 392 mm de longitud total, señalando además que la talla media comercial de la especie se ubicaba alrededor de los 300 mm de longitud estándar. Otros trabajos realizados al sur de Brasil, Suriname y Guyana, reportan ejemplares con tallas y pesos muy inferiores a las del presente estudio (Dos Santos, 1968; Vazzoler y Braga, 1983; Uyeno et al., 1983; Lowe-McConnell, 1966). Tales diferencias entre las tallas en distintas regiones geográficas, pudieran estar asociadas con los sistemas y artes de pesca utilizados en la explotación, así como con las características ambientales de la zona de distribución de la especie. De esta manera se esperaría obtener ejemplares de mayor talla promedio en localidades costeras y cercanas a deltas, donde suele ocurrir el desove y la abundancia de ejemplares maduros debería ser más elevada. Adicionalmente, diferentes intensidades de pesca entre regiones geográficas pueden influir sobre el tamaño de los ejemplares encontrados en ellas. El efecto causado por una pesquería sobre una población o stock, cuando es explotada intensamente, o cuando los artes no tienen efecto selectivo sobre la fase explotada, es la reducción gradual de la talla promedio de la población con el tiempo (Ricker, 1979). Este efecto se produce también cuando se ejerce presión pesquera sobre el estrato joven de la población. Relación talla - peso El análisis de los datos de longitud y peso permite, primeramente, describir en forma matemática la relación entre la talla y el peso, de tal manera que se pueda estimar uno a partir del otro; medir la variación del peso esperado para un pez o grupos de peces, como indicador de las condiciones alimentarías y desarrollo gonadal (Le Cren, 1951; Almeida et al., 1995; Sparre y Venema, 1995). Igualmente, la información de la relación talla-peso se utiliza para la comparación de grupos de peces (Caillouet, 1993), para estimar biomasa y evaluar stocks, entre otros (Gulland, 1983; Entsua-Mensah et al., 1995; Kulbicki et al., 1993; Beyer, 1987, 1991; Benedito et al., 1997). En el Cuadro 2 se presenta el resumen del análisis de la relación talla-peso para C. jamaicensis. La ecuación que relaciona la longitud y el peso en individuos machos quedó establecida como:
Mientras que para las hembras fue:
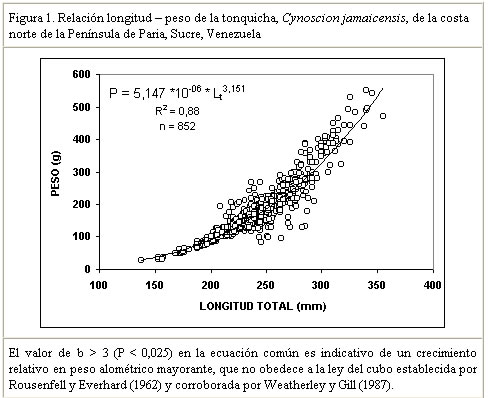
La comparación de las pendientes (t Student=1,12; P>0,05) e interceptos (t Student=1,68; P>0,05), indicó que no existían diferencias significativas entre las ecuaciones para machos y hembras, por lo que se estimó una ecuación común para ambos sexos: W = 5,147 x 10-6 x Lt 3,151 (Figura 1). Cuadro 2.
