 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 21, No. 3, 2003, pp. 237-259 Growth of Donax denticulatus (Linné, 1758) (Bivalvia Donacidae) in La Guardia inlet, Margarita Island, Venezuela Crecimiento de Donax denticulatus (Linné 1758) (Bivalvía: Donacidae) en la ensenada La Guardia, isla de Margarita, Venezuela Jesús S. Marcano1*, Antulio Prieto2, Asdrúbal Lárez1 y Hebel Salazar1 Instituto Nacional de Investigaciones Agrícolas, CIAE Sucre-
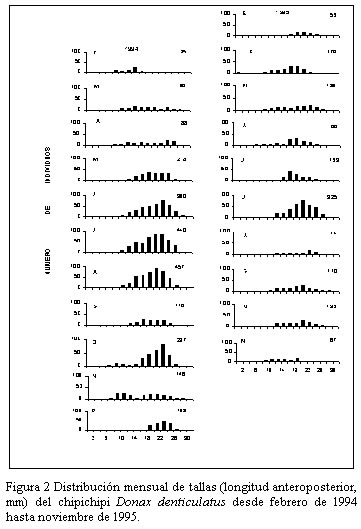
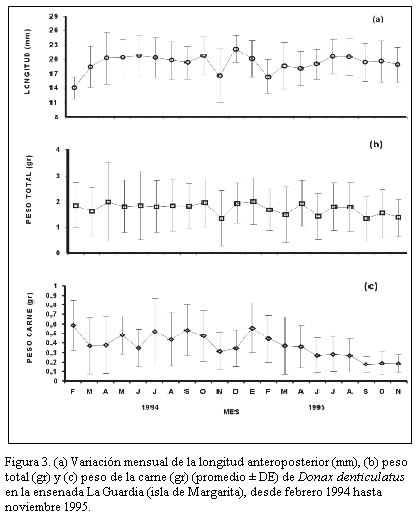
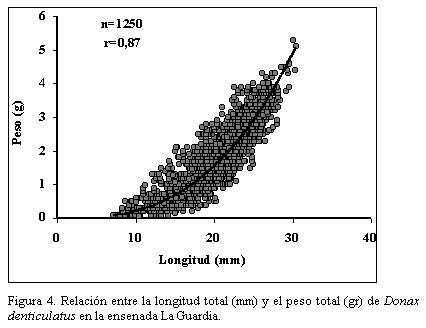
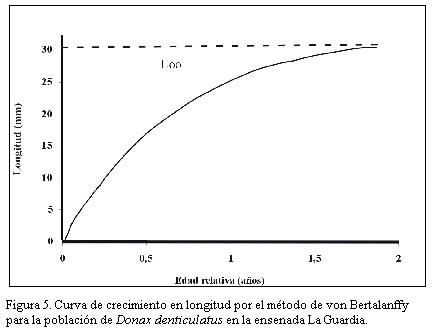
Nueva Esparta. Apartado 236. Cumaná 6101, estado Sucre. Venezuela. *Correo-E: jmarcano@inia.gov.ve Recibido: 03/07/03 Aceptado: 14/01/04 Code Number: zt03016 SUMMARY Donax denliculatus is a dominant member of high energy sandy beach communities of the Venezuelan coasts in the Caribbean Sea, a population of this bivalve was sampled monthly in La Guardia inlet Margarita island, between February 994 and November 1995. Quantitative samples were taken in perpendicular transects at l1 stations to determined length-frequency distribution and growth. The population structure presented a size range between 2 and 32 mm length, being mainly formed by individuals from 14,1 to 22,2 mm, that were dominant from march to October. Recruitrnent rate was low, and the major contribution of recruits was found in October – November 1994 and February – March 1995. The low body weight during November and March seem to be related to the reproductive cycle of this specie. Growth parameters of the von Bertalanffy seasonal growth equation were L¥ =30.02 mm, k= 1.79 year-1, C= 0.60 and ts= 0.2, with a faster growth rate between February – August and a life span of 1.5 year. Key words: Seasonal growth, population structure, Donax denticulalus, Margarita island, the Guard cove. RESUMEN Donax denticulatus es un miembro dominante en las comunidades de playas arenosas marinas de alta energía de las costas venezolanas en el mar Caribe. En el presente trabajo se muestreó la especie, desde febrero 1994 hasta noviembre 1995 en la ensenada de La Guardia isla de Margarita. Muestras cuantitativas fueron tomadas sobre transectas perpendiculares a la línea de la costa, en 11 estaciones para determinar la distribución de tallas y el crecimiento. La estructura de tallas de la población presentó un intervalo entre 2 y 30 mm, estando constituido principalmente por bivalvos con tallas entre 14,1 y 22,2 mm, que fueron dominantes entre marzo y octubre. La tasa de reclutamiento fue baja y la mayor contribución de los reclutas fue encontrada en octubre - noviembre 1994 y febrero- marzo 1995. El bajo peso corporal de los individuos durante noviembre y marzo, parece estar relacionado con el ciclo reproductivo de la especie. Los parámetros de la ecuación de crecimiento estacional de von Bertalanffy fueron L∞ = 30,02 mm, K= 1,79 mm año-1, C = 0,60 y t = 0,20, indicando un rápido crecimiento entre febrero - agosto y una longevidad de 1,5 años. Palabras clave: Crecimiento estacional, estructura poblacional, Donax denticulalus, isla de Margarita, ensenada La Guardia. INTRODUCCIÓN El bivalvo Donax denticulatus, Linné 1958, llamado comúnmente chipichipi, pertenece a la familia Donacidae y se distribuye desde el mar Caribe, a lo largo de las costas suramericanas hasta Brasil. En Venezuela es abundante en las playas arenosas de fuerte oleaje, desde el estado Zulia hasta nororiente de Venezuela (Novoa et al., 1998), siendo uno de los cuatro bivalvos mas consumidos en la isla de Margarita y en el estado Sucre por la alta densidad que presenta en la playas (Etchevers, 1975). El chipichipi puede alcanzar una longitud anteroposterior de 30 mm, con una talla media de alrededor de 25 mm. Puede recolectarse sobre la