 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 20, No. 4, 2002, pp. 461-472 Replacement rates of carbon stable isotope (13C) in muscle tissue of pintado, Pseudoplatystoma corruscans (Agassiz, 1829) Taxas da recolocação do isotope estável do carbono (13C) no tecido do músculo do pintado, Pseudoplatystoma corruscans (Agassiz, 1829) Valéria R. B. Furuya*, Carmino Hayashi 1, Wilson M. Furuya and Eduardo S. Sakaguti2 1
Universidade Estadual de Maringá, Departamento de Biologia,
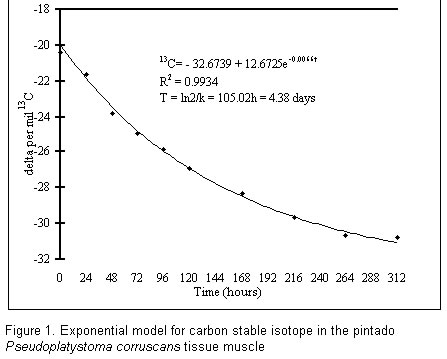
87020-900, Maringá – PR, Brasil. *E-mail: furuya@wnet.com.br Code Number: zt02029 RESUMO Este estudo foi realizado com o objetivo de avaliar a taxa de substituição do isótopo estável de carbono (13C) no tecido muscular do pintado, Pseudoplatystoma corruscans, na fase juvenil, baseado na hipótese de que o consumidor reflete isotopicamente sua dieta. Foram utilizados 480 juvenis de pintados, com peso vivo médio de 0.71 ± 0.13 g e 45 dias de vida após eclosão, distribuídos em 24 aquários de 14 l. Antes de iniciar o período experiemental e desde o início da alimentação exógena, os peixes foram alimentados com náuplios de artemia. Após este período foram alimentados com pós-larvas de lebistes (Poecilia reticulata) por 13 dias. Foram coletados dois peixes de cada aquário para análise de δ 13C. Pelo modelo exponencial, obteve-se uma meia-vida para o tecido muscular do pintado de 4.38 dias. O tempo de substituição de 99% do carbono da dieta foi de 29.91 dias. Os resultados demonstraram que o consumidor reflete a composição isotópicas de sua dieta, validando o uso do isótopo estável de carbono como marcador de carbono. Palavras-chave: 13C, isótopo estável, pintado, Pseudoplatystoma corruscans, substituição, tecido muscular. SUMMARY This work was undertaken out to the evaluate the replacement rates of carbon stable isotope (13C) in the muscle tissue of Pseudoplatystoma corruscans, during the juvenile stage is provided, based on the hypothesis that the consumer reveals its diet isotopically. Four hundred and eighty pintados, average life weight 0.71 ± 0.13 g and 45 days old, were distributed in 24 – 14 l aquariums. Previous to the experimental period and since the start of exogenous feeding, the fish were fed on nauplii artemia. After they were fed on post-larval guppies (Poecilia reticulata) for 13 days. Two fish from each aquarium were collected for d 13C analysis. Exponential model for muscle tissue of the pintado estimated a half-life equal 4.38 days. Exchanging time for 99% of carbon diet amounted to 29.91 days. Since results show that consumer reveals isotopic composition by its diet, the use of carbon stable isotope is validated in so far as time of substitution is concerned. Key words: 13C, muscle tissue, pintado, Pseudoplatystoma corruscans, replacement, stable isotope INTRODUCTION The pintado, Pseudoplatystoma corruscans (Agassiz, 1829), of the Pimelodidae family, found in the Amazon, de la Plata and São Francisco basins (Petrere, 1995), has carnivorous feeding habits and is one of the main fish species in sports and professional fishing (Godinho et al., 1989). It is the second largest predator in the Paraná River (Sato et al., 1988) with high ratings in the fishing yield (Marques, 1993). The pintado in one the freshwater species whose farming in rapidly growing in Brazil. Pintado culture had expanded from traditional pond to intensive pen culture in this country. During the 70s the use of stable isotopes became more widespread in the evaluation of energy flux in animal tissues (Tieszen et al., 1983) and since the 90s they have been employed to determine substitution rates (Gearing, 1991; Meyer-Burgdorff and Rosenow, 1995; Fraser et al., 1998). The use of C as a marker is based on the hypothesis that the consumer reveals its diet isotopically, with 13C enrichment, by increasing from 1‰ (DeNiro and Epstein, 1978; Rau et al., 1991) to 2‰ with regard to diet (Fry and Sherr, 1984; Kennedy and Krouse, 1990; France and Peter, 1997). Thus, d 13C values are not directly related to the species or to feeding habits, but to the diet predominantly