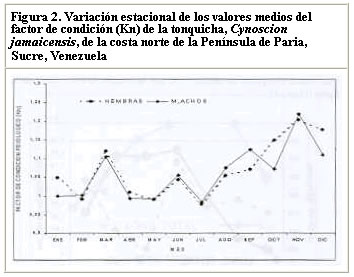
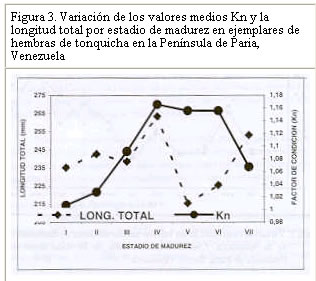
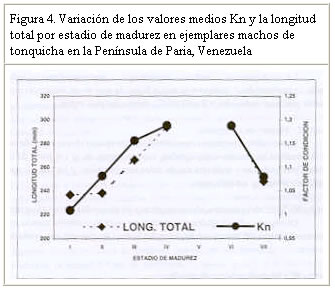
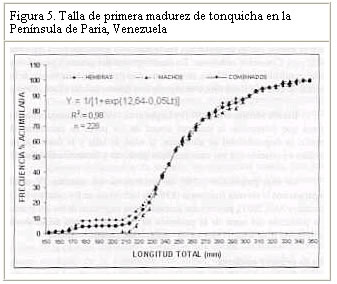
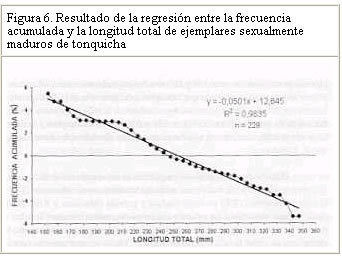
Resultados similares (crecimiento alométrico mayorante) fueron reportados por Nomura (1962) para ésta misma especie al Sur de Brasil. Por su parte, Dos Santos (1963) reportó un crecimiento alométrico minorante (b = 2,98) sin observar diferencias significativas entre machos y hembras de Cynoscion jamaicensis, también al Sur de Brasil. Colassaco (1987), igualmente reportó un crecimiento alométrico minorante (b = 2,89), para esta misma especie en la plataforma Unare – Píritu en Venezuela. Es posible que C. jamaicensis de la plataforma Píritu-Unare sea una unidad de stock distinta a la estudiada en el presente trabajo, por cuanto la barrera ecológica representada por la fosa de Cariaco, que separa los dos habitats de la especie, estaría dificultando el intercambio genético entre las dos poblaciones. Le Cren (1951) estableció que además de diferir el valor de b entre especies y habitáculos, también se pueden encontrar variaciones en épocas diferentes, debido al sistema de muestreo empleado. Poco se conoce acerca de la relación talla-peso de peces explotados comercialmente en la región oriental. La creciente importancia de la industria pesquera de la región sugiere que tales informaciones puedan ser utilizadas para propósitos biológicos o para estimaciones estadísticas de pesquería. Factor de condicion fisiológico (Kn) El factor de condición fisiológico (Kn) en peces varía de acuerdo con las influencias de los factores fisiológicos y ambientales, siendo conocidas sus fluctuaciones durante un ciclo reproductivo, particularmente en ejemplares adultos (Bagenal, 1957). Esto se interpreta como indicación relativa del bienestar del pez y su comportamiento durante el proceso reproductivo, y se utiliza para tener una idea de la conveniencia del ambiente, o para la comparación entre peces de zona geográficas distintas (Rounsefell y Everhard, 1962; Almeida et al., 1995; Vazzoler, 1996; Anibeze, 2000). Los valores promedio mensual del factor de condición fisiológico (Kn) a lo largo del período de estudio fluctuaron entre 0,98 y 1,22 para ambos sexos de la población de Cynoscion jamaicensis, estimándose un valor promedio anual de 1,069 y 1,060 para hembras y machos respectivamente, indicando con ello las buenas condiciones fisiológicas de la especie en el área de estudio (Figura 2). Los valores máximos de este factor fueron observados durante el mes de marzo y en el período agosto-diciembre, coincidiendo con los meses de mayor desarrollo gonádico y máximas tallas de la especie. El análisis de correlación de Pearson entre los comportamientos mensuales del Kn para los dos sexos, fue altamente significativo (r = 0,93; P < 0,01), demostrando con ello que el proceso reproductivo de C. jamaicensis es realizado por ambos sexos de manera simultánea, como suele ocurrir en la mayoría de las especies de peces marinos (Le Cren, 1951; Rounselfell y Everhard, 1962; Vazzoler y Braga, 1983; Isaac-Nahum y Vazzoler, 1983; Antoniutti et al. 1985; Braga, 1986). El factor de condición (Kn) aumentó a medida que la gónada de los ejemplares maduraba, siendo máximos sus valores en los estadios IV y V. A partir del estadio VI, cuando la especie inicia el proceso de desove, el factor de condición desciende hasta alcanzar su valor más bajo cuando está completamente desovado (estadio VII). La relación existente entre la talla y el Kn presentó un comportamiento similar al observado entre este y los estadios de madurez, observándose que el Kn aumenta con la talla del ejemplar. No obstante este comportamiento, en los estadios V y VI de las hembras, el valor