 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Zootecnia Tropical, Vol. 22, No. 1, 2004, pp. 49-69 Fijación de nitrógeno por Leucaena leucocephala, Gliricidia sepium y Albizia lebbeck y su transferencia a las gramíneas asociadas Nitrogen fixation of Leucaena leucocephala, Gliricidia sepium and Albizia lebbeck and its transfer to the associated grasses Selina Camacaro[1]*, Juan C. Garrido1 y Wilfre Machado[2] [1]Instituto de Producción Animal, Facultad de
Agronomía, Universidad Central de Venezuela. Apartado
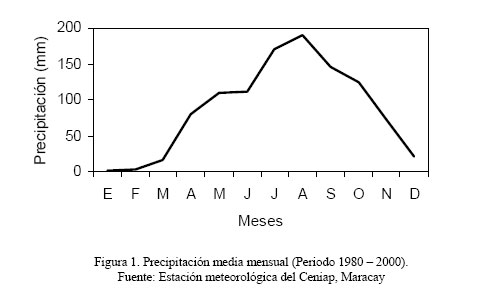
4579, Maracay 2101, estado Aragua, Venezuela. *Correo-E: camacaros@agr.ucv.ve Code Number: zt04004 RESUMEN Se realizó un estudio en el campo experimental del Instituto de Producción Animal, Facultad de Agronomía, de la Universidad Central de Venezuela, con el objeto de evaluar la fijación de nitrógeno (N) de Leucaena leucocephala, Gliricidia sepium y Albizia lebbeck y su transferencia a una pastura mixta asociada (Cynodon dactylon x Digitaria swazilandensis). Se utilizaron parcelas asociadas de 10x2 m, a dos densidades de siembra (4.000 y 8.000 plantas/ha) y parcelas de gramíneas de 4x5 m, fertilizadas con seis niveles de nitrógeno (0, 50, 100, 200, 400 y 800 kg N/ha). Se utilizó un diseño completamente aleatorizado con tres repeticiones y 11 tratamientos. Se evaluaron las variables materia seca de gramíneas, materia seca de leguminosas herbáceas, materia seca de leguminosas arbustivas, materia seca comestible, materia seca de malezas, rendimiento total de nitrógeno y nitrógeno fijado y transferido. Los resultados obtenidos en producción de biomasa, rendimiento de N, fijación y transferencia de N, evidencian que con estas tres leguminosas es posible sustituir la fertilización nitrogenada en un rango de 0 a 200 kg N/ha. Por otro lado, el factor densidad de siembra tiene efecto solo para L. leucocephala, en cuanto a fijación y transferencia de N se refiere. Además, la habilidad de A. lebbeck de ceder una mayor proporción del N fijado sugieren su potencial para ser asociado con gramíneas con mayor demanda de N, en las condiciones en las que se desarrolló el experimento. Palabras clave: Leucaena leucocephala, Gliricidia sepium, Albizia lebbeck, fijación de N, transferencia de N. SUMMARY A study in the experimental field of the Animal Production Institute, Agronomy Department, of the Universidad Central de Venezuela was carried out in order to evaluate the N fixation of Leucaena leucocephala, Gliricidia sepium, and Albizia lebbeck and its transfer to an associate mixed pasture (Cynodon dactylon x Digitaria swazilandensis). Associated plots of 10x2 m were used with two densities (4,000 and 8,000 plants/ha) and grass plots of 4x5 m, fertilized with six nitrogen levels (0, 50, 100, 200, 400 and 800 kg N/ha). A randomized design with three repetitions and 11 treatments was used. The following variables were evaluated: grass dry matter, herbaceous legumes dry matter, dry matter of shrub legumes, edible dry matter, weeds dry matter, nitrogen total yield and fixed and transferred nitrogen. The results obtained in biomass production, yield of N, fixation and transfer of N evidenced that with three shrub legumes is possible to substitute the nitrogen fertilization in a range from 0 to 200 kg N/ha. On the other hand, the sowing density factor had an effect on L. leucocephala only, for fixation and transfer. Also, the ability of A. lebbeck of giving a bigger proportion of the fixed