arena, hasta una profundidad de 20 cm, porque con el pie y la forma aguzada de su concha, se entierra rápidamente. En algunas playas arenosas se encuentran sumergidos siempre donde la columna de agua es inferior a 1 metro (Gómez, 1999), manteniéndose en movimiento según la marea y la actividad del oleaje, estrategia de movimiento y migración continua que es energéticamente costosa (Levinton, 1982). La utilización de materia orgánica particulada como alimento, hace que estos bivalvos tengan mayor disponibilidad cerca de la desembocadura de los ríos; situación que le ocasiona a muchos organismos marinos dificultades fisiológicas, pues en estos lugares, las fluctuaciones de salinidad en el agua pueden ser mayores. Sin embargo, D. denticulatus parece no tener problemas ya que en estado silvestre tolera salinidades sobre 35‰ y en condiciones experimentales salinidades tan bajas como 1,25‰ (Castagna y Chanley, 1973) Las investigaciones sobre crecimiento en poblaciones de moluscos en las zonas templadas son abundantes (Lavoie et at., 1968; Burke y Mann, 1974; Warwick y Price, 1975; Guillou y Moal, 1980; Godoy, 1982; Brethes et al., l986), pero escasas en las tropicales y subtropicales (Laudien et al., 2001; Nel et al., 2001). Para el mar Caribe existen investigaciones sobre D. denticulatus, en aspectos ecológicos (Wade, 1967), historia de vida (Wade, 1968), morfología funcional (Wade, 1969), el efecto de los factores ambientales y los patrones de dispersión espacial (Sastre, 1984, 1985). En Venezuela existen investigaciones sobre D. denticulatus en aspectos bioecológicos (Gómez, 1956; Etchevers, 1975; Vélez, 1975; Farache, 1980; Brito, 1981; Vélez y Venables, l98l a, b; Vélez et al., 1985 y Rodríguez, 1987); aspectos ecofisiológicos (Neuberg, 1982; Martín et al., 1983 y Carboniní et al., 1997) y aspectos bioquímicos (López, 1985 y De Mahieu et al., 1987, 1988). Las estadísticas oficiales de este molusco reflejan niveles de capturas relativamente bajos de esta especie y actualmente no existe normativa legal que regule su extracción. Sin embargo, se ha recomendado que la pesca se permita de noviembre a abril (Gómez, 1999). De allí la necesidad de estudiar mas el crecimiento de esta especie para poder elaborar un plan de manejo a futuro, acorde con una explotación sostenible del recurso. En el presente trabajo se analiza la estructura de tallas de D. denticulatus en la ensenada La Guardia, isla de Margarita, y se estiman sus parámetros de crecimiento. MATERIALES Y MÉTODOS Área de estudio La ensenada La Guardia está situada en la zona norte de la isla de Margarita, localizada geográficamente entre los 11° 00´00´´ N y 64° 00´ 16´´ O, en su extremo occidental (El Saco) y los 11° 3´ 15´´ N y 64° 12´21´´ O en el extremo oriental (La Guardia). Está formada por una barra arenosa de 23,5 km de longitud que une a la población de La Guardia con la península de Macanao (Figura 1). Las características ecológicas del ambiente fueron descritas por Etchevers (1975) y Mendoza y Marcano (2000). Muestreo de la población Las muestras de D. denticulatus fueron recolectadas desde febrero de 1994 hasta noviembre de 1995 en 11 estaciones, con separación de 2 km entre ellas y en cada transecta-estación por mes. Para la obtención de las muestras de D denticulatus, se empleó un cilindro de acero inoxidable de 15,5 cm de diámetro y 25 cm de altura, el cual se introdujo en el sustrato de la zona de distribución del bivalvo, entre 0 y 6 m perpendicular a la costa, y se extrajeron tres cilindros (réplicas) al azar. Seguidamente el sedimento se pasó a un tamiz de 2 mm de apertura para separar los bivalvos. A los ejemplares de D. denticulatus recolectados en las muestras de cada estación se les determinó la longitud anteroposterior (Lt, mm), utilizando un vernier con líneas de división 0,05 mm, peso