del Kn se mantiene como uno de los más elevados a pesar de que la talla promedio en ellos son las menores observadas durante el periodo de estudio (Figura 3). Ello obedece a que la talla promedio de los ejemplares en esos estadíos resultó ser muy inferior a la talla promedio general (241 mm) de la especie, además de que él número de ejemplares encontrado en dichos estadios fue relativamente bajo. En los machos se aprecia una relación más clara entre el aumento del factor de condición, la madurez gonadal y la talla de los ejemplares. Sin embargo, no se encontraron ejemplares en estadio V, lo que interrumpe la continuidad de la figura (Figura 4). León (1982) señaló que los factores que determinan la condición de los peces pueden ser: genéticos, calidad de alimentación, hábitos alimenticios, estado sanitario, etapas de desove y talla por edad. En este sentido, Vazzoler y Braga (1983) y Vazzoler (1996) señalaron que el factor de condición (Kn) está íntimamente relacionado con el ciclo reproductivo de los peces, siendo su variación explicada por la intensa actividad de las gónadas. Generalmente el factor de condición tiende a ser mínimo durante la época de desove, debido a la cantidad de energía gastada en el acto que precede a esta función fisiológica, y que es liberada en forma de óvulos (Htun-Han 1978; Braga, 1986). Posteriormente, aumenta gradualmente a medida que se van normalizando las funciones metabólicas después de la postura (Joakimsson y Hempel, 1974). Diferencias en el factor de condición han sido interpretadas como medidas de varios caracteres biológicos tales como almacenamiento de grasa, conveniencia del medio ambiente, salud de un individuo o desarrollo gonadal (Le Cren, 1951). Tapia et al. (1988 a y b) señalaron que el factor de condición (Kn) está asociado a fenómenos biológicos y ecológicos del reclutamiento, crecimiento, madurez y desove. Los valores bajos coinciden con la época de desove y presencia de individuos de tallas grandes, mientras que los valores altos coinciden con el reclutamiento de los individuos juveniles y subsiguiente maduración de la población. Madurez sexual Morfológicamente las gónadas de esta especie son estructuras pares, situadas en posición anterodorsal, las cuales se extienden a lo largo de la cavidad celómica, donde se fijan fuertemente por mesenterios delgados y transparentes. Las principales características macroscópicas, correspondientes a cada estadío de madurez de C. jamaicensis, según la escala para peces tropicales con desoves parciales de Holden y Raitt (1975), fueron las siguientes: Estadío I.- Inmaduros o vírgenes. Este estadío se encuentra representado en ejemplares jóvenes que aún no han desovado. Los ovarios son pequeños, ocupan menos de la 1/4 parte de la cavidad, son turgentes y translúcidos, de color rosado pálido, y los óvulos son difíciles de observar a simple vista. Estadío II.- Inicio de la maduración de vírgenes y de desovados en recuperación. Ovarios firmes, alargados y turgentes, de color ligeramente rosado, en ocasiones la irrigación vascular es evidente, los huevos se observan con ayuda de una lupa. Estadío III.- En maduración o desarrollo. Ovarios aumentados de tamaño, de coloración rosada, con mayor irrigación vascular. Los huevos son visibles a través de la túnica, de apariencia granulosa. Estadío IV.- Desarrollado. Los ovarios son considerablemente grandes, ocupando más de las 2/3 partes de la cavidad del cuerpo. Presentan una coloración rosado amarillento, como resultado de la formación del vítelo, con gran cantidad de vasos sanguíneos. Los huevos son opacos y se hacen más visibles a simple vista. Estadío V.- Grávida. Los ovarios ocupan la mayor parte de la cavidad del cuerpo, presentando vasos sanguíneos abundantes y prominentes sobre la superficie de coloración amarillo brillante. Los óvulos son completamente redondos y traslúcidos. Estadío VI.