N suggests its potential to be associated with grasses with more demand of N, under the conditions that the experiment was developed. Key words: Leucaena leucocephala, Gliricidia sepium, Albizia lebbeck, Nitrogen fixation, Nitrogen transfer. INTRODUCCIÓN El rápido aumento de la población en la mayoría de los países tropicales, impone severas restricciones a los recursos naturales, por lo tanto hay un deterioro progresivo de los suelos (Greenwood y McKenzie, 2001; Pinzón y Amézquita, 1991), como consecuencia de la deforestación, agricultura intensiva y prácticas de manejo ganadero muy perjudiciales tales como: el sobrepastoreo y el fuego; además los suelos tropicales tienen una muy baja fertilidad natural (Sanchez y Salinas, 1981), lo cual limita la producción y calidad de las pasturas, y por consiguiente no cubre los requerimientos de los animales. En tales condiciones, las leguminosas arbustivas forrajeras adquieren una importancia relevante debido a que utilizan principalmente, para sus procesos fisiológicos, el nitrógeno atmosférico fijado simbióticamente por algunos géneros de bacterias de la familia Rhizobiaceae (Whiteman, 1980). Parte de ese nitrógeno puede ser transferido a la gramínea acompañante, aunque la proporción de esa transferencia puede ser influenciada por muchos factores: especie, condiciones edafoclimáticas, densidad de siembra, etc. (Blair et al., 1990). En algunos experimentos con asociaciones de gramíneas y leguminosas arbustivas se han encontrado aumentos en la cantidad de biomasa y mejoras en la calidad de la gramínea (Escobar et al., 1995), elevando significativamente la productividad por unidad de superficie (Cachtpoole y Blair, 1990; Belsky, 1992); por esta razón es de suponer que la leguminosa está siendo determinante en la asociación y que la fijación y transferencia de nitrógeno a la gramínea acompañante es considerable. Se sabe que la fertilización no sólo aumenta considerablemente los costos de producción, sino que también puede causar problemas de contaminación ambiental (Bohlool et al., 1992). En este sentido, la asociación de leguminosas arbustivas y gramíneas en los potreros, resultaría en una disminución del uso de fertilizantes químicos, mantenimiento del equilibrio ecológico, disminución de la contaminación y aporte de fuentes alimenticias a las explotaciones ganaderas. El propósito de este trabajo fue evaluar el efecto de la incorporación de tres leguminosas arbustivas (Leucaena leucocephala, Gliricidia sepium y Albizia lebeck) sobre la producción de biomasa total, rendimiento de N total y la fijación de nitrógeno y su transferencia a las gramíneas acompañantes (Digitaria swazilandensis y Cynodon dactylon). MATERIALES Y MÉTODOS El estudio se realizó en el campo experimental del Instituto de Producción Animal, Facultad de Agronomía, Universidad Central de Venezuela, Maracay, Venezuela (67º 35´ O y 10º 14´ N), a 450 msnm. El área está ubicada en una zona de vida bosque seco tropical, con promedios anuales de precipitación 942 mm, con una época lluviosa desde mayo a noviembre y una seca desde diciembre a abril, con el 96,8 y 3,2%, respectivamente, de las precipitaciones totales (Figura 1), humedad relativa de 82% y 25,5ºC de temperatura (INIA, Estación Meteorológica Maracay, 2000). El suelo utilizado fue clasificado como un Fluventic Haplustoll (Márquez, 1989), perteneciente a la serie Maracay (Ostos, 1993) franco arenoso mixto, bien drenado y de fertilidad media (Cuadro 1).