total (Pt, g), peso de la carne húmeda (PCH, g) y el peso seco libre de concha (PSLC, g) en una balanza con resolución 1 mg. La composición de tallas de la fracción muestreada de la población de chipichipi se determinó por mes mediante el uso de histogramas de frecuencias de talla (mm), de elaborados con intervalos de clase de 2 mm de longitud. El crecimiento relativo del chipichipi se determinó mediante el establecimiento de la relación existente entre el peso total del animal (P) en gramos y la longitud anteroposterior (L) en mm, la relación entre ambas variables se expresa mediante la ecuación: P = a Lb donde: a = intercepto y Los parámetros de crecimiento L∞ (longitud máxima teórica) y K (coeficiente de crecimiento) se estimaron utilizando la forma estacional de crecimiento de von Bertalanffy. Con este propósito se utilizó, en primer lugar el método de Bhattacharya (1967) para descomponer gráficamente las distribuciones plurimodales de frecuencias, suponiendo normalidad de cada componente. Con la finalidad de obtener los estimados preliminares de L∞, K y los valores de oscilación estacional se utilizo la técnica de Gulland y Holt (1959). Posteriormente, tomando los valores obtenidos por este método, se usó el análisis de superficie de respuesta del ELEFAN 1 (Gayanilo et al., 1989) para determinar los valores en los parámetros de crecimiento que presentaron el valor más elevado del criterio de bondad de ajuste (Rn) a fin de obtener la forma estacional del modelo de crecimiento estacional. De acuerdo con Pauly y Gatschutz (1979), este modelo puede ser representado por la expresión: Lt = L∞ [1-exp(-k(t-to) + (CK/2p ) sen 2p (t-ts))] donde: C, representa la amplitud de oscilación del crecimiento En cada muestreo se determinó la temperatura superficial, salinidad del agua y la materia orgánica del sedimento, por los métodos descritos por Strickland y Parson (1972). RESULTADOS La población de D. denticulatus, analizada en base a las distribuciones de frecuencia mensual de 3627 individuos recolectados durante el período comprendido entre febrero 1994 y noviembre 1995 en la ensenada La Guardia, presentó un amplio intervalo de tallas comprendidas entre 2 y 30 mm. (Figura 2). El mayor porcentaje de individuos correspondió a las tallas entre 14 y 22 mm durante todo el período de estudio. Las distribuciones mensuales de longitud fueron bimodales y la proporción de bivalvos con tallas inferiores a 7 mm de longitud fue escasa, presentándose la mayor incidencia en octubre y noviembre (Figura 2). La distribución de la cantidad total de individuos recolectados a lo largo de la franja de 6 m de playa estudiados presentó una distribución normal, con la máxima concentración entre los 3 y 4 m. La variación mensual promedio de la longitud señala valores mínimos desde octubre de 1994 a febrero de 1995 (Figura. 3a), y el promedio mensual del peso total durante el período de estudio osciló entre 1,3 y 2,0 g (Figura 3b) y entre 0,3 y 0,6 g para el peso de la carne (Figura 3c). En noviembre se observaron los menores promedios de peso y longitud y la mayor variación del peso total. La relación entre la longitud del cuerpo (Lt) y el peso total (PT) de D. denticulatus durante el período de estudio se expresó por la ecuación PT = 0,0004 Lt 2,7311 (r = 0,87, n = l.250) (Figura 4). Para la relación entre el peso seco libre de concha (PSLC) y, la longitud del cuerpo, se obtuvo la ecuación PSLC = 0.0019 Lt 2,81 ( r = 0,81; n = 602). Los parámetros de crecimiento de D denticulatus para
el período febrero 1994 noviembre 1995, determinados a partir de distribuciones
de talla agrupadas en clases de 2 mm de longitud anteroposterior, permitieron
obtener con el método gráfico de Gulland y Holt (1959) estimados preeliminados
de longitud asintótica de L∞ = 29,42 K = 1,58 año, C = 0,50 y ts = 0,15.