- Desove. Los huevos fluyen con una ligera presión en el abdomen. Casi todos los huevos son translucidos, con muy pocos opacos en el ovario. Estadío VII.- Desovado. Ovarios flácidos, sin encontrarse vacíos completamente; no hay presencia de huevos opacos en el ovario. Este último es de coloración violácea, con irrigación sanguínea moderada, ocupando aparentemente la mitad de la cavidad del cuerpo. Las características de las gónadas observadas en el presente estudio tienen el mismo patrón de desarrollo (siete estadíos) señalados por Overstreet (1983) para Cynoscion nebulosus. En otras especies del género Cynoscion se señalan resultados muy similares, observándose un desarrollo asincrónico de los ovocitos, lo cual es característico de especie con desove parcial (Tabb, 1961). Bassile Martins et al. (1975) y Lagler et al. (1977) señalan que entre los factores que determinan la madurez sexual de los peces se encuentran la herencia, la disponibilidad de alimentos, la edad, la talla y la fisiología del individuo en relación con sus características genéticas y factores ambientales. La baja proporción (17%) de hembras en los estadíos IV a VI, comparado con la elevada frecuencia (83%) de individuos en los estadíos I y III (Marcano y Alió, 2001), parece estar asociado a una migración de los integrantes de la población del norte de la península de Paria hacia otras áreas, donde posiblemente tiene lugar el proceso de maduración gonadal y desove. Talla de primera madurez Se observó que las hembras inician el proceso de madurez a menor edad y talla que los machos. En este sentido, la talla mínima de primera madurez de las hembras de C. jamaicensis se estimó en 153 mm LT, mientras que la de los machos fue 206 mm LT. La estimación de la longitud total a la cual madura el 50% de la población muestreada (L50) fue de 249 mm (con un intervalo de confianza del 95% de 234 a 264 mm) para las hembras y 259 mm (con un intervalo de confianza de 95% de 233 a 286 mm) para los machos (Figura 5). La superposición de estos intervalos de confianza revela que los valores de L50 para machos y hembras no difieren significativamente, por lo que se estimó un valor común de 251 mm (y un intervalo de confianza del 95% de 237 a 268 mm de LT). De hecho, la estructura de tallas de hembras y machos maduros no difiere significativamente (Prueba de Kolmogorov – Smirnov, P>0,05). El valor de L50 común calculado por medio de la regresión de la curva linearizada de frecuencia acumulada vs. longitud total (251 mm; Figura 6), difiere del estimado gráficamente (247 mm), lo cual es motivado a la falta de linearidad en la distribución de los datos en el intervalo cercano al 50% de la curva de frecuencia acumulada. Adicionalmente, se observa que a partir de 265 mm de longitud total, más del 75% de la población se encontraba apta para el proceso de reproducción. La estimación de la talla de primera madurez por medio de la regresión de la curva linearizada, minimiza el sesgo por errores en el muestreo, así como las deficiencias en la representatividad de los ejemplares en algunas de las clases de tallas. Desde el punto de vista de la administración del recurso, esta talla debe servir de base para la selectividad de los artes de pesca con que se explota, facilitando así la liberación de los ejemplares menores a dicha talla. Colassaco (1987) estudió la misma especie en la plataforma Píritu-Unare y encontró resultados diferentes en la talla de primera madurez (261 y 268 mm para hembras y machos, respectivamente). Por otro lado, Vazzoler (1962) y Vazzoler y Braga (1983), analizando la misma especie al sur de Brasil, encontraron valores relativamente bajos para la