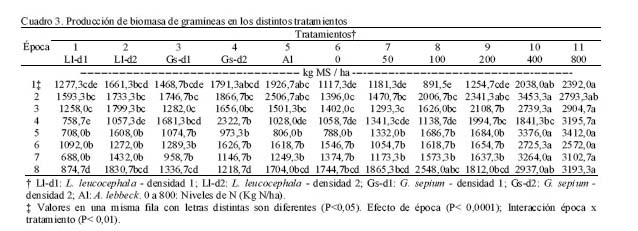
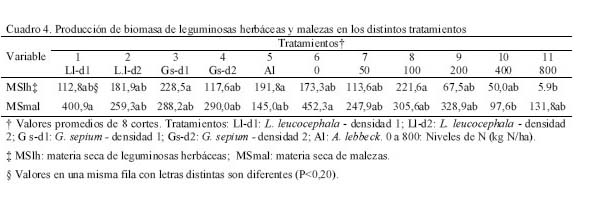
Para este estudio se utilizó la técnica descrita por Whiteman (1980), para lo cual se establecieron parcelas asociadas (10 x 12 m) (gramíneas + leguminosa) a dos densidades de siembra (d1: 2,5 x 1 m, 4.000 plantas/ha y d2: 2,5 x 0,5 m, 8.000 plantas/ha) de las leguminosas Leucaena leucocephala (T1-d1 y T2-d2), Gliricidia sepium (T3-d1 y T4-d2)y Albizia lebbeck (T5, solo d1). Además, se trabajó con parcelas de gramíneas solas (Digitaria swazilandensis y Cynodon dactylon) (4 x 5 m), fertilizadas con seis niveles de nitrógeno (urea como fuente) (T6: 0, T7: 50, T8:100, T9: 200, T10: 400 y T11: 800 kg N/ha, respectivamente). Las parcelas asociadas se establecieron en Diciembre de 1998, sobre una pastura ya fundada de Digitaria swazilandensis y Cynodon dactylon. El período de evaluación fue de 14 meses (Julio 1999 hasta Septiembre 2000). A los seis meses de establecida la leguminosa, se efectuó una poda a 50 cm y 5 cm de altura para leguminosa y gramínea, respectivamente. Los muestreos de gramíneas y leguminosas fueron realizados cada seis semanas, a partir de la fecha de poda de uniformización (Junio 99) y retiro del el material cortado de las parcelas. El área experimental fue regada semanalmente durante la sequía y no fue pastoreada. Cada nivel de nitrógeno se aplicó fraccionado una semana después de cada muestreo. Para las leguminosas sólo se cosechó las tres hileras centrales considerando el resto como bordura, tomando una muestra compuesta por tratamiento y por repetición. Las muestras se secaron a estufa a 60ºC hasta peso constante, para determinar materia seca y N (AOAC, 1980). Para la gramínea de las parcelas asociadas y solas se procedió de manera similar que para las leguminosas. La fijación aparente de N (kg/ha), en las parcelas asociadas se estimó totalizando la producción de N de la leguminosa + N de la gramínea asociada, de ese total se restó la producción de N de la gramínea sola (T6). La transferencia aparente de N (kg/ha) se estimó restándole al N de la gramínea asociada la producción de N de la gramínea sola (T6); ambas metodologías son descritas por Johansen y Kerridge (1979). Se utilizó un diseño completamente aleatorizado con tres repeticiones, realizándose un análisis de varianza y una prueba de medias de Duncan para las variables materia seca de gramíneas (MSgr), materia seca de leguminosas arbustivas (hoja + tallo) (MSla), materia seca comestible (MScom: MSgr + MSlh + MSla), nitrógeno fijado y transferido. A las variables materia seca de leguminosas herbáceas (MSlh) y materia seca de malezas (MSmal) no se les detecto la homogeneidad de la varianza, por lo que se utilizó la prueba de Friedman (α= 0.20). RESULTADOS Y DISCUSIÓN Producción de biomasa de Leucaena leucocephala, Gliricidia sepium y Albizia lebbeck La MSla fue muy similar durante los primeros tres muestreos (Cuadro 2), evidenciándose el potencial de las tres leguminosas arbustivas, puesto que no hubo diferencias entre ellas, con excepción del T3 (G. sepium-d1), que produjo menos biomasa que L. leucocephala (d1 y d2) (P<0.05) para los tres muestreos.