Posteriormente, utilizando la técnica de Guayanilo et al., (1989) se obtuvieron mejores ajustes con valores de L∞ = 30,02 mm, K = 1.79, año to = 0, C = 0,60 y ts = 0,20 (Figura 5). De esta manera, la expresión final del crecimiento estacional puede ser representada por la ecuación: L∞ = 30,02 [1- e –1,79t + (0,60. 1 ,79/2p ) sen 2p (t -0,20)] la cual indica que el período de mayor crecimiento en la población ( ts = 0,20) para ambos años corresponde al lapso comprendido entre febrero y agosto. Según este modelo, la tasa de incremento en longitud es alta (3,6 mm/mes) durante los primeros 3 meses después del reclutamiento, disminuyendo luego (2,3 mm/mes) durante los próximos 4 meses y muy baja (1 mm/mes) después de los 9 meses de edad. En la zona de estudio la temperatura mínima del agua intersticial en el sustrato del hábitat de D. denticulatus varió de diciembre a mayo entre 22 y 25 °C y fueron máximas de julio a noviembre, cuando se registraron valores entre 27 y 29 ˚C. Igualmente la materia orgánica contenida en el sedimento osciló entre 1 y 2,9 % con un promedio de 1,57 %, y el material en suspensión varió entre 33 y 77,8 mg /1. DISCUSIÓN La distribución de clases de tallas en Donax denticulatus observada en la presente investigación coinciden con la señalada por Novoa et al. (1998), quienes reportaron que en la región oriental, el mayor volumen de la captura de esta especie se encuentra conformado por individuos con tallas comprendidas entre 15 y 20 mm de longitud y que la máxima longitud alcanzada es de 24 mm. Igualmente las distribuciones mensuales bimodales, así como la escasa proporción de bivalvos con tallas inferiores a 7 mm, los cuales presentaron la mayor incidencia en octubre y noviembre, sugieren la existencia de un reclutamiento máximo durante estos meses, coincidiendo con lo informado por Vélez et al. (1985) para una población de D. denticulatus en Punta Araya, estado Sucre. Wade (1969) indica para la misma especie en Jamaica, que el mayor pico de reclutamiento ocurre en el período abril-mayo. En la presente investigación se observaron bajos reclutamientos en el lapso abril-mayo de 1994 y 1995, debido probablemente a una alta mortalidad de post larvas o a formas con diferentes tasas de crecimiento dentro de una cohorte, como ha sido señalado para otras especies tropicales (Moore y López, 1970; Vélez, 1976). Laudien et al. (200l) han señalado que Donax serra, en playas de Namibia, presentó discontinuidades en el ciclo anual de reproducción, fenómeno que relacionaron con la temperatura superficial del agua. La máxima concentración de bivalvos observada entre los 3 y 4 m coincide con los señalamientos previos de Etchevers (1975) en la misma área y Wade (1967) en Puerto Rico. Probablemente las condiciones del área de estudio: la pendiente, el oleaje y el sustrato así como la ubicación de D. denticulatus en relación con Donax striatus y T. mactroides, especies con las que coexiste en el área, influyen en la variación de a distribución por tallas de D. denticulatus durante el período de estudio (Etchevers, 1975) Los factores que determinan la distribución de estas especies de bivalvos en la zona intermareal arenosa han sido discutidos por Wade (1967) y Ansell et al. (1980) quienes señalan como los principales responsables a la granulometría, el grado de exposición a las olas y el contenido orgánico de la arena; por esta razón las poblaciones de D. denliculatus se presentan en playas aluviales cerca de las desembocaduras de los ríos o pantanos. Penchaszadeh y Olivier (1975) citan que D. hanleyanus tiene una distribución vertical en las costas bonaerenses que varía en relación a la morfología de las playas, la intensidad del oleaje, la marca y las características sedimentológicas. Los valores obtenidos de materia orgánica y de material en suspensión en el área de estudio son más elevados que los reportados en otras áreas del Caribe (Marcano, 1993); en Jamaica el sedimento contiene solamente 0,15% de materia orgánica (Wade, 1969). Estos altos valores en la ensenada La Guardia podrían explicar las altas densidades de la especie reportadas por Etchevers (1975). Los bajos valores promedios de la longitud observados desde octubre 1994 a febrero 1995 pueden estar relacionados con el