talla de primera madurez, al estimarlas en 180 y 154 mm de longitud total para machos y hembras, respectivamente. Según estos autores, la explotación de esta especie en el sur de Brasil se efectúa en áreas muy cercanas a la costa, donde la población juvenil es muy abundante y vulnerable a los artes de pesca empleados. La proporción de juveniles en las capturas observada por estos autores superaba el 44%. Estas diferencias estarían justificadas por el comportamiento reproductivo de la especie en el área, ya que se ha determinado que el proceso de maduración está relacionado con las condiciones ambientales, geográficas y con caracteres intrínsecos de la especie, lo que induce a suponer que se tratan de stocks o poblaciones diferentes. Moffet et al. (1979) estimaron para Cynoscion arenarius en Galveston, Texas, la talla de primera madurez alrededor de 185 y 200 mm de longitud standard para hembras y machos, respectivamente. Por su parte, Shlossman y Chittenden (1981) encontraron que C. arenarius en Free Port (Texas), inicia su proceso de maduración entre 140 y 180 mm de longitud total, cuando alcanzan su primer año de edad, sugiriendo que la amplitud del intervalo de tallas depende de la estación del año. Para Cynoscion maracaiboensis la talla de primera madurez fue estimada por Espinoza (1972) en 530 y 630 mm, para machos y hembras, respectivamente, aunque esta especie alcanza tallas considerablemente mayores que C. jamaicensis. El hecho de que la población estudiada se encuentre conformada por una alta proporción (83%) de individuos en fases inmaduras o en maduración (I-III) y la baja proporción (17%) en fases madura y grávida (Marcano y Alió, 2001), pudiera estar asociado a un proceso migratorio reproductivo entre la zona de pesca y la costa. Este se desencadenaría al iniciarse la reproducción cuando los ejemplares alcanzan la talla mínima de madurez. Dicho comportamiento también fue observado por Overstreet (1983) en las tallas de primera madurez de Cynoscion nebulosus, en las costas de Mississippi, en contraposición a lo encontrado por Klima y Tabb (1959) en la misma especie, quienes no detectaron este comportamiento migratorio hacia la costa en el noreste de Florida. Los resultados encontrados en el presente estudio indican que la población de C. jamaicensis, del norte de la península de Paria inicia su proceso de maduración a tallas superiores a la talla promedio observada en las capturas comerciales (241 mm). El hecho de alcanzar su madurez a grandes tallas se debe a que en su proceso de desarrollo, parte de su ciclo vital (maduración y desove) lo cumple en zonas estuarinas y protegidas, cercanas a la costa, donde las unidades de pesca de arrastre no tienen acceso, permitiendo de este modo que los ejemplares se desarrollen normalmente y puedan alcanzar tallas promedios grandes, con la cual entran a la fase explotada. CONCLUSIONES
AGRADECIMIENTOS Deseamos agradecer la participación de los técnicos M. Márquez, G. Vizcaíno, A. Urbaneja, R. Chacón y R. Márcano, en el procesamiento del material biológico, en la trascripción y preparación de las bases de datos. Así mismo, agradecemos el apoyo de los empresarios J. Rodríguez, G. Spinalli, T. Natoli y L. Garayar al permitir las labores de investigación a bordo de sus embarcaciones de arrastre. Finalmente, el trabajo a bordo de los técnicos observadores quienes recolectaron las muestras es altamente apreciado. BIBLIOGRAFÍA
Copyright 2002 - Zootecnia Tropical. Free full-text also available online at http://www.ceniap.gov.ve/bdigital/ztzoo/ztindice.htm The following images related to this document are available:Photo images[zt02007f6.jpg] [zt02007f1.jpg] [zt02007f2.jpg] [zt02007f5.jpg] [zt02007f3.jpg] [zt02007f4.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}