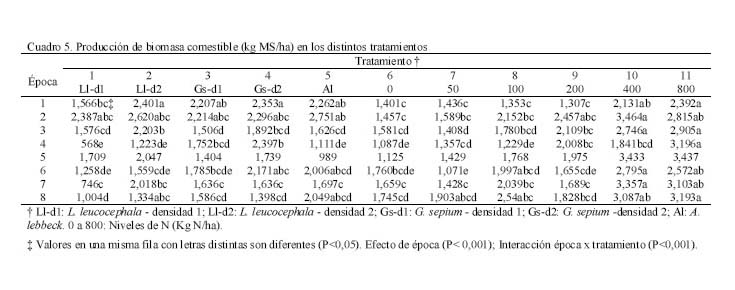
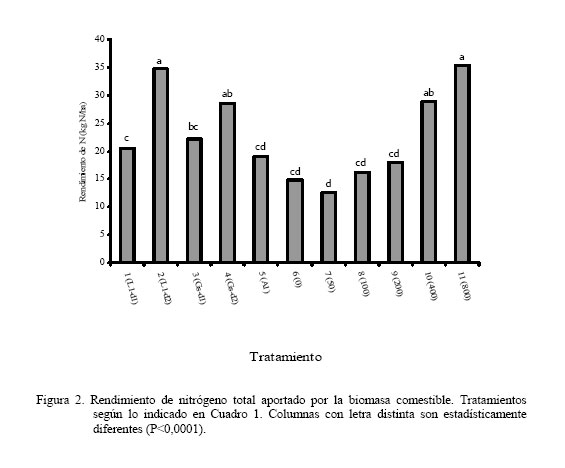
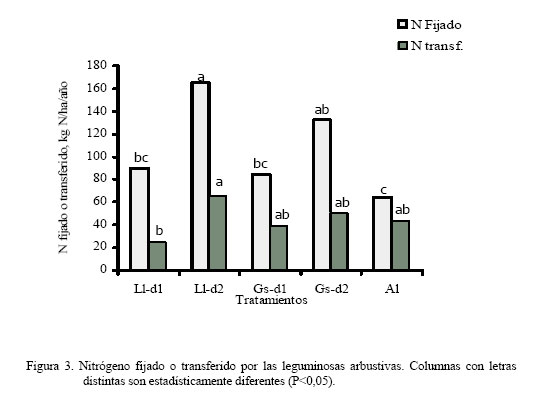
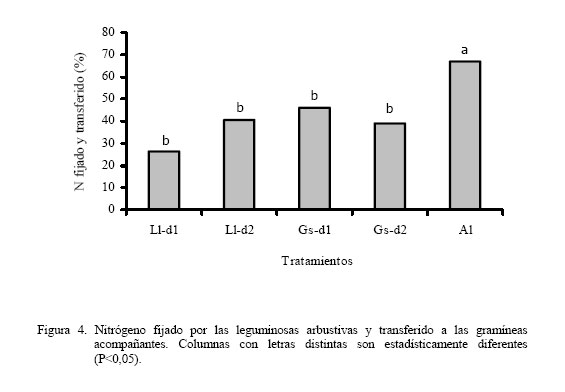
Los dos primeros muestreos coincidieron con la época lluviosa (Figura 1) y el tercero con un período de transición lluvia-sequía; sin embargo, el cuarto coincide con la época seca, lo cual pudiera haber afectado el crecimiento de las leguminosas arbustivas, trayendo como consecuencia que no hubiera diferencias en la producción de biomasa. Por otro lado, a pesar que para el quinto muestreo todavía era época de sequía, la A. lebbeck mostró superioridad con respecto al resto de los tratamientos (P<0,05), excepto con el T4. La respuesta de la A. lebbeck, en cuanto a producción de biomasa, pudiera estar relacionada con su adaptación a zonas semiáridas y muy secas (Lowry et al., 1994), como se observa claramente en el sexto muestreo. Sin embargo, ya para el séptimo muestreo la A. lebbeck sólo fue superior a los tratamientos 1 y 3 (P<0,05), resultando similar a los tratamientos con d2 (2 y 4), debido quizás al inicio de las lluvias, que favoreció el crecimiento de todas las especies. Para el octavo muestreo, A. lebbeck y L. leucocephala, no mostraron diferencias entre ellas, pero si entre la primera y la G. sepium-d1 (P<0,05). La MSla obtenida muestra un efecto marcado de época (P<0,001) e interacción época x tratamiento (P<0,001) y una tendencia de las leguminosas L. leucocephala y G. sepium a producir mayor cantidad de biomasa cuando son sembradas a la densidad 2. Por otro lado, A. lebbeck se muestra muy promisoria para las condiciones de bosque seco, ya que su comportamiento en líneas generales fue similar a las otras dos, a pesar de haber sido evaluada sólo a una densidad. En general, la MSla obtenida, para las tres especies, fue muy baja comparada con lo reportado en la literatura para las mencionadas leguminosas (Faría-Marmol, 1996; Dávila y Urbano, 1996; Escobar et al., 1995; Palma, 1997; Francisco et al., 1996). Estas diferencias pudieran explicarse por la influencia de muchos factores, tales como disponibilidad de agua, fertilidad del suelo, densidad de siembra, frecuencia de corte, etc. En este caso particular, la frecuencia de corte (42 días), la disponibilidad de agua en el suelo y la corta edad de las plantas al inicio del experimento, podrían haber sido determinantes para tales resultados. Al respecto, Wong y Mohd (1986), trabajando con L. Leucocephala y G. sepium en condiciones de mayor precipitación (2.260 mm) encontraron valores similares a los reportados en este trabajo, para 4 y 8 semanas de corte, en plantas muy jóvenes (1 a 3 años). Producción de biomasa de gramíneas Durante los ocho muestreos (Cuadro 3) los tratamientos 10 y 11, mostraron su superioridad (P<0,05) en concordancia con lo reportado en la literatura en relación a la alta respuesta de Digitaria swazilandensis y Cynodon dactylon a dosis crecientes de N (Flores y Parra, 1976; Hernández y Cárdenas, 1983). Por otro lado, los tratamientos asociados resultaron similares a los tratamientos 10 y 11, para el primero y segundo muestreo, respectivamente (Cuadro 3), con la excepción del tratamiento 1 que en el primer muestreo que resultó diferente (P<0,05), lo que muestra los efectos positivos de la asociación de leguminosas arbustivas (Bronstein, 1984; Catchpoole y Blair, 1990, Belsky, 1992; Escobar et al., 1995; Elgersma y Hassink, 1997). Aunque, en algunas investigaciones (Dunn et al., 1994; Jayasundara et al., 1997) de asociaciones de gramíneas de mediana a baja calidad con leguminosas no reportan efectos significativos. En este caso se piensa en un efecto resultante de la poca habilidad en este tipo de gramíneas para utilizar nutrientes del suelo (Humpreys, 1981). Para el caso de los muestreos que coincidieron con la época de transición lluvia-sequía (muestreo 3) y sequía (muestreos 4 y 5), los tratamientos 10 y 11 difirieron (P<0,05) del resto de los tratamientos, con excepción de los tratamientos 3, 4, 7 y 9 en el muestreo 4, los cuales fueron similares (P>0,05) al tratamiento 10. La respuesta de los tratamientos en estos muestreos está asociado, evidentemente, a un efecto de época (P<0,001) y de interacción época x tratamiento (P<0,001). Sin embargo, cuando se inicia el período de lluvia (muestreos 6 y 7), el comportamiento de los tratamientos fue similar al mostrado en el muestreo 5; esto pudo ser debido a que la precipitación de los meses abril a julio del año 2000 fue sensiblemente más baja que en años anteriores (INIA, 2000). Para el último muestreo, la producción de biomasa volvió a ser muy heterogénea como se observa en el muestreo 4, a pesar de corresponderse con el período de lluvia. Los tratamientos 8, 10 y 11 fueron similares entre ellos, con la excepción del tratamiento 11 el cual resultó superior (P<0,05) al resto de los tratamientos. En el presente experimento las densidades utilizadas y las especies de leguminosas no produjeron un efecto significativo en los resultados de MSgr. Al respecto, la literatura reporta que debe haber un balance en las densidades para que haya una respuesta positiva (Hall, 1978; Whitney y Green, 1969) además de la respuesta ínterespecífica que se genera (Giller et al., 1991). Los resultados obtenidos en este ensayo indican que es posible reemplazar la fertilización nitrogenada de gramíneas (0 a 200 kg N/ha) con las leguminosas asociadas. Producción de biomasa de leguminosas herbáceas Por los resultados obtenidos (Cuadro 4) pareciera que la presencia de leguminosas arbustivas y la fertilización a dosis bajas de N, favorece la existencia de leguminosas herbáceas, caso contrario a lo ocurrido con la aplicación elevada de N, ya que hubo diferencias (P<0,20) entre el tratamiento 11 y los tratamientos 3, 5 y 8, pero no (P>0,20) entre estos y el resto. Sin embargo, Dunn et al. (1994) no encontraron efectos significativos sobre las leguminosas herbáceas ni por efecto de especie ni de densidad, cuando evaluaron A. lebbeck, Casuarina cunninghamiana y Eucalyptus maculata. Las leguminosas herbáceas constituyen un componente importante de la biomasa comestible, puesto que son consumidas por los animales y también fijan y transfieren nitrógeno, como lo demostraron Elgersma y Hassink (1997), quienes evaluaron Trifolium repens asociado con Lolium perenne, encontrando una fijación aparente de N desde 150 a 545 kg N/ha y la transferencia desde 21 a 37% del N fijado. Producción de biomasa comestible En los dos primeros muestreos (lluvia) (Cuadro 5), los tratamientos asociados (1 al 5) fueron similares a los tratamientos fertilizados con altas dosis de N (10 y 11), con la excepción del tratamiento 1, que difirió del 2, 4 y 11 (P<0,05), en el primer muestreo. En los dos siguientes muestreos, 3 y 4, que se correspondieron con transición lluvia-sequía y sequía, respectivamente, el comportamiento fue menos uniforme que en los primeros dos muestreos, encontrándose diferencias (P<0,05) entre los tratamientos 10 y 11 y el resto de los tratamientos, con la excepción del tratamiento 10, el cual que resultó similar a los tratamientos 3, 4 y 9. La respuesta de los tratamientos en el muestreo 4 pudiera estar relacionada con que durante este muestreo no hubo diferencias en la producción de biomasa de leguminosas arbustivas (Cuadro 2). La MScom de los tratamientos para el muestreo 5 (sequía) muestra claramente el efecto de época (P<0,001), no observándose diferencias significativas en ninguno de los tratamientos. Durante el muestreo 6, de los tratamientos asociados, sólo G. sepium-d2y A. lebbeck fueron similares a los tratamientos 10 y 11, pudiendo ser esto un indicador de la rápida recuperación de estas leguminosas después de un período de sequía. Una vez que se estableció de nuevo el período de lluvias (muestreos 7 y 8), la respuesta de los tratamientos fue más uniforme que durante la sequía, observándose un efecto de la interacción época x tratamiento (P<0,001), similar a lo observado en los primeros dos muestreos. Por otro lado, en el muestreo 7, de los tratamientos asociados el 2 resultó similar al 11, y en el muestreo 8, sólo los tratamientos 2 y 5 resultaron similares al 10 y 11. La respuesta obtenida en la MScom a lo largo de todo el experimento pudiera estar condicionada por factores, tales como frecuencia de corte, edad de las plantas de leguminosas arbustivas, remoción total de biomasa después de cada corte, ya que era de esperarse una producción de biomasa mayor, puesto que ésta variable esta conformada por biomasa de gramíneas, leguminosas arbustivas y herbáceas. Catchpoole y Blair (1990) demuestran esto al conseguir diferencias significativas en la producción de biomasa comestible entre leguminosas arbustivas en monocultivo, cultivos asociados gramíneas-leguminosas y gramíneas solas. A pesar de los bajos valores de MScom, los tratamientos asociados (T1 al T5) mostraron valores similares, en la mayoría de los casos, a los tratamientos fertilizados con altas dosis de N (10 y 11). Estos resultados señalan el beneficio económico de la inclusión de leguminosas en las pasturas al reducir los costos por fertilización mineral. La participación porcentual de las leguminosas en la MScom varió de 18% en T3 hasta 26% en T1, en las parcelas asociadas. En las parcelas de gramíneas solas, varió de 0,2% en T11 hasta 12% en T6 y T8, lo cual representa ventajas adicionales, ya