ciclo reproductivo de la especie; cabe destacar que Vélez et al. (1985) definen dos períodos anuales en el ciclo reproductivo de D. denticulatus (enero- junio y julio - diciembre), presentándose mayor actividad reproductiva entre julio y diciembre. Este fenómeno podría explicar la ubicación en noviembre de los mas bajos valores de longitud. Si se estima que los organismos presentan una longevidad máxima de aproximadamente 18 meses, esto indicaría que una nueva cohorte estaría ocupando el hábitat, resultados estos que concuerdan con el patrón reproductivo señalado para la especie en otras áreas del Caribe (Wade, 1968; Trueman, 1971; Vélez et al., 1985). El período con temperaturas máximas y mayores variaciones de temperaturas diurnas se correspondieron con el de mayor actividad reproductora y de reclutamiento lo que indica que estas fluctuaciones de temperatura son importantes en la época de reproducción de bivalvo tropicales (Penzias, 1969; Vélez et al., 1985). Los incrementos de crecimiento obtenidos en esta investigación para Donax denticulatus son semejantes a las reportadas para la misma especie en Jamaica (Wade, 1969), Punta Araya, Venezuela, (Vélez, 1985) y en Playa Brava, Venezuela (García et al., 2003) y para D. cuneatus en la India (Nayar, 1954). Sin embargo, son muy superiores a las reportadas para D. variabilis y D. timidus en Texas, EUA (Loesch, 1957), pero inferiores a la D. dentifer en Costa Rica (Palacios et al., 1983) (Cuadro 1). En el coeficiente de crecimiento obtenido en el presente estudio para D. denticulatus de la ensenada La Guardia fue superior al reportado por Vélez et al. (1985) en playas de la península de Araya (K = 0,43) debido a que este último fue calculado con base a meses. Igualmente la longitud asintótica (L∞ = 30,02 mm) también fue superior a la obtenida por este último autor para la misma población (L∞ = 18.5 mm) a pesar de estas diferencias, ambas estimaciones indican que al año se alcanzaría mas del 90% de la longitud asintótica (Cuadro 1).
Investigaciones realizadas en D. trunculus (Guillou y Moal, 1980; Ansell et al., 1980; Gaspar et al., 1995) en playas atlánticas europeas y en la franja mediterránea, reportan valores de K inferiores a este trabajo. No obstante, los L∞ reportados por estos autores son superiores a los obtenidos para D. denticulatus. Estos resultados se deben probablemente a que los bivalvos de las zonas templadas presentan menor tasa metabólica en comparación con sus homólogos tropicales (Pereira et al., 1988). De allí que los bivalvos tropicales crezcan más rápidamente que los de las zonas templadas, pero estos últimos experimentan mayor longevidad (Lavoie et al., 1968; Caddy y BiIlard, 1976; Brethes et al., 1986). Sin embargo dentro de una misma especie los bivalvos longevos poseen tallas mayores que los jóvenes. Se ha observado que los organismos marinos tropicales, en su mayoría de vida corta presenta un crecimiento mas acelerado de sus contra partes de altas latitudes y alcanzan la L∞ en corto tiempo con una tasa de crecimiento elevada (Urban y Tesch,1996). La longevidad estimada de 18 meses para esta población es semejante a la informada para otras poblaciones de la misma especie en el Caribe tropical, (Brito, 1981; Vélez et al., 1985; García et al., 2003) pero resultó ser muy inferior a la de D. spiculum del océano Indico tropical que vive sólo 6 meses (Ansell et al., 1978). De acuerdo a los resultados obtenidos, puede pensarse que la población analizada podría ser sensible a una explotación pesquera para bivalvos con tamaño comercial (>19 mm Lt). Sin embargo es necesario realizar estudios sobre biomasa, mortalidad, manejo y ordenación de a pesquería CONCLUSIONES
AGRADECIMIENTOS Queremos expresar nuestro agradecimiento a los técnicos de la Estación Local de Nueva Esparta del INIA: Hargadys Sanabria, Dimas Rodríguez, Francisco Gómez y Andrés Carrión, por su cooperación en la recolección y procesamiento de las muestras. Especial reconocimiento al Lic. José Alió por la corrección del manuscrito y sus sugerencias. BIBLIOGRAFÍA
The following images related to this document are available:Photo images[zt03016f4.jpg] [zt03016f5.jpg] [zt03016f2.jpg] [zt03016f1.jpg] [zt03016f3.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}