consumed (Gearing, 1991; Forsberg et al., 1993). Since isotopes differ in the number of neutrons and thus in their molecular mass, they vary in their physical and chemical properties according to difference in their mass. Isotopic effect causes fractionation, or rather, changes in the isotopic abundance in a system’s various components. The enrichment of a given isotope produces a depletion of another. In the case of interpreting substitution rates, the rate in which isotopes combine in biological compartments is required. Now this varies according to species, tissue and physiological stage of the animal (Kennedy and Krouse, 1989) and to the nutrition characteristics of the food consumed (Gaebler et al., 1966). Replacement time of 13C in tissues may be high in large animals, ranging from seventy four days in adult cattle (Jones et al., 1981) to 6 – 12 days in human (Nakamura et al., 1982). Half-life, that means, the time in which final atoms of a diet incorporated are equal to half the number of total atoms, may reach ten days in humans (Katzemberg and Krouse, 1989), close to five days in shrimps (Parker et al., 1989) and four days in commercial egg-laying hens (Carrijo, 2000). Variations in d‰ 13C values may occur in the different tissues of the same animal through a carbon pool, with rapid or slow substitutions. Half-life will vary according to a given organism or tissue (Thompson and Ballou, 1956). Metabolically more active tissues, such as the liver, have a more rapid substitution than the slower ones (conjunctive tissues). While isotopic inscriptions of tissues with rapid metabolism show recent diets, slow substitutions indicate diet rates of consumed food in a previous period (Libby et al., 1964). Knowledge on the natural abundance of food sources is needed so that carbon stable isotope could be used as a marker of carbon flow to evaluate the turnover rate. Further, substitution rates should be determined so that the time necessary for total substitution of body carbon may be estimated. Our research aims at determining the substitution rate of carbon in the muscles of juvenile specimens of the pintado (Pseudoplatystoma corruscans) in lab conditions, by the isotopic variation of d‰ 13C. MATERIALS AND METHODS Experiment was undertaken at the Aquaculture Lab of the Biology Department of the State University of Maringá from 16 to 28 January 2000. Four hundred and eighty fishes juvenile specimens of the pintado (P. corruscans) were used. With an average live weight 0.77 ± 0.15 g and forty four days old after hatching, they were obtained by induced reproduction at the Research and Training Center in Aquaculture (CEPTA) in Pirassununga, SP, Brazil. Fish were distributed in 24 aquariums (14 l each), with sides and bottom covered with black tarpaulin, and maintained by an aeration system through a porous stone linked to a central blower. Organic material was siphoned from the aquarium daily at 8 h. Before to the experimental period (since the start of exogenous feeding), the fishes were fed on nauplii of artemia, every six hours, from 5 to 23 h, during forty days. After forty two days they were fed on post-larvae guppy (Poecilia reticulata), mean live weight 0.01 g, from the Botanical Garden of Maringá, during thirteen 3 days. Both types of food were fed to satiation. A sample of 600 fishes was selected for d‰ 13C lab analysis and dehydrated in a forced aeration buffer for 48 h. A pool with two pintados from each aquarium was formed for d‰ 13C analysis. Head, viscera, fins and skin of fish were removed and a composed sample of guppies was employed for the analysis of the isotope. Fish collection was undertaken daily until the seventh day and henceforth every two days. Samples were dehydrated in forced aeration buffer during 48 h at 50°C and ground during three minutes in a cryogenic mill at -190°C. They were then conditioned in labeled glass bottles for later analysis at the Stable Isotopes Lab of the Bio-Science Institute of UNESP, Botucatu, SP, Brazil. Physical and chemical characteristics of water were monitored every three days. Temperature, pH, electrical