que las leguminosas aportan minerales que normalmente son deficitarios en las gramíneas (N, Ca, P, S, etc.), y la presencia de taninos en las leguminosas protege la proteína de la degradación ruminal, proporcionando N adicional para su absorción en el intestino delgado (Norton, 1994). Producción de biomasa de malezas La presencia de malezas en los diferentes tratamientos fue muy variable (Cuadro 4). Los tratamientos 1 y 6, fueron superiores (P<0,20) al tratamiento 10, pero no al resto de ellos (P>0,20). Por otro lado, los tratamientos 1 y 6 mostraron los menores valores de gramíneas (Cuadro 3) y el T10 mostró el valor más bajo de malezas. Lo anterior pareciera mostrar una relación de competencia entre las gramíneas deseables y las malezas, ya que estas últimas también eran gramíneas, no deseables, que competían por los recursos luz, agua y nutrientes, con las deseables. Por otro lado, hubo la tendencia a que las parcelas asociadas mostraran, en promedio, una biomasa de malezas mayor (277 kg MS/ha) que las parcelas de gramíneas fertilizadas (261 kg MS/ha). Esto pudiera estar asociado con la mayor habilidad de Digitaria swazilandensis y Cynodon dactylon de responder a dosis crecientes de N (Flores y Parra, 1976; Hernández y Cárdenas, 1983). Rendimiento de nitrógeno total La asociación de gramíneas con las leguminosas arbustivas L. leucocephala y G. sepium(T2 y T4) produjeron cantidades similares de N a los tratamientos con altas dosis de N (T10 y T11) (Figura 2). Hubo mayor producción de N con la L. leucocephalaa d2 que con d1 (P<0,001). G. sepiumse comportó similarmente en esta variable entre ambas densidades. Por otro lado A. lebbeck fue similar a las d1 de ambas leguminosas. Los beneficios aportados por la asociación pueden apreciarse mejor, si se expresan en base anual el T7, el cual fue el más bajo (P<0,001), rindió 100 kg N/ha/año, y el T11 aportó 282,4 kg N/ha/año, seguido por el T2 con 278,9 kg N/ha/año. Además, puede suponerse que los aportes de N por parte de las leguminosas serían mayores, bajo otras condiciones, puesto que en el actual experimento, después de cada muestreo hubo una remoción total del material remanente, reduciendo la disponibilidad de residuos orgánicos para procesos tan importantes como la mineralización. Los rendimientos de N reportados en este experimento son mayores que los reportados por Jayasundara et al. (1997) para las leguminosas L. leucocephala, G. sepium, y Senna siamea (no fijadora de N2), pero no a los reportados para L. leucocephala, G. sepium, Calliandra calothyrsus y Sesbania grandiflora por Catchpoole y Blair (1990). Es importante considerar la especie de gramínea presente en la asociación, ya que se conoce la habilidad diferencial de las gramíneas para la utilización del N disponible en el suelo y por otro lado ha sido sugerido que la competencia ínter especifica puede modificar los patrones de consumo de N en una leguminosa (Giller et al., 1991). La asociación de gramíneas-leguminosas representa una ventaja adicional, ya que al incluir una leguminosa en los potreros no sólo se obtienen beneficios de mayor producción de biomasa y N, como consecuencia de la fijación y transferencia, si no un aumento en el aporte de otros nutrientes críticos en la alimentación animal, como el P, Ca, S, etc. (Norton, 1994). Nitrógeno fijado y transferido Los resultados obtenidos (Figura 3) indican claramente la habilidad de Leucaena leucocephala y Gliricidia sepium de fijar altas tasas de N lo cual ha sido demostrado ampliamente en diferentes trabajos (Danso et al., 1992; Awonaike et al., 1992; Liyanage et al., 1994; Jayasundara et al., 1997), sobre todo cuando se siembran a altas densidades. Por otro lado, los reportes de literatura sobre los procesos de fijación y transferencia de N en A. lebbeck son muy escasos, por lo que los resultados obtenidos en este trabajo son alentadores además de novedosos. Los valores obtenidos en este trabajo para L. leucocephala y G. sepium fueron más bajos que los reportados por la literatura, lo cual pudiera estar condicionado por las prácticas de manejo aplicadas al ensayo, tales como la frecuencia de corte de 42 días, disponibilidad de agua en el suelo y la corta edad de las plantas de leguminosas arbustivas al inicio del experimento. Las diferencias en la cantidad de N fijado, como resultado de la edad de las plantas de leguminosas arbustivas también ha sido reportado por Muofhe y Dakora (1999), quienes obtuvieron con Aspalathus linearis valores mas bajos para plantas de 1 y 2 años con respecto a aquellas de 3 años. Por otro lado, Albizia lebbeck fue capaz de fijar tanto nitrógeno como las otras dos leguminosas, a la densidad de 4.000 plantas/ha (Figura 3). Sin embargo, A. lebbeck fue capaz de transferir la mayor proporción de N (P<0,05) (Figura 4). La importancia del factor densidad para la fijación y transferencia de N parece ser determinante en L. leucocephala, pero no en G. sepium y A. lebbeck. Catchpoole y Blair (1990) y Jayasundara et al. (1997) no reportaron diferencias para la transferencia de N de L. leucocephala y G. sepium a la gramínea acompañante (7,6 y 21% de N transferido, respectivamente). Por otro lado, los reportes de leguminosas herbáceas parecieran indicar una mayor transferencia de N que las arbustivas. Se han obtenido valores desde 21 a 37% N fijado transferido para Trifolium repens (Elgersma y Hassink 1997) y desde 12 al 17% para Macroptilium atropurpureum, Neonotonia wightii, Desmodium intortum y Lotononis bainesii (Johansen y Kerridge, 1979). CONCLUSIONES Bajo las condiciones en las cuales se realizó el experimento se concluye:
Los resultados obtenidos en producción de biomasa, rendimiento de N, fijación y transferencia de N, evidencian que con estas tres leguminosas es posible sustituir la fertilización nitrogenada en un rango de 0 a 200 kg N/ha. El factor densidad de siembra en cuanto a la fijación y transferencia de N debe ser considerado cuando la leguminosa es L. leucocephala. A. lebbeck es una especie que aporta N para las asociaciones con gramíneas de alta demanda de este elemento al menos en las condiciones en las que se desarrolló este experimento. BIBLIOGRAFÍA
Copyright 2004 - Zootecnia Tropical. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}