conductivity (μS/cm) and dissolved oxygen were measured by 0 – 50°C thermometer (Quimis-401B), conductivity meter (Quimis-405P), portable digital pH meter (Quimis-400H) and oxygen meter (Quimis-408P) respectively. Calculation of ratio of isotope 13C/12C for the analysis of natural variation in δ‰ by mass spectrometer required carbon conversion of matrix or sample in CO2. Carbon was obtained by destruction process in which all the carbon in the sample was oxidized till its transformation into carbon dioxide, at high temperature in an oxygen atmosphere, for 5 minutes, as described by Ducatti et al. (1979). Combustion underwent three stages. The first stage comprised the purification of oxygen flow (<0.5 kgf cm-2) used in combustion. Oxygen was burnt in a quartz oven with granulated copper oxide (CuO) at 450°C. The CO2 obtained was contained in an ascarite chemical trap. In the second stage sample underwent combustion in a Vyscor oven with granulated CuO at 900°C, under oxygen pressure higher than 1 atmosphere. In the third stage gases formed during combustion were isolated and retained (Carrijo, 2000). Halogens remained in the quartz oven with metallic silver at 45°C, whereas sulphur oxide and nitrogen oxide were kept in the chemical trap with manganese dioxide. Water and CO2 formed during combustion of the sample remained in the cryogenic trap made up of dry ice, alcohol (-80°C) and liquid nitrogen (-190 °C) respectively. Stages were executed under a continuous flow of oxygen which was aspired by dual-stage mechanical pump. Routine procedures followed Licatti (1997). Samples and standards were analyzed in a dual-access mass spectrometer of isotopic ratios, DELTA - Finnigan Mat, with six collectors gauged to Software Isodat (Finnigan Mat, 1994) which provided correction and 13C/12C ratio according to international PDB standard with equation suggested by Carrijo (2000): d ‰ 13C = Ra/Rp 1000, where d ‰ 13C = relative enrichment of sample measured by PDB standard and Ra and Rp = isotopic ratio 13C/12C of sample and of standard respectively. Values are in d ‰ measured by PDB standard; each sample was done in duplicate, with analysis error of <0.4‰ and standard deviation less than 1‰ within sample. Data were analyzed by Origin® 6.0 Professional’s regression equation method (Microcal Software, 1999). For quantitative measurement of the velocity of carbon substitution of diets in fish tissue after a certain time interval, an exponential time equation was used: theoretical differential equation Y = A + B e-kt, where Y is the value of d ‰ 13C of tissue under analysis; A is the initial condition; B is the asymptotic rate of d ‰ 13C for the tissue; k is the ‘substitution’ rate of carbon in the tissue; t is time (in hours) as from diet substitution. Experimental results of relative enrichment, d ‰ of ratio 13C/12C with regard to time (in days) were consequently obtained. Half life (T) of 13C for muscle tissue for 50% of each diet in t = T was calculated by equation T = ln 2/k, where T is half life; ln is Napierian logarithm and constant k the time unit, which provides the ‘velocity’ in the exchange process of stable isotopes in the tissues. It is a constant of the substitution rate of the tissue’s isotope. RESULTS AND DISCUSSION Values for temperature (26.69 ± 0.84°C), pH (6.92 ± 0.24), electrical conductivity (12.89 ± 0.78 m S/cm) and dissolved oxygen (5.92 ± 0.24 mg/l) did not vary greatly throughout the experiment and remained within normal levels for tropical fish. Values of d ‰13C when fish were fed on nauplii of artemia show that the forty-day period comprising the start of exogenous feeding until the beginning of feeding on guppies (Poecilia reticulata) was sufficient for body carbon exchange. In fact, homogeneity in values obtained during the last collections of fish fed on nauplii of artemia has been reported. A higher rate of carbon exchange was recorded until the 9th day of collection (216 h) which occurred in -1‰ day on an average. Gradual decrease in incorporation rate was reported after this period; increment was slight after eleven days (Table 1).
Mean values of d ‰ 13C of carbon stable isotope in the muscle tissue of the pintado fed on nauplii of artemia during the pre-experiment period varied from -20.40 to -30.79‰. Value of muscle tissue of the pintado fed on post-larvae guppies at the end of the experiment amounted to 0.36‰ heavier when compared to its diet. Result confirms the supposition that consumer reflects its diet in an isotopic manner. A 13C enrichment occurred in approximately 1‰ (DeNiro and Epstein, 1978; Rau et al., 1991) to 2‰ with regard to diet (Fry and Sherr, 1984; Kennedy and Krouse, 1990; France and Peter, 1997). Substitution of 99% of diet carbon with a confidence interval of 95% reached 29.91 days (717.84 h) (Figure 1). Time for this substitution was higher than that found by Parker et al. (1989) for shrimp (Penaeus vannamei), taking into consideration d 13C as a natural marker in the food of the crustaceans with an equilibrium of d 13C of muscle tissue after twenty one days. A half-life of 4.38 days was obtained for the pintado’s muscle in the suggested exponential model. Time was lower than that spent in tissue substitution in humans (Katzemberg and Krousie, 1989), which amounted to ten days, so that the incorporation of the number of final atoms of the diet could be equal to half the total atoms. This is close to results by Parker et al. (1989), which were close to five days for muscle tissue of the shrimp (Penaeus vannamei) and slightly lower, or four days, for the liver of laying hens (Carrijo, 2000). Carbon substitution rate varies for each species and for each tissue (Kennedy and Krouse, 1989), as may be surmised in the research by Tieszen et al. (1983) and Carrijo (2000) on the jerboa (Meriones unguiculatus) and the laying hen respectively. According to Tieszen et al. (1983), tissues with quick metabolism indicate recent diets, while those with low metabolic substitution indicate long-period diets. Metabolically active tissues have a more rapid carbon substitution rate (Tieszen et al. 1983), whereas those with a short half-life develop high incorporation speed in stable isotopes. Libby et al. (1964) state that, as a rule, metabolically more active tissues, such as liver, pancreas and fat tissues, have more rapid ‘substitution’ rates than less active ones, such as bones and conjunctive tissue. This fact has been shown by Carrijo (2000) when different tissues of the laying hen were analyzed. Biochemical fractionation may also be influenced by assimilation rate of macronutrients in the consumed food (Gaebler et al., 1966; Kennedy and Krouse, 1989). In our research a diet with near characteristics to that which fish of this species feed on in their habitat has been preferred so that a compatible rate of isotope incorporation could be achieved. The interpretation of the substitution rate requires knowledge of rate at which consumed isotopes are incorporated and eliminated in a specific biological compartment. Jones et al. (1981) obtained equilibrium in the d 13C value of adult cattle hair after seventy four days. This period was higher than that found by Nakamura et al. (1982) for human hair, six to twelve days, and also higher than values of approximately twenty five days for the egg and of twenty to thirty five days for the liver of laying hens (Carrijo, 2000). According to Kennedy and Krouse (1989), reading of ‘substitution’ rate, or rather, the rate isotopes are incorporated and eliminated in a specific biological compartment, should be evaluated in animals in a state of equilibrium. This occurs in adult animals, since it is difficult to distinguish the incorporation of new from growing tissue in young ones. Due to difficulties in obtaining and maintaining adult fish in lab conditions, since many factors may influence consumption and utilization of food, only fish in the growing stage have been used in our research. One of the limiting factors in our study was the information on mechanisms by which stable isotopes undergo fractionation. Since there is a lack of information on the exchange rate of stable carbon isotope in the muscle tissue of the pintado, these are important for studies on isotope compositions of diet and tissues. Results show the feasibility of the use of carbon stable isotope as a ‘substitution’ rate marker of carbon in the muscle tissue of the pintado. CONCLUSIONS Exponential model for muscle tissue of the pintado estimated a half-life equal 4.38 days. Exchanging time for 99% of carbon diet amounted to 29.91 days. The stable isotopic composition of the carbon in an pintado tissue muscle reflects the 13C/12C ratio of its diet. BIBLIOGRAFIA
Copyright 2002 - Zootecnia Tropical. Free full-text also available online at http://www.ceniap.gov.ve/bdigital/ztzoo/ztindice.htm The following images related to this document are available:Photo images[zt02029f